 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
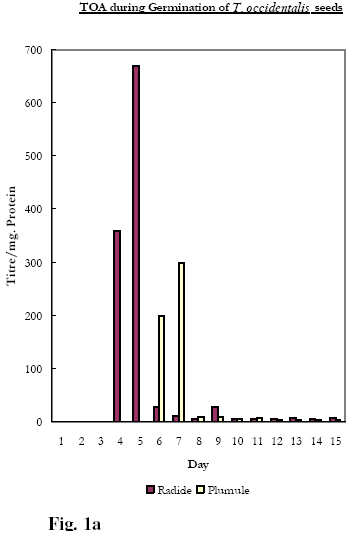
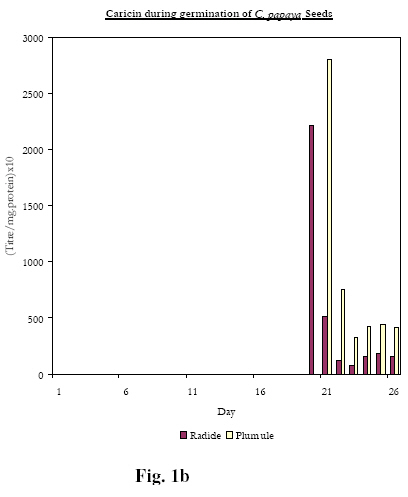
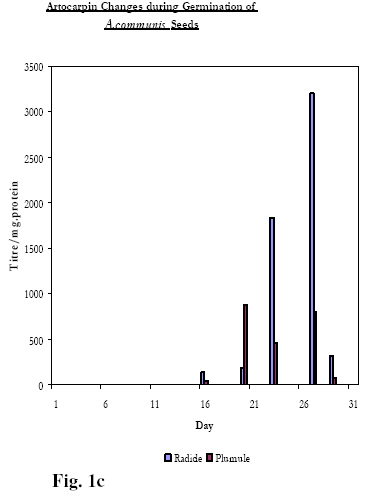
Biokemistri, Vol. 20, No. 1, June, 2008, pp. 11-15 Lectins, Mitogenicity and Seed Germination: A Comparative Study with the Seeds of Telfairia occidentalis (Hook, F.) (Curcurbitaceae), Carica papaya (Linn) (Caricaceae) and Artocarpus communis (J.R. & G. Forst) (Moraceae) Rachel A. Togun1*, BeatriceO. Emma-Okon1 and Akintunde A. Aboderin2 1College of Health Sciences, Obafemi Awolowo University, Ile-Ife, Osun State, Nigeria Received 24 May 2007 Code Number: bk08002 Abstract The fate of lectins contained in the seeds of T. occidentalis, C. papaya and A. communis have been followed during the germinative process. Under the same culture conditions, the emergence of both the radicle and the plumule (the 3rd and 5th day; 18th and 20th and 12th and the 15th day respectively for T. occidentalis, C. papaya and A. communis) was preceded, 24 hours earlier, by the appearance of lectin in the developing tissues. The data suggest that the mitogenic lectins: Telfairia occidentalis Agglutinin (TOA) and Artocarpin accelerates the process of germination relative to that observed for seeds (C. papaya) containing the non-mitogenic lectins, Caricin Keywords:Lectins, Mitogenicity, Seed Germination INTRODUCTION In an earlier study, the fate of a mitogenic lectin from the seeds of Telfairia occidentalis Agglutinin, (TOA) was monitored during germination as a means of identifying the possible role of this lectin in the plant1. The results from the study suggested that one of the roles of the mitogen is to stimulate the appearance of the radicle and the plumule. Given that, to our knowledge, this was the first time such a specific physiological role had been ascribed to any lectin; it was of interest to broaden the scope of the work by inquiring if the observation implied a necessary dependence of the effect of the lectin on its mitogenicity. In this report, we have extended the study to the germination of two lectin-containing seeds whose characteristics have been reported upon in previous work from our laboratories. Firstly, Carica papaya seed contains a family of structurally related lectins (Caricin)2. The agglutinin activity is inhibited exclusively by lactose and does not stimulate the incorporation of 3H-Thymidine and 35S-Methionine into lymphocytes from pig, mouse, and human blood samples under conditions in which Con A, PHA as well as the Agglutinins from Telfairia occidentalis, Tetracarpidium conophorum and Artocarpus communis were mitogenic2,3,4. C. papaya, it must be observed, belongs to the family, Caricaceae, with only one genus Carica which is found in the west tropical sub region of Africa. Caricaceae and Curcurbitaceae, to which T. occidentalis belongs, are two families in the order Curcurbitales. Mitogenic TOA, like Carica agglutinin, was inhibited by β-galactosides generally, e. g. Lactose, but also by Galactose. Secondly, the seeds of A. communis, a plant belonging to the family Moraceae contain a mitogenic haemagglutinin (Artocarpin) that is specifically inhibited by the Melibiose, (6-O-α-D-galactopyranosyl-D-glucose), but neither by Lactose nor Galactose4. Similar to our earlier study on the germination of the seed of T. occidentalis 1, we compare the distribution of the non-mitogenic C. papaya seed lectin during germination and plant development to those of the mitogenic lectins from the seeds of T. occidentalis and A. communis MATERIALS AND METHODS: The seeds of T. occidentalis, C. papaya and A. conophorum were collected from the mature, ripe fruit pods from the plants in the campus of Obafemi Awolowo University, Ile-Ife, Nigeria during the fruiting seasons. The seeds to be planted