 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
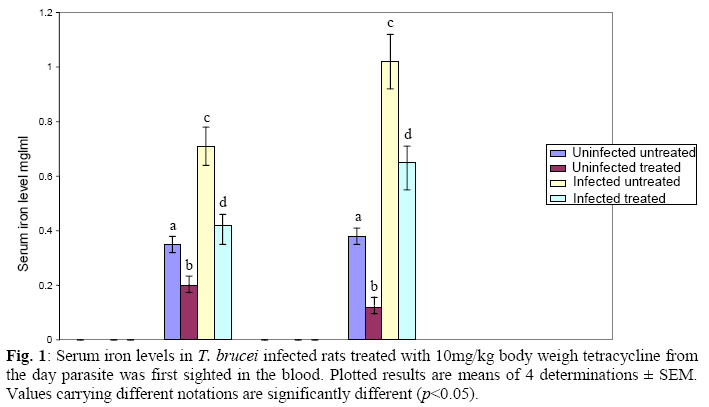
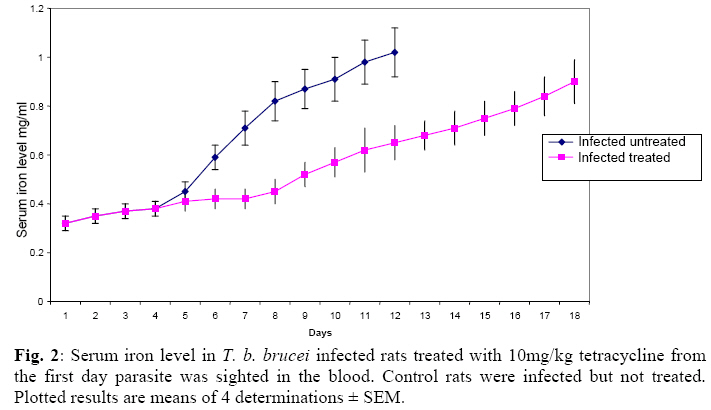
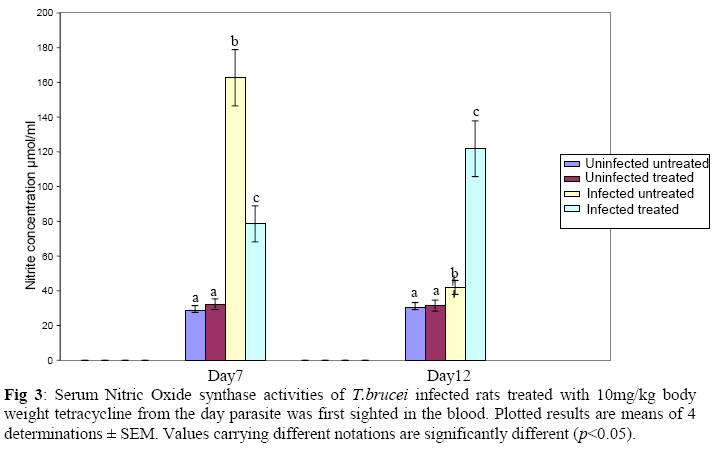
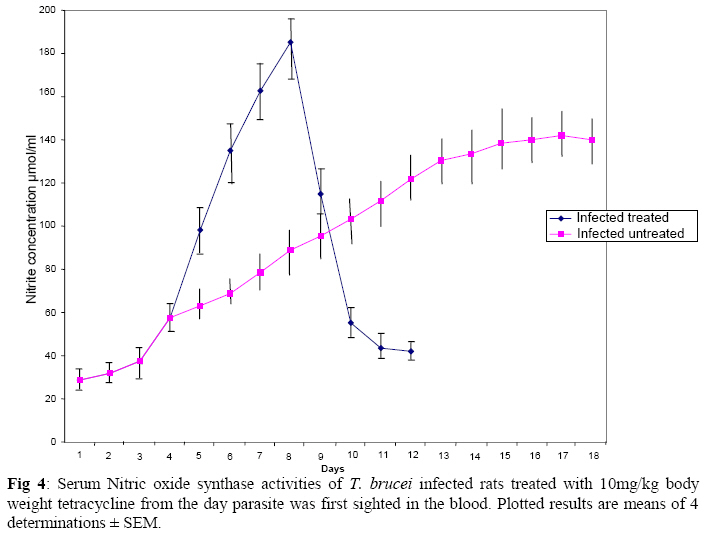
Biokemistri, Vol. 21, No. 1, June, 2009, pp. 41-51 Serum Iron and Nitric Oxide Production in Trypanosoma brucei Infected Rats Treated with Tetracycline Justine T. Ekanem, Titilayo O. Johnson* And Elizabeth A. Balogun Department of
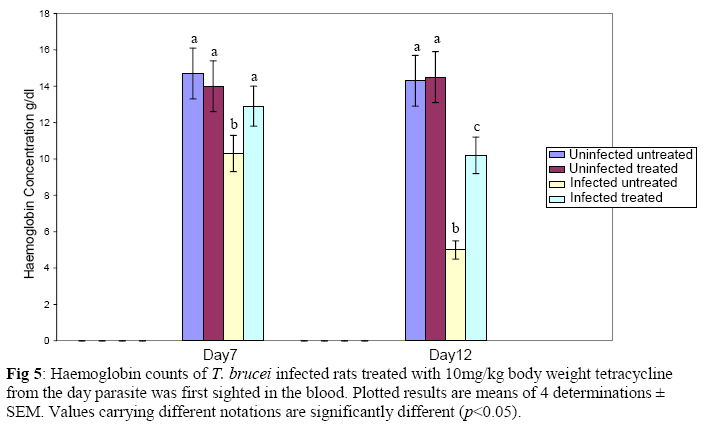
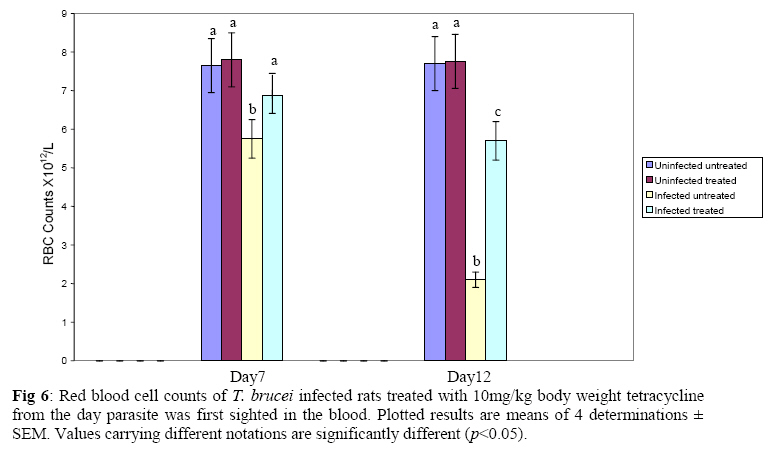
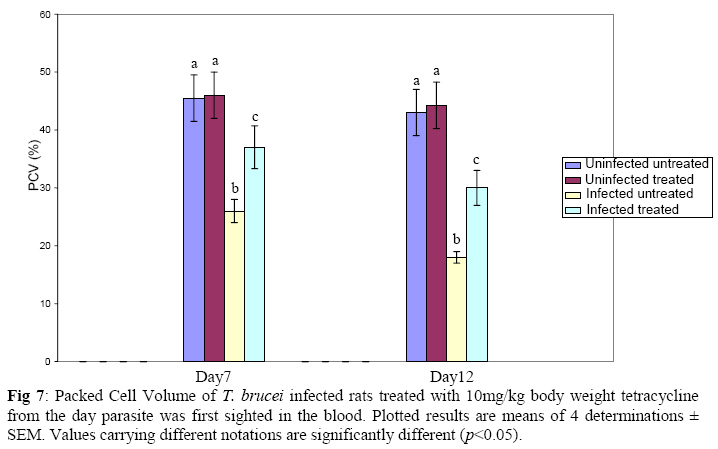
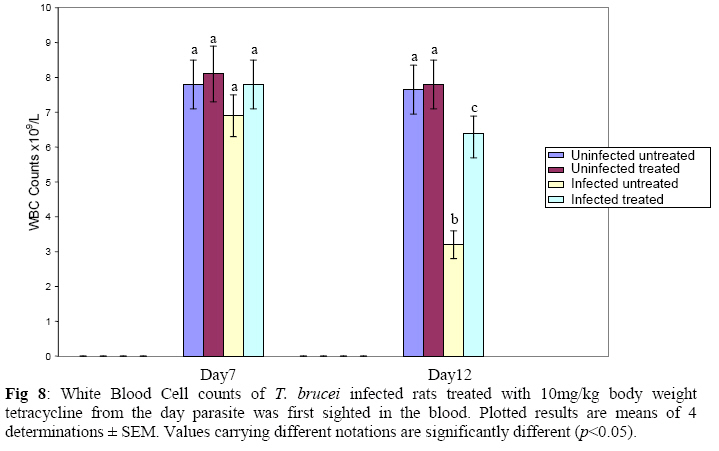
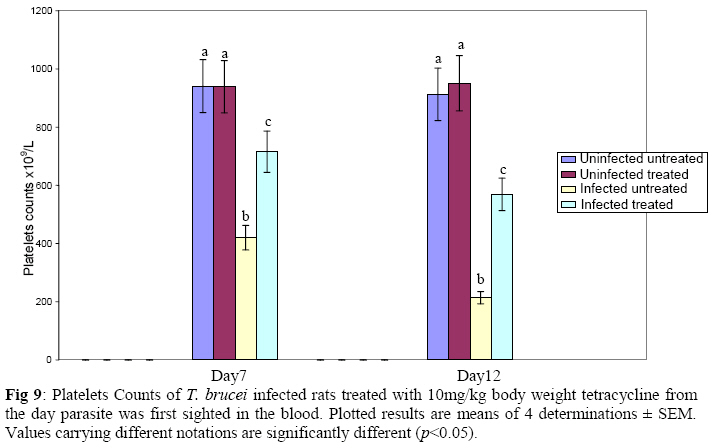
Biochemistry, University of Ilorin, Ilorin, Kwara State, Nigeria Received 2 April 2009 Abstract The effects of tetracycline on serum iron, nitric oxide production, and haematological parameters in Trypanosoma brucei infected rats were investigated. T. brucei infected and uninfected rats were treated with tetracycline intraperitoneally at 10mg/kg body weight. Four rats each from various groups were treated daily from the first day parasites were sighted in the blood and continued until the animals died. Infected rats, treated and untreated, were sacrificed daily for the serum iron levels and nitric oxide synthase activities. For haematological parameters, infected and uninfected but treated rats were sacrificed on days 7 and 12 along with untreated rats. Results showed that tetracycline brought about a significant reduction in the serum iron status and a modulation of nitric oxide synthase activity of T. brucei infected rats. Red blood cell counts, haemoglobin concentrations, packed cell volumes, white blood cell and platelet counts were also significantly higher in infected as a result of treatment with tetracycline. The results from this study suggest that, tetracycline which has previously been reported as a possible cheap drug in the management of African trypanosomiasis, probably exerts its trypanocidal action through reduction in serum iron status, which in turn modulates nitric oxide synthase activity, thereby reducing the severity of anaemia in infected rats Keywords: Nitric oxide synthase, tetracycline, African sleeping sickness INTRODUCTION Tetracycline, a broad spectrum antimicrobial drug with iron chelating property was found in previous studies to be effective against both the early and late stages of the infection in rats and the trypanocidal action was suggested to be as a result of iron-chelation1,2. Iron is an essential nutrient for living cells that play important roles in cellular process such as synthesis of DNA, RNA and proteins, electron transport, cellular respiration, cell proliferation, and regulation of gene expression3. Both iron deficiency and iron over load