 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biokemistri, Vol. 19, No. 2, Dec, 2007, pp. 43-48 Cofactor interactions in the activation of tissue non-specific alkaline phosphatase: Synergistic effects of Zn2+ and Mg2+ ions Femi J. Olorunniji*, Adedoyin Igunnu, Joseph O. Adebayo, Rotimi O. Arise and Sylvia O. Malomo Department
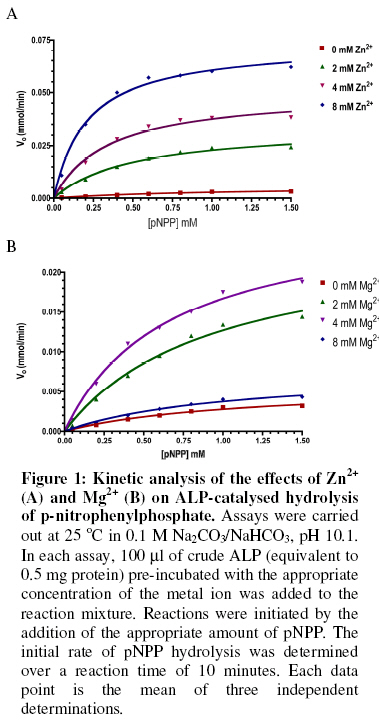
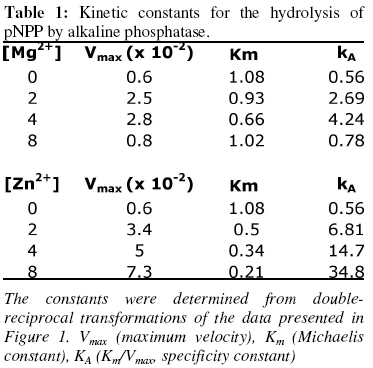
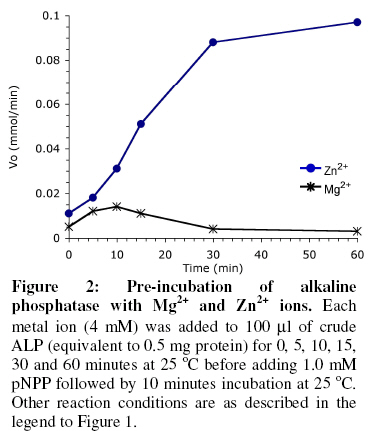
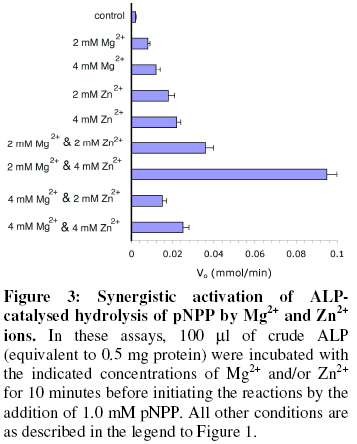
of Biochemistry, University of Ilorin, P.M.B. 1515 Ilorin, Nigeria Received 19 March 2007 Code Number: bk07007 Abstract The interactions of Mg2+ and Zn2+ ions in the activation of non-specific tissue alkaline phosphatase were investigated using crude extracts of rat kidney. Activation of alkaline phosphatase by the metal ions was accompanied by changes in the kinetic parameters of p-nitrophenylphosphate hydrolysis. The results suggest some synergistic interactions between Mg2+ and Zn2+ ionsin promoting the hydrolysis of p-nitrophenylphosphate by alkaline phosphatase. The results show that assays of alkaline phosphatase activity in homogenised tissue samples will give better responses if both Mg2+ and Zn2+ ions are included in the reactions Keywords:Alkaline phosphatase; kinetics; Enzyme-cofactor interaction; synergism INTRODUCTION The roles of metal ions in metalloenzymes include direct participation in catalysis, stabilization of protein structure and regulation of enzymatic activity. Membrane alkaline phosphatase (ALP) is a metal-containing enzyme that serves as a good model for the study of metal ion interactions in enzyme catalysis. Native E. coli ALP contains three metal ion binding sites (two Zn2+ sites and one Mg2+ site), and studies on their roles and interrelationships have provided some insights into the mechanism of the enzyme1. E. coli ALP is a zinc metalloenzyme, which can be activated by magnesium ion2. Removal of the Zn2+ leads to loss of catalytic activity while its replacement by other divalent cations (Mn2+, Co3+, Cd2+, and Cu2+) resulted in lower maximal activity2. It has been shown that while low concentrations of Mg2+ stimulated the refolding of E. coli ALP, high concentration actually inhibited its reconstitution into the active form3. This suggests that Mg2+ mediates stabilization and destabilization of the catalytically active structure of ALP at low and high concentrations respectively4,5. In E. coli ALP, Mg2+ is thought to have a regulatory effect on the expression of catalytic activity and maintenance of structural integrity of the enzyme1. The specific binding of Mg2+ to apo-ALP depends on both pH and the cooperative effects of Zn2+ binding. Mg2+ alone does not confer catalytic activity on ALP, but it does regulate the Zn2+-induced restoration of activity and perhaps, structural integrity of the metal-binding loci1,6. Mammalian ALPs are glycoproteins that are present as different isoenzymes in several tissues including bone, intestine, kidney, and placenta. Like their E. coli counterpart, they are zinc-metalloenzymes that can be activated by Mg2+ ions; both ions being essential for catalysis and structural stability. The human genome, like other mammalian systems contains four ALP loci, one coding for the tissue nonspecific ALP (TNAP) expressed in a variety of organs such as liver, bone, kidney, etc. and three tissue-specific ALP (TSAP) genes coding for the intestinal AP (IAP), placental ALP (PLAP), and germ cell ALP (GCAP). TNAP shows approximately 50% sequence similarity with the TSAP isozymes. Mammalian