 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biokemistri, Vol. 23, No. 3, Dec, 2011, pp. 124-128 Original Article Suicide inactivation of horseradish peroxidase by excess hydrogen peroxide: The effects of reaction pH, buffer ion concentration, and redox mediation Sylvia O. Malomo1*, Raphael I. Adeoye1, Lateef Babatunde1, Ibraheem A. Saheed1, Martin O. Iniaghe2, and Femi J. Olorunniji1,3* 1Department
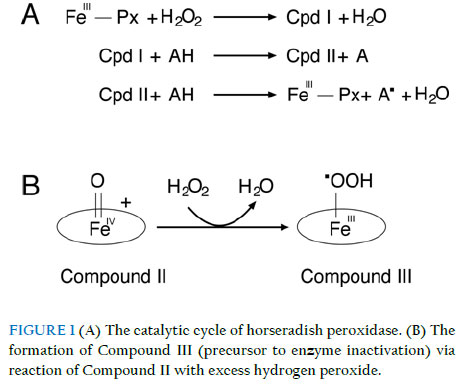
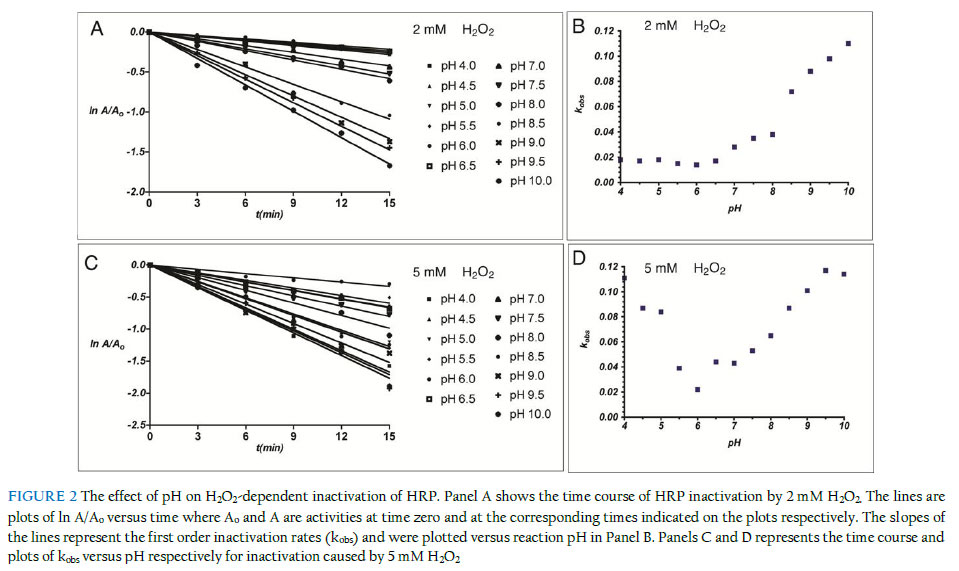
of Biochemistry, Faculty of Science, University of Ilorin, Ilorin, Nigeria; Received: 19 October 2011 Code Number: bk11017 ABSTRACT The inactivation of peroxidases by its oxidant substrate H2O2 limits the usefulness of these versatile enzymes. Here, we investigated the effect of reaction conditions on inactivation of horseradish peroxidase by excess H2O2. Inactivation was more pronounced at pH extremes, indicating that reactions in which the oxidation products induce significant changes in reaction pH could accentuate the loss of peroxidase activity. In reactions carried out in sodium acetate buffer, higher inactivation rates were observed when the buffer ion concentration was increased, an indication that peroxidase might be generating reactive radicals from the buffer molecules. Promethazine exerted a modest protective effect against inactivation; however, higher concentrations of the redox mediator caused a slight increase in inactivation, likely due to the formation of reactive promethazine radicals, which in turn attack the protein via a mechanism different from that caused by excess H2O2. These findings will help in defining the optimal reaction conditions that preserve the activity of the peroxidase molecules. INTRODUCTION Peroxidases have promising potential uses in industrial-scale catalytic conversion due to the combination of their catalytic efficiency and relative specificity of enzymes in general. However, the rate and efficiency of peroxidase-catalysed reactions are affected by the progressive inactivation of the enzyme by the substrate H2O2. This process, referred to as mechanism-based inhibition or suicide inactivation, limits the potential use of peroxidases in industrial-scale conversion processes. Despite the obvious value of peroxidases, their present commercial uses are limited, primarily by the low stability of peroxidases in the presence of their natural substrate, hydrogen peroxide. All hemeproteins, including peroxidases, are inactivated in the presence of catalytic concentrations of hydrogen peroxide. This process, which is described as a suicide inactivation, is especially important in the absence of reducing substrates, and its mechanism has not been fully elucidated (Valderrama et al., 2002). The catalytic mechanism of horseradish peroxidase described by Dunford (1982) is summarized in Figure 1A. The ground state ferric peroxidase (FeIII-Px) donates two electrons to H2O2 resulting in formation of a redox intermediate of the enzyme called compound I (CI), and H2O. Compound I