can be pathogenic. Iron displays carcinogenic activity and the ability to favour tumor growth, due to catalytic effects on formation of hydroxyl radicals, promotion of tumor cell multiplication and suppression of activity of host defense cells4. As a result of this, iron chelation has been successfully tested in the clinical setting as a component of anticancer therapy5. Iron chelation therapy has recently been suggested to be a potent means of controlling trypanosomes in vivo through the inhibition of ribonucleotide reductase, the iron-requiring cell cycle dependent enzyme that plays a central role in cell division and proliferation6. Another importance of this therapy, which is of interest in this study, is the modulation of the activity of nitric oxide synthase- a cytokine inducible enzyme which catalyses the formation of the antiparasitic free radical- nitric oxide in macrophages7. African trypanosomiasis or Sleeping sickness is a parasiticdisease in people and in animals. It is caused by protozoa of genus Trypanosoma and transmitted by the tsetse fly; the disease can lead to damage of the central nervous system and a sleeping sickness syndrome in the sub-Saharan region. Some of the fatal consequences of African trypanosomiasis are anaemia and tissue damage8. A number of toxic factors have been found in the crude lysates of Trypanosoma species9 purified trypanosomes have also been found to release several proteinaceous substances not necessarily proteases which in addition to proteases might be responsible for haemolysis and the consequent anaemia in African trypanosomiasis10. Another possible mechanism for the pathogenicity of anaemia is the impairment of bone marrow function. Studies have shown that bone marrow is severely stressed during infections with T. brucei and T. musculi11,12. Impairment of bone marrow function and iron reutilization in cattle infected with T. congolense has also been reported13. Erythrocyte destruction and tissue damage could also be due to factors produced or induced by the presence of trypanosomes in the blood14. The variable surface glycoproteins (VSG) of the blood stream form of trypanosomes are strongly immunogenic and elicit high level of IgM antibodies. Raised IgM levels increase osmotic pressure, erythrocyte sedimentation rate and blood viscosity. Circulating immune complexes activate kallikrein, kinin, complement and blood coagulation systems, which in turn lead to increased vascular permeably, oedema, inflammation and tissue damage15. Over production of nitric oxide by inducible nitric oxide synthase (iNOS) in macrophages is another mechanism proposed for the pathogenicity of anaemia and tissue damage in African trypanosomiasis7. In this study, we have assessed the possible beneficial