ALPs show 25-35% sequence identity with the E. coli enzyme in those regions of the protein assuming a-helix and b-strand secondary structures, and critical for catalysis. The catalytic residues, i.e. Asp91, Ser92, Arg166, and ligands coordinating the divalent metal ions (Zn2+ and Mg2+) are all conserved2. These structural similarities suggest that mammalian ALPs may catalyse hydrolysis of phosphate monoesters via a similar mechanism as the E. coli enzyme. Structural and functional aspects of the role of divalent cations have been extensively studied in E. coli ALP. Such studies have been facilitated by the availability of several X-ray structures of E. coli ALP. In contrast, less direct information is available on the structure and mechanism of mammalian ALPs. Owing to the practical importance of assaying tissue ALP activities in applied biochemical studies involving use of animals7 and the mechanistic interest of the subject, we have been studying the effect of cofactors and inhibitors on the catalytic properties of mammalian ALPs5,8. Kinetic analysis shows that high concentrations of Mg2+ inhibit tissue non-specific ALP in crude extracts by decreasing Vmax of pNPP hydrolysis5. Here we report a synergistic interaction of Mg2+ and Zn2+ ions in the activation of tissue non-specific ALP. The implications for the determination of alkaline phosphatase in crude tissue extracts are discussed. MATERIALS AND METHODS Reagents, chemicals, and enzyme source ALP substrate, the sodium salt of p-nitrophenylphosphate (pNPP) was obtained from Sigma Chemical Company, St. Louis, US. MgCl2 and ZnCl2 were products of British Drug House, UK. All other chemicals used in this study were of high quality research grade. The tissue non-specific ALP used here was obtained from rat kidney. Fresh kidney tissue from Rattus Norvegicus was homogenized in 0.25 M sucrose solution at 4 °C and the crude homogenate was centrifuged at 4000 rpm for 20 minutes at the same temperature. The supernatant was collected and dispensed in Eppendorf tubes and stored frozen. Fresh aliquots were used each day for the experiments described in this report. It is generally believed that to perform a large number of kinetic measurements under constant conditions it is more important to have large stabilized enzyme preparations than small preparations with a higher activity9. Thus, we did not attempt further purification since the enzyme is unusually stable even in impure crude preparations10. The key experimental results obtained with the crude ALP extract were validated with homogenous calf intestinal ALP obtained from New England Biolabs. Determination of alkaline phosphatase-catalysed hydrolysis of pNPP Alkaline phosphatase activity was measured by the rate of hydrolysis of appropriate concentrations of p-nitrophenylphosphate (pNPP) at 25 °C in 0.1 M Na2CO3/NaHCO3 buffer, pH 10.1 as previously described9. Enzyme activity is expressed as the mmol of p-nitrophenol released per minute. Reaction mixtures containing the enzyme source and buffers in the presence of the appropriate metal ions were kept at 25 °C for 10 minutes. In all assays, 100 ml of the crude enzyme preparation equivalent to 0.5 mg total protein were added to catalyse the hydrolysis of pNPP. Reactions were initiated by the addition of the appropriate concentration of the substrate, pNPP. Incubation was allowed for 10 minutes before stopping the reaction by the addition of 0.1 M KOH. The absorbance was read at 400 nm against a blank of the buffered substrate on a Specronic-21 UV-Vis spectrophotometer and