consists of an oxoferryl protein cation radical, in which one of the oxidation equivalents exists as Fe (IV) while the other is a porphyrin-centred cation radical. Compound I reacts with the donor reductant substrate (AH) generating substrate free radicals and another redox intermediate, Compound II (CII). In Compound II, the oxoferryl species remains intact as it is in Compound I, but the cation radical is reduced. Another one-electron reduction of Compound II by a second donor reductant molecule leads to the regeneration of the ground state ferric enzyme and formation of a second equivalent of the substrate free radical. The molecular mechanism underlying H2O2-mediated inactivation of peroxidases is extraordinarily complex due to the multitude of reactions that can occur subsequent to the reaction of the haem iron with the hydroperoxide. Despite peculiarities among different hemeproteins, a common inactivation mechanism comprising several stages has been proposed (Valderrama et al., 2002). In the absence of substrate, or when exposed to high concentrations of H2O2, peroxidases show the kinetic behavior of a suicide inactivation, in which hydrogen peroxide is the suicide substrate that converts Compound II into a highly reactive peroxy-iron(III)porphyrin free-radical called Compound III (Figure 1B). Compound III is not part of the peroxidase cycle (Adediran, 1996; Huwiler et al., 1986), but it is produced under excessive exposure of protonated Compound II to oxidative species in a reaction partially mediated by superoxide free radical. ESR spin trapping and spectral analyses have demonstrated the occurrence of this species after the oxidative treatment of cytochrome c, horseradish peroxidase, prostaglandin H synthase, lignin peroxidase, and manganese peroxidase. Despite representing different structural groups, the kinetic models for the hydrogen peroxide mediated inactivation of horseradish peroxidase and ascorbate peroxidase are similar in that they are time dependent and show saturation kinetics. In both cases, the addition of a reducing substrate protected the enzyme from inactivation (Valderrama et al., 2002). The addition of excess substrate would preclude the suicide inactivation by competing with hydrogen peroxide for Compound II, as has been previously suggested. Once formed, Compound III might follow at least three alternative decomposition pathways. Due to the proximity of the bound peroxyl radical of Compound III to the porphyrin ring, it is likely that once formed, this reactive species would gain access to the tetrapyrrole structure and oxidize the porphyrin moiety. Secondly, Compound III might return to the ground state after catalyzing the oxidation of the surrounding protein, yielding an oxidized amino acid side chain group. Alternatively, the electron donor might be a substrate molecule, in which case the porphyrin moiety would be repaired, and a ground state enzyme would be regenerated. It is necessary to understand the factors that come into play in the inactivation