effects of iron chelation therapy and its modulation of nitric oxide synthase activity which may in turn regulate the severity of anaemia in trypanosome infections. MATERIALS AND METHODS Experimental rats Male Wistar rats, weighing approximately 200g obtained from the Department of Biochemistry, University of Ilorin, Ilorin were kept in rat cages and fed with grower’s mash and water ad libitum for at least one week before the commencement of the experiment. Trypanosomes Bloodstream forms of Trypanosoma brucei brucei (virulent stock) obtained from the Nigerian Institute of Trypanosomiasis Research, Vom and maintained by repeated passages in albino rats until needed. Administation of drug Trypanosoma brucei infected and uninfected rats were treated with tetracycline intraperitoneally at 10mg/kg body weight. Infected and uninfected rats were administered daily with tetracycline from the first day parasites were sighted in the blood and continued until the infected animals died. Inoculation of rats Experimental rats were infected with T. b. brucei. Highly infested blood as observed under light microscope was obtained from the tail of an infected rat directly into phosphate saline glucose (PSG), pH 7.5 without anticoagulant. At 105 trypanosomes per ml, 0.5ml of suspension was passaged into the experimental albino rats by intraperitoneal injection. Collection of blood Blood was collected from the jugular veins of the animals into sample bottles with and without anticoagulant. The blood with anticoagulant was kept at 40C until required for haematological studies, usually not more than 48hrs. Clear serum was obtained from the other blood sample by centrifugation of the clear part of blood after clotting at 3,000g for 15min and kept frozen until required, usually after 24hrs. Determination of serum iron level The serum iron level was determined spectrophotometrically by the O-phenanthroline method16. Ferrous iron reacts with o-phenonthroline to form an orange-red complex (C12 H8 N2) Fe2+. The colour intensity is independent of the acidity in the pH range 2-9 and is stable for long periods. Ferric iron was reduced with hydroxylammonium chloride and the iron-phenanthroline complex as the perchlorate was extracted with nitrobenzene and measured at 515 nm against a reagent blank. Determination of nitric oxide synthase activity The activity of nitric oxide synthase was determined by the estimation of nitrite concentration in the serum sample using the Greiss reaction assay17. The measurement of nitrate/nitrite concentration or of total nitrate and nitrite concentration (NOx) is routinely used as an index of NO production. In this method, nitrite was first treated with a diazotizing reagent, i.e., sulfanilamide (SA), in acidic media to form a