were prepared as previously described 1. Briefly, large labelled plastic bowls were punched at the bottom to allow for effective drainage, and filled to two-third levels with humus soil. After wetting the soil, several washed seeds of each plant, according to the label, were sown just below soil surface levels. Watering was done daily until the experiment was over. The pots were kept in an open space where they were exposed to sunlight and other natural conditions. Samples were harvested in duplicates daily or every other day starting from the day after the planting, i.e. day 1, for analysis. Seeds that were not planted at all represent the day 0 samples. Each sample of C. papaya seeds contained several of the very tiny seeds enough to weigh about 0.5 mg, while the samples of T .occidentalis and A conophorum consisted of just one seed or plant. As from when germination of seeds started and thereafter, the lengths of the plumule and radicle were measured. Using a sharp scalpel, each plant was separated into its different components- radicle, plumule and cotyledon or later, cotyledon, root, stem, and leaves. Each component was ground, and extracted independently of the others. Preparation of crude extracts Dried seeds (Day 0) as well as swollen seeds and different parts of germinating plants were ground in five volumes of phosphate buffered saline (PBS, 0.15 M sodium phosphate buffer, pH 7.2, containing 0.15 M NaCl) and stirred overnight at 40C. Each suspension was centrifuged at 3000 rpm at room temperature and the resultant supernatant was stored frozen until required. Haemagglutination assay Haemagglutination activity of extracts of seeds and different parts of germinating seedlings and plants was determined as earlier described 5. Haemagglutination experiments were performed in V-shaped microtitre plates. Lectin preparation (100µl) was serially diluted in doubling dilutions in the wells of the microtitre plates with PBS (pH 7.2). This process was carried out for two complete rows (or 24 wells) for each sample. 100 µl of human red blood cells previously fixed with glutaraldehyde (1% v/v) or Formaldehyde6 was added to each of the wells. The plates were incubated at room temperature for 2- 24hours to allow for the agglutination of erythrocytes. The titre of the lectin was taken as the reciprocal of the last dilution showing agglutination of erythrocytes. Protein concentration determination Protein concentrations were determined according to a modification of the Lowry technique7. Mitogenicity Tests The ability of a range of concentrations of T. occidentalis, C. papaya and A. conophorum seed extracts to stimulate human and murine lymphocytes was assessed by determining the level of 3H-Thymidine and 35S-Methionine uptake as previously described5 RESULTS A full description of the stage-by-stage changes observed in the morphological structure of planted seeds of T. occidentalis during germination and the accompanying lectin distribution, in particular the cotyledon, radicle and plumule, has been provided in the earlier work1. These data as well as equivalent results for the seeds of C. papaya and A. communis have been summarised in Table 1 and Figures 1a, 1b, 1c. These data show that, in terms of the morphological changes at the onset of germination, the seeds of the three lectin-containing plants show the same tendencies: viz.: as is the case for dicotyledonous seeds, the radicle appears prior to the appearance of the plumule. In addition to this, the data also demonstrate that, as shown earlier for the seeds of T. occidentalis1, the appearance of these plant tissues is preceded by the transient presence of lectin, which rapidly disappears following the proper establishment and maturity of the tissues. Figures 1a, 1b, 1c, in which the fates of TOA, Caricin and Artocarpin during the course of germination in addition to the data in Table 1, show that there are however differences. C. papaya seeds containing the non-mitogenic Caricin are the slowest to germinate, with the radicle and plumule emerging on the 18th and 20th day after planting, the corresponding figures for T. occidentalis and A. communis seeds being 3rd and the 5th day and the 12th and the 15th day respectively. Table 1: Time required for mitogenic and non-mitogenic seed germination
