 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
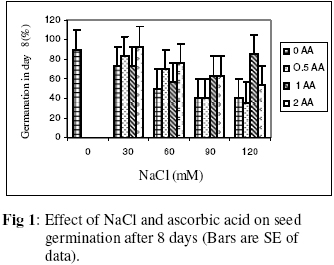
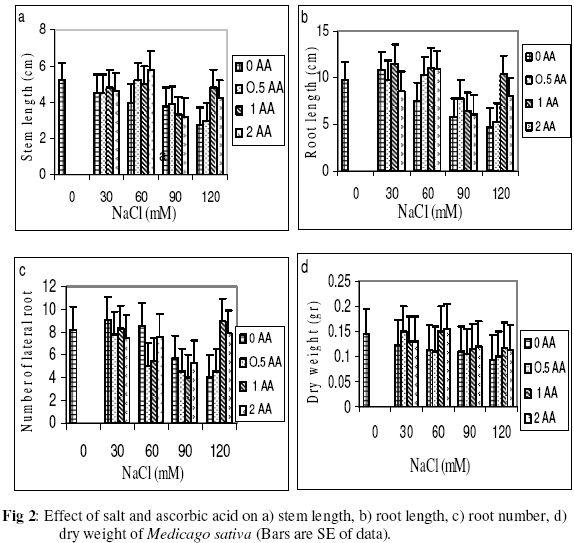
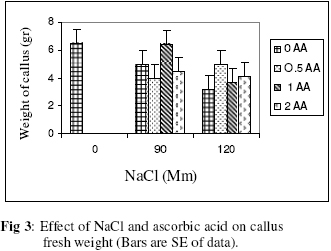
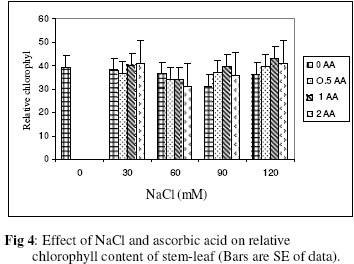
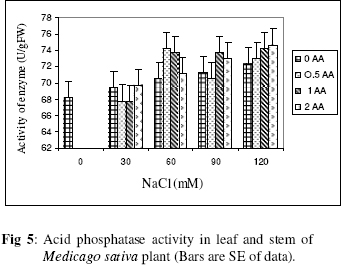
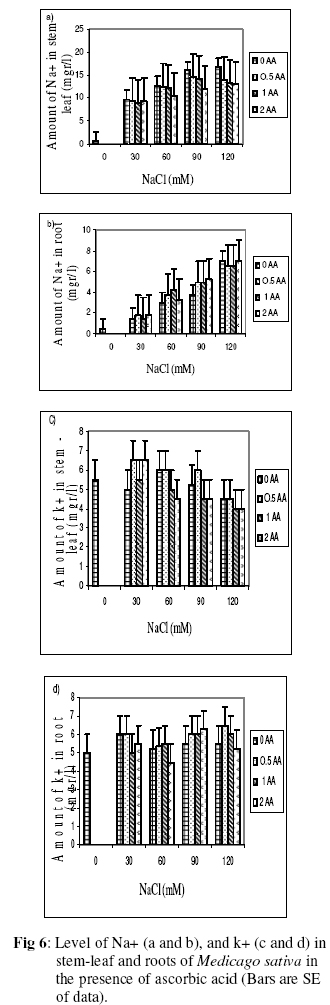
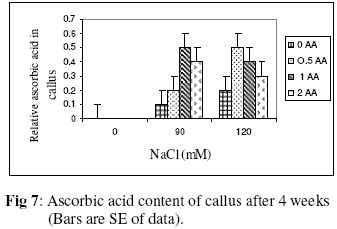
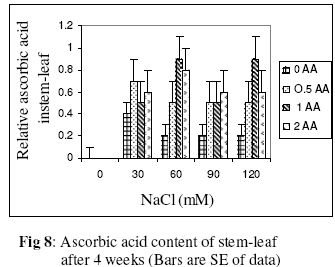
Biokemistri, Vol. 18, No. 2, December, 2006, pp. 63-69 The effects of ascorbic acid on salt induced alfalfa (Medicago sativa L.) in in vitro culture Lila ARAB and Ali Akbar EHSANPOUR* Department of Biology, Faculty of Science, Isfahan University, Isfahan, Iran *Author to whom correspondence should be addressed. E-mail: ehsanpou@yahoo.com Received 17 February 2006 Code Number: bk06010 Abstract Ascorbic acid as an antioxidant agent has already been used for increasing of stress tolerance. Callus was produced from stem segments of alfalfa (Medicago sativa L.) on MS medium supplemented with 2,4 dichlorophenoxy acetic acid, naphthalene acetic acid and kinetin (2 mg/l each).Calli were then transferred to the same medium containing 0, 30, 60, 90, 120 mM NaCl and 0, 0.5, 1.0, 2.0 mM ascorbic acid. Addition of ascorbic acid to the medium improved seed germination and also increased the activity of acid phosphates, chlorophyll content, and dry mass. The Na+ and K+ content of stem-leaf and root was relatively increased with some variations. The fresh weight of calli was also increased by ascorbic acid under salt stress condition Keywords: Ascorbic acid, in vitro, Medicago sativa, salt stress INTRODUCTION Salinity induces a wide range of metabolic perturbation in higher plant. Although plant in nature have evolved several adaptation mechanisms to cope with the presence of salt in their environment, an understanding of these mechanisms still remains incomplete, mainly due to the complexity of salt stress, which present an ionic component, and an osmotic component on the other hand1. Moreover, physiological characterization of salt affected plants in natural environments remains difficult. Salin soils are typically very patchy in their salinity2, and ions other than Na and Cl could contribute to the high electrical conductivity of salt-affected lands3. Medicago species are relatively salt tolerant4. The production of activated oxygen species (AOS) which can damage DNA, protein, chlorophyll and membrane function is a by-product of oxidative metabolism in chloroplasts, mitochondria and peroxisomes. AOS production is further enhanced in response to various abiotic stress, such as drought, salt, extreme temperature and herbicides. Numerous studies have shown that the level of antioxidative enzyme is increased when plants are exposed to biotic or abiotic stresses5,6. Various plant tissues are known to accumulate up to milimolar concentration of L-ascorbic acid. It plays an important role as an antioxidant and protects the plant during oxidative damage by scavenging free radicals and active oxygen species that are generated during salt stress condition7,8. In vitro tissue culture is a useful tool to study the cellular mechanisms of the stress and its application at the cellular level, independent of regulatory mechanisms occurring at the whole plant level. Salt resistant line has been selected using plant cell culture techniques9. Nevertheless, the phenomenon of physiological adaptation to NaCl stress is not completely understood. The present study was therefore undertaken to investigate the effects of various salt concentrations and exogenous ascorbic acid on plant growth under in vitro condition. MATERIALS AND METHODS Seeds of Medicago sativa cultivar (cv) Rehnany were obtained from the Seed and Seedling Production Center in Isfahan, Iran. Seeds were surface sterilized