transient diazonium salt. This intermediate was then allowed to react with a coupling reagent, N-naphthyl-ethylenediamine (NED), to form a stable azo compound. Haematological parameters Red blood cells, white blood cells and platelets counts were provided by a haemocytometer. The packed cell volume of whole blood sample was determined for each sample using capillary tube on a microhaematocrit centrifuge at 10,000xg for 5 min and read on a microhaematocrit reader. 20μl of whole blood was properly mixed with 4.0ml Drabkin solution, a mixture of hydrated potassium cyanide and potassium ferrocyanide. It was then allowed to stand for 10minutes. The optical densities of the standard and samples were obtained at 540nm and the haemoglobin concentration was calculated. RESULTS Serum iron level Tetracycline caused a significant decrease in the levels of iron in the serum of uninfected treated rats when compared with the uninfected untreated ones. The levels of iron increase rapidly in the serum of infected untreated rats but the concentrations were lower in the infected treated rats (Figs 1 & 2). Nitric oxide synthase activity There was no significant difference in the NO synthase activity of uninfected treated rats compared with the untreated ones on days 7 and 12. In the infected untreated rats, activities of NOS increased significantly from days 1 to 8 after which there was a drop up to day 12. The enzyme activities in the infected treated rats were lower compared with those of the untreated ones from days 5 to 8, but higher on days 9 to 18 (Figs. 3 & 4). Haematological parameters There were no significant differences in the HB, RBC, PCV, WBC and platelet counts of uninfected treated and untreated rats on days 7 & 12. Marked reductions in the levels of these parameters were observed in the infected untreated rats on days 7 & 12, but there were improvements in the infected treated rats (Figs 5, 6, 7, 8, 9). DISCUSSION The continuous increase in the level of iron in the serum of infected untreated rats in this study is likely to be as a result of iron loss from erythrocytes into the plasma of infected animals. Studies have shown that African trypanosomiasis is accompanied by a severe drop in packed cell volume (PVC) and red blood cells (RBC) counts18-21 as a result of erythrocytes destruction. The results obtained in this study also showed a reduction in RBCs, PVC and haemoglobin concentration along with the increased level of serum iron. The loss of iron into the plasma is related to anaemia observed in infected rats22-24. Accumulation of “free” iron is injurious to the animal because it can catalyze the formation of hydroxyl radical from hydrogen peroxide25. The hydroxyl radical is highly reactive, and attack lipids, protein and nucleic