DISCUSSION There are three possible reasons that might account for the observed behaviour of the seeds of the three plants in this comparative study. The first is agglutinating power and sugar specificity of the lectins. The second is taxonomy; and the third is mitogenicity. It is unlikely that agglutinating power by itself can sufficiently account for our observations. For while it is true that Caricin is a relatively weak agglutinin2, with maximum titres of the order of 28, as a consequence of which the stimulation of germination might be delayed; and TOA5, with titres of 212 and Artocarpin4 with titres of 216, possess stronger agglutinating powers, it would by these same facts be expected that the emergence of the tissues in A. communis should have preceded those of T. occidentalis. This clearly is not the case. Furthermore, of the three agglutinins, TOA is the only one with a broad specificity for Galactose and, in general, β-galactosides such as Lactose. Artocarpin and Caricin on the other hand have very narrow specificities involving respectively 6-O-α-D-galactopyranosyl-D-glucose (Melibiose)4 and Lactose2. The identity of which β-galactosides might be fulfilling important functions during germination has not been addressed in this communication and remains a subject for further inquiry. In a similar vein, taxonomy may not a major factor in trying to explain our data. T. occidentalis and C. papaya both belong to the family Curcurbitales. In spite of this, not only is mitogenecity not a common property of TOA and Caricin, the behaviour of the seeds during germination is also different. A. communis on the other hand belongs in the family Moraceae; the seeds contain a mitogenic lectin, and, subject to the observation above on sugar specificities, their behaviour during germination is closer to that of T. occidentalis seeds. It is mitogenicity, a common feature of both TOA and Artocarpin and is lacking in Caricin, which we believe may be an important factor to account for our results and which will be in conformity with our earlier data1 on T. occidentalis seed germination. Brill et al.8 working on the expression of MsLEC-1 and MsLEC-2 antisense genes in alfalfa plant lines have also suggested that a role for lectins in plants is in embryogenesis, vegetative and reproductive development. This study, in a comparative manner, assessed the role of non-mitogenic and mitogenic lectins of Carica papaya, Telfairia occidentalis, and Artocarpus communis in the germination and growth process of plants. The data that has been presented confirmed our earlier finding that Telfairia occidentalis mitogenic lectin played a role in the multiplication of tissues to be differentiated as shown by the behaviour of another mitogenic lectin- containing plant, Artocarpus communis. It also showed that the non mitogenic lectin- containing plant, Carica papaya also behaved like the mitogenic plants after germination, but that the germinative process was delayed, while the lectin-containing cotyledon later transformed to leaves. REFRENCES
© 2008 Nigerian Society for Experimental Biology. All rights reserved. The following images related to this document are available:Photo images[bk08002f1b.jpg] [bk08002f1a.jpg] [bk08002f1c.jpg] | ||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}