for 1-2 min in 95% ethanol (v\v) then treated in 20% sodium hypochlorite for 20 min, followed by 3 washes with sterile distilled water and were placed on MS medium containing 0, 30, 60, 90, 120 mM NaCl and 0.5, 1, 2 mM exogenous ascorbic acid under aseptic condition. Then for 2-10 days post germination, and after 4 weeks, growth parameters (stem and root length, number of lateral root and dry weight and relative cholorophyl by chlorophyll meter (Min-LTA, 5RAD-502), acid phosphatase according to method of Julie et al.10 and Na+, K+ level in tissues were determined using flame photometer (Perkin-Elmer Coleman 51-ca). For callus production, 50 stem segments from in vitro grown plants on MS (Murashige and Skooge, 1962)11, were cultured on MS medium supplemented with NAA (naphthalene acetic acid), 2,4-D (2,4 dichlorophenoxy acetic acid), kinetin, (each at 2 mg/l) and 1 g/l yeast extract. All cultures were kept in culture room at 25°C. Calli were produced after 4-6 weeks, then were sub-cultured at 2 weeks intervals. After 3 subcultures, 5 segments of callus (approx. size 8-10 mm) with 10 replications were transferred to the same medium containing 0, 90, 120 mM NaCl and 0.5, 1, 2 mM ascorbic acid. Fresh weights of the calli were measured after 4 weeks. Approx. 1 gram callus per each treatment with 3 replications was used for ascorbic acid determination using the method of Smirnoff12. All data were analyzed according to Duncan test at p<0.05. RESULTS The effects of salt and ascorbic acid on germination of Medicago sativa seeds are as shown in Figure 1. Result indicated that seed germination decreased with increasing NaCl concentration, but exogenous ascorbic acid in the medium increased germination even at 90 and 120 mM salt. Growth parameters such as stem and root length, number of root and dry weight were affected by both NaCl and ascorbic acid (Fig. 2 a, b, c, d). These parameters were decreased as concentration of salt was increased in the medium, however, plant growth were improved by adding exogenous ascorbic acid significantly. Addition of ascorbic acid to the culture medium contaning NaCl improved growth of callus in Medicago sativa cv. Rehnany significantly (p<0.05) as shown in Figure 3. When high concentration of NaCl were applied to the medium, callus fresh weight were decreased dramatically. The exogenous ascorbic acid increased fresh weight significantly (p<0.05) at concentration of 120mM NaCl. Similar response was observed in chlorophyll content of stem and leaf (Fig 4). Acid phosphatase activity was also measured in leaf and stem from in vitro grown plants under salt stress. Comparison between salt treated and control shows that enzyme activity increased by increasing of salt concentration. However, it seems that enzyme activity was positively affected by adding ascorbic acid but the difference between ascorbic acid treated and non treated callus was not significant (Fig. 5). The pattern of Na+ and K+ in stem-leaf and roots of plants cultured in medium containing salt and ascorbic acid is shown in Figure 6. Sodium content of stem-leaf and roots were increased due to increasing NaCl in the medium, but potassium level was basically higher than sodium. Ascorbic acid at 2.0 mM in the medium decreased the level of Na+ content of stem-leaf and roots significantly (p<0.05). The amount of ascorbic acid in callus was measured four weeks post culture using spectrophotometer at 600 nm. The results are shown in Figures 7 and 8. The level of ascorbic acid content in cells after 4 weeks was increased as the amount of this antioxidant was increased in the medium. However, the highest amount of ascorbic acid was observed when, 0.5 and 2.0mM ascorbic acid was added to the medium. Discussion Salinity is a major factor in reducing plant