acids26. The initial reaction with each of these molecules is the formation of peroxides (lipid peroxides) that can interact with other molecules to form cross links. This process leads to further destruction of red blood cells and damage to cells in other organs of the body26. In the infected treated rats, the levels of iron in the serum were reduced compared with those of untreated rats’ probably as a result of iron chelation. This property of tetracycline has been suggested in previous studies to contribute to its trypanocidal action because when infected rats were treated with tetracycline iron complex, there were little or no improvements in the parasitaemia and lifespan of the rats, unlike when tetracycline alone was used1. The iron chelating property of tetracycline has also been demonstrated by Grenier et al27, they suggested that this property contributes to its antimicrobial property against actinobaccillus actinomycetemcomitan since when tetracycline was saturated with Fe, its activity against the bacteria was inhibited27. Apart from inhibiting proliferation of parasites1, removal of excess iron through chelation by tetracycline could also possibly prevent iron mediated injury to cells thereby reducing the pathology of anaemia and tissue damage associated with the infection. The improved levels of haematological parameters observed in this study strongly support this suggestion. The initial increase in NOS activity of infected untreated rats observed in this study which corresponds to increased NO production is part of the initial immune responses of the animal after inoculation of parasites. The inducible NO (iNOS) is induced by cytokines, tumor cells and microbial product mainly in macrophages and generate large quantity of NO that have been implicated in antimicrobial and antitumor defense4,8,29. The tumoricidal action of macrophage derived NO is mainly due to inhibition of iron-containing enzymes and involves repression of DNA synthesis, mitochondrial respiration and the enzymes of the citric cycle in target cells30,31. In the case of African trypanosomiasis, there are evidences that NO production has little or no effects on proliferation of the parasites in vivo. Studies using mice deficient in inducible NOS showed that parasitaemia increased at the same rate in both iNOS deficient homozygous and heterozygous mice, and peak parasitaemia values were the same in both groups and that NO production depresses T-helper I-like responses generated during T. brucei infections, thus promoting survival of the parasite32. Previous studies have also shown a rapid increase in the parasitaemia of infected rats from day 1 of infection until the death of the rats1, and this does not correspond with the rapid increase in NO