process, in order to optimise the efficiency of the reactions in the face of competing enzyme inactivation. This study looks at the effects of buffer concentration, reaction pH, and promethazine, a known redox mediator on the catalytic conversion of ABTS by horseradish peroxidase, and its inactivation by excess H2O2. MATERIALS AND METHODS Reagents, substrates and enzyme Horseradish peroxidase and H2O2 were products of Sigma-Aldrich. Other chemicals and reagents used were of high quality research grade. The concentration of HRP, expressed as haem concentration was measured spectrophotometrically based on Soret absorbance at 428 nm by using a millimolar absorption coefficient per haem of 89 mM-1cm-1. Determination of enzyme activity HRP activity was determined by measuring the rate of oxidation of 2,2'-azino-bis(3 ethylbenzothiazoline-6-sulphonic acid) (ABTS) at 20 oC in a 3.0 ml reaction mixture containing 50 mM sodium acetate buffer, pH 5.4, 0.025 mM ABTS and 0.25 mM H2O2. Reactions were initiated by adding H2O2 as the last component of the mixture. The initial rate of formation of the ABTS oxidation product was measured at 414 nm. Desired concentrations of H2O2 were prepared daily from a 30% stock solution and determined spectrophotometrically using horseradish peroxidase as catalyst for I3- generation from I- (Cotton & Dunford, 1973). Concentrations determined this way were confirmed by absorbance measurements at 240 nm, taking E240 as 43.6 M-1cm-1. Kinetics absorbance measurements were performed with a UV/visible spectrophotometer model Camspec M105. pH dependence of inactivation kinetics The kinetics of irreversible inactivation of HRP by H2O2 was studied under pseudo-first order conditions. HRP (20 nM) was incubated with H2O2 (2 mM or 5 mM) at 20 oC in 50 mM buffer at the appropriate pH in a total reaction volume of 1.0 ml. Sodium Acetate (pH 4.0 to 5.5) and Sodium Phosphate (pH 6.0 to 10.0) were used in these experiments. At 3 minute intervals, aliquot of 50 ml were withdrawn and diluted 500-fold, and the residual activities (A) were determined. The activity of an equal amount of the enzyme without pre-incubating with H2O2 was determined and taken as the control (Ao). Plots of ln (A/Ao) against time were used to obtain the pseudo first order inactivation rate constants (kobs). The H2O2 concentrations used in this study were significantly higher than that of the peroxidase, to ensure pseudo-first order inactivation conditions. At least four kobs determinations were made for each H2O2 concentrations. Double reciprocal transformation of the data was carried out using the same software to obtain Kitz-Wilson inactivation constants, kinact and Ki according to the following equation: 1/kobs = Ki/kinact(1/[H2O2]) + 1/kinact
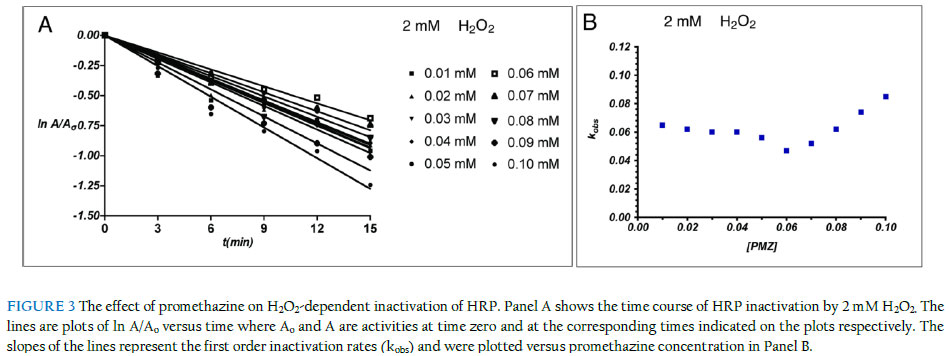
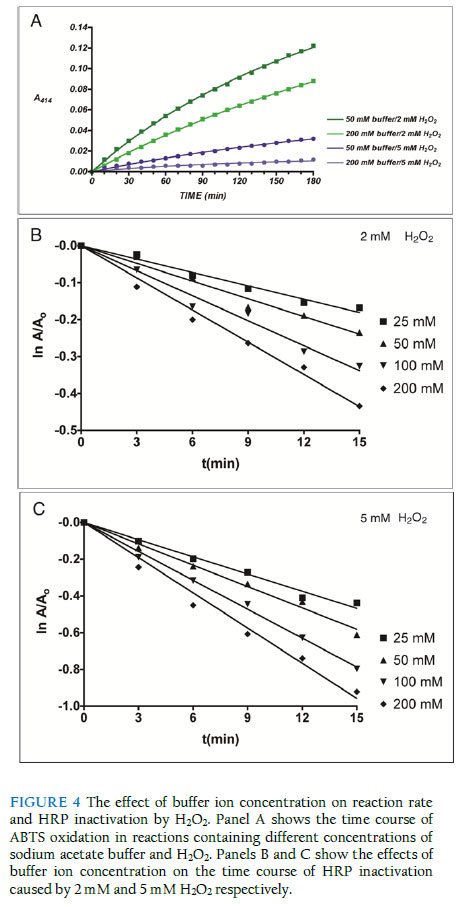