the corresponding activities recorded. All measurements of reaction rate were performed in triplicate. Protein concentration was determined using Biuret method11. RESULTS Activation of alkaline phosphatase by Mg2+ and Zn2+ An analysis of the roles of Zn2+ and Mg2+ ions in the activation of ALP was carried out by investigating the kinetics of pNPP hydrolysis in the presence of 0, 2, 4 and 8 mM of the two metal ions separately (Figure 1). In experiments to determine kinetic constants, activity was monitored by measuring the absorbance change at varying substrate concentrations (0.2–2 mM pNPP) in the presence of varying metal ion concentrations. The effects of Mg2+ and Zn2+ on the Km for pNPP and Vmax values were obtained from Lineweaver-Burk plots (Table 1).Within the range of substrate concentration examined, the hydrolysis of pNPP by ALP followed saturation kinetics (Figure 1A). Kinetic analysis showed that a progressive decline in activation of ALP occurred as Mg2+ concentration was raised from 2 mM to 8 mM as observed by the progressive decrease in the maximum reaction rate Vmax. The activation of alkaline phosphatase by Mg2+ was also accompanied by a biphasic change in the Michaelis constant (Km). Increasing Mg2+ concentration from 2 mM to 4 mM resultedin a decrease in the Km for pNPP. However, the Km was higher at 8 mM Mg2+ concentration. The activation of ALP by Zn2+ followed a pattern similar to that of Mg2+ in affecting both the Vmax and the Km (Figure 1B). However, the effect of Zn2+ on the Km for pNPP did not show the biphasic pattern seen with Mg2+. There was a consistent decrease in the Km value as the Zn2+ concentration was increased from 0 to 8 mM (Table 1). Pre-incubation of alkaline phosphatase with Mg2+ and Zn2+ The results shown in Figure 1 and Table 1 indicate that Mg2+ ion at higher concentrations inhibit pNPP hydrolysis. This could be due to occupancy of the Zn site that is essential for catalysis by the higher concentration of Mg2+. Hence, we sought to investigate this possibility by determining how pre-incubation of the enzyme with Zn2+ and Mg2+ ions prior to the addition of the substrate affects catalytic activity. Each metal ion (4 mM) was added to the appropriate amount of the enzyme for different incubation periods (0, 5, 10, 15, 30 and 60 minutes) before adding 1.0 mM pNPP for 10 minutes. Figure 2 shows the effect of separate pre-incubation of ALP with 4 mM Zn2+ and 4 mM Mg2+ on phosphatase activity. The results show that the effects of Zn2+ and Mg2+ in activating ALP were time-dependent. The activity of ALP increased progressively when pre-incubated with Zn2+ from 0 to 60 minutes. For Mg2+ however, the activity of ALP increased progressively with time of pre-incubation up to 10 minutes after which a progressive decrease in activity was observed. Synergistic interaction of Mg2+ and Zn2+ on alkaline phosphatase activity Results presented in Figures 1 and 2 show that both Mg2+ and Zn2+ ions are required for ALP activation. Hence, we sought to determine if any synergistic interactions exist between both cofactors in the activation of ALP, and to determine the optimal concentrations of both ions for full activity. The effects of Mg2+ and Zn2+ ions on ALP activity were investigated by determining the rate of hydrolysis of 1.0 mM pNPP under different conditions. The conditions are: Control (0 mM Mg2+, 0 mM Zn2+), 2 mM Mg2+, 4 mM Mg2+, 2 mM Zn2+, 4 mM Zn2+,2 mM Mg2+ & 2 mM Zn2+, 2 mM Mg2+ & 4 mM Zn2+, 4 mM Mg2+ & 2 mM Zn2+, and 4 mM Mg2+ & 4 mM Zn2+. Significant differences between the rates of pNPP