growth and productivity. The effect of salt stress on plant and corresponding callus depends on four responses: dehydration of the cells through the low water potential, nutritional imbalance caused by the interference of saline ions with essential nutrients in both uptake and translocation processes, toxicity due to the high accumulation of Na and Cl in the cytoplasm as well as the production of activated oxygen species during salt stress. A common response to increasing the salt concentration is decreasing of growth rate of callus. It has been reported that under salt stress condition the growth of sensitive callus of Lycopersicon esculentum is decreased dramatically. Moreover, when high concentration of exogenous proline (10mM) and NaCl were applied to the medium the dry weight of Medicago sativa cv. Yazdi was increased13. The exogenous ascorbic acid in the medium increased germination, dry weight, and ascorbic acid content in stem-leaf tissues as well as callus. In fact exogenous ascorbic acid increased level of ascorbic acid uptake by different tissues as reported by Arrigoni et al.14. One of the remarkable roles of ascorbic acid in seed germination and cell growth under salinity is its anti-oxidant activity, rather than its possible utility as an organic substrate for respiratory energy metabolism. Several finding indicate that salt-induced low growth of plant and callus is associated with increasing of the cell damaging active oxygen species (AOS). The effect of additional ascorbic acid on plant survival is associated with the partial inhibition of a few interactions in AOS production15. The inhibitory effect of ascorbic acid on lipid peroxidation is related to plant survival but its actual mechanism(s) are not yet clear. One possibility is that addition of ascorbic acid would increase the leakage of essential electrolytes following peroxidative damage to plasma membrane16,17. The amount of sodium accumulation in stem-leaf decreased when 1.0 and 2.0mM ascorbic acid was added to the medium. No significant effect of ascorbic acid addition to the medium was found on potasium level of stem-leaf. We have found that ascorbic acid increased number of roots in salt treated seedlings. In fact the new roots and leaves produced by ascorbic acid under salt stress condition might be due to the cell division and differentiation of meristem cells. When ascorbic acid was added to the medium, despite of the concentration levels, the activity of acid phosphatase was increased. It has been well documented that salt and water stress increase acid phsphatase activity. Acid phosphatase is known to act under salt stress by maintaining a certain level of inorganic phosphate which can co-transported with H+ along a gradient of proton motive force. In contrast, a few reports indicating that acid phosphatase activity is independent of phosphate level18. However, the relationship between ascorbic acid and phosphate level and acid phsphatase activity remind to be evaluated in details in the future. In conclusion, this report shows that Medicago sativa plant treated with ascorbic acid remarkably increase the capacity of germination, seedling survival by changes or balancing Na/K level as well as intracellular content of ascorbic acid. Results indicating that the salt tolerance level was increased using ascorbic acid. ACKNOWLEDGEMENTS Authors wish to thank all members of Graduate Directorship of Isfahan University for their support of this project ReferenceS
© 2006 Nigerian Society for Experimental Biology. The following images related to this document are available:Photo images[bk06010f4.jpg] [bk06010f3.jpg] [bk06010f2.jpg] [bk06010f1.jpg] [bk06010f7.jpg] [bk06010f6.jpg] [bk06010f8.jpg] [bk06010f5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}