production observed in this study. The inability of NO to act against trypanosomes in vivo is therefore likely to be related to the excessive release of iron into the plasma during infections33,34 It has also been suggested that the NO catalyzed destruction of the iron-sulphur proteins requires exogenous ferrous iron. Therefore the excessive release of Fe, together with the over production of NO observed in this study possibly contribute to the reduction of RBCs, HB, PCV observed in this study. The sudden drop in the NOS activity observed in the serum of infected untreated rats from days 9 to 12 in this study might be due to increased level of iron which probably inhibits its production. It might also be as a result of increased serum and CSF levels of some cytokines as well as prostaglandin E235. In the infected treated rats, there is a reduction in the NOS activity from days 10-12 probably as a result of reduced level of parasites brought about by the effect of the drug, i.e. depriving the parasite of iron for effective proliferation. This reduction in NO production also limits its destructive effects on cells as shown by the improved haematological parameters observed for the infected treated rats. The reduced RBC counts, PCV and HB concentration in the infected untreated rats is an indication of anaemia, which is a well-recognized and inevitable consequence of trypanosome infection in domestic animals and man36. The bases of anemia are the increased rate of destruction of red blood cells. Proposed mechanisms include hemolysis as a result of erythrophagocytosis, hemodilution and depression of erythropoiesis37,38. Other reports have also shown that over production of NO during infections with T brucei plays a significant role in the anaemia. Impairment of bone marrow function is another possible cause of anaemia in T. brucei and T. musculi infection11,12. This suggestion also explains the reduction in WBC and platelets counts recorded for the infected untreated rats. A reduced WBC count (Leucopenia) result in increased risk of infection which means the infected animal is exposed to other infections. A decrease in platelet counts (thrombocytopenia) causes either internal or external hemorrhage. This could be another possible mechanism for the pathogenesis of anaema in trypanosomiasis. The increased level of haematological parameters in the infected treated rats is an indication of improved state of anaemia. REFERENCES
© 2009 Nigerian Society for Experimental Biology. The following images related to this document are available:Photo images[bk09006f5.jpg] [bk09006f2.jpg] [bk09006f1.jpg] [bk09006f9.jpg] [bk09006f3.jpg] [bk09006f4.jpg] [bk09006f6.jpg] [bk09006f8.jpg] [bk09006f7.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}