hydrolysis obtained under the different conditions were determined at 5% confidence level using the Duncan’s Multiple Range Test12. The results of this experiment are shown in Figure 3 and the pattern indicates a synergistic interaction between the two metal ion cofactors. DISCUSSION It has been established that E. coli alkaline phosphatase contains three classes of metal binding sites in which Zn2+ occupies the catalytic and structural sites, while Mg2+ ions are bound at the regulatory site1,13. The activation of ALP-catalysed hydrolysis of pNPP observed in this study by the two metal ions is exerted via both Vmax and Km effects. The decline in activation at 8 mM Mg2+ observed here is consistent with the observation in our earlier study that high concentration of Mg2+ inhibits the non-specific ALP from rat liver5. It was shown that Mg2+ acts as an activator within optimal concentrations but became inhibitory at higher concentrations. One possible explanation for this set of results is that excess Mg2+ ions displaced Zn2+ from the catalytic site since both metal ions can bind to the same site. The effect of Mg2+ concentrations on the Michaelis constant (Km) suggests that Mg2+ activates kidney ALP by increasing the affinity of the enzyme for pNPP. This agrees with the report that the binding of Mg2+ to kidney ALP promotes a conformational change9, which causes the displacement of the substrate into the vicinity of zinc-coordinated water molecule, thus leading to increased affinity of the enzyme for its substrate. The concentration-dependent activation of kidney ALP by Zn2+ agrees with the findings of Bosron et al. (1977)1. The observation that Zn2+ affects Km may suggest that Zn2+ induces its activation effect on ALP through its interaction with the free enzyme. The observed increase in Vmax due to increasing Zn2+ concentration most likely reflects the catalytic requirement for the metal ion in phosphate ester hydrolysis14. Pre-incubation of ALP activity with the metal ion cofactors led to increase in the hydrolysis of ALP. Hence, the metal ions probably mediate certain time-dependent effects that result in stimulation of activity. Such progressive activation is characteristic of conformational changes at or around the active site and may involve optimizing substrate binding or the actual steps of catalysis. A structural role for Mg2+ has been proposed in earlier studies5,13. The inhibition that results after prolonged incubation of ALP with Mg2+ may be due to displacement of Zn2+ from its binding site on the enzyme15. Earlier works showed that ALP contains two Mg2+ binding sites and four Zn2+ binding sites in its active site14,16. Thus, the synergistic interaction observed between Mg2+ and Zn2+ in activating ALP (Figure 3) suggest that both metal ions are required in a defined optimal ratio. The scope of this study is not sufficient to ascertain the exact ratio of the two metal ions required for optimal hydrolase activity. Mg2+ is usually employed as the only cofactor in routine ALP assays in clinical chemistry as well as in toxicological investigations7,8,14. However, the results obtained in this study suggests that better responses are likely to be obtained in such assays if carried out in the presence of both Mg2+ and Zn2+ at appropriate concentrations. REFERENCES
© 2007 Nigerian Society for Experimental Biology. All rights reserved. The following images related to this document are available:Photo images[bk07007f3.jpg] [bk07007t1.jpg] [bk07007f2.jpg] [bk07007f1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}