Inactivation kinetics in the presence of promethazine The kinetics of irreversible inactivation of HRP by H2O2 was studied under pseudo-first order conditions. HRP (20 nM) was incubated with promethazine (0.01 to 0.10 mM) at 20 oC in 50 mM sodium acetate buffer (pH 5.4) in the presence of H2O2 (2 mM) in a total reaction volume of 1.0 ml. Incubation of HRP with 2 mM H2O2 without promethazine was carried out under identical conditions. In all cases, H2O2 was added last to the incubation mixture containing HRP and promethazine to initiate the inactivation reaction. At 3-minute intervals, aliquot were withdrawn and analysed as described above. Ionic strength dependent inactivation kinetics The kinetics of irreversible inactivation of horseradish peroxidase (HRP) by hydrogen peroxide was studied under pseudo first order conditions. HRP was incubated with two different concentration of H2O2 (2 mM and 5 mM final concentration) at room temperature at varying buffer concentration (25 mM, 50 mM, 100 mM and 200 mM final concentration). The inactivation process was initiated with the addition of H2O2 to the mixture containing phosphate buffer system at pH 7.0. Aliquot of 3 ml were withdrawn and the residual activities were determined at 3-minute interval and treated as described above. Investigation of effect of ionic strength on the rate of HRP activity The time course of ABTS oxidation by HRP was studied at different buffer concentration. The reaction was initiated by the addition of H2O2 to the mixture containing the buffer (pH 7.0), HRP and ABTS at time zero and the absorbance reading were taking every 10 seconds for 180 seconds at wavelength of 414 nm. Results and discussion pH dependence of irreversible inactivation of HRP by H2O2 We investigated the pH-dependent effect of H2O2 on the inactivation of HRP by monitoring the residual activity of 20 nM of the enzyme incubated with excess H2O2. The first order rate of peroxidase inactivation in the presence of 2 mM (Figure 2A) and 5 mM (Figure 2C) H2O2 were studied as a function of pH. The results show that the inactivation of horseradish peroxidase by H2O2 is influenced by the reaction pH. The observed pattern was different at the two H2O2 concentrations studied. At low concentration of H2O2 (2 mM), the first order rate of inactivation (kobs) was limited and fairly constant between pH 4.0 and 6.5, followed by a steady increase up to pH 10 (Figure 2C). We did not investigate these effects beyond pH 10 since global unfolding of the protein becomes more likely at extreme pH conditions. This pattern suggests that under the conditions of these experiments, the enzyme is more susceptible to suicide inactivation in the alkaline pH range. In contrast to the mild rate of inactivation observed in the presence of 2 mM H2O2 between pH 4 and 6, inactivation rate was markedly high when the reactions were carried out with 5 mM H2O2. As shown in Figure 2D, inactivation by 5 mM H2O2 presented biphasic pH dependence. At 5 mM H2O2 concentration, the inactivation rate was higher in the acidic range compared to the reactions in which 2 mM were used (0.11 min-1 at pH 4) and decreased linearly to 0.02 min-1 (pH 6). The rate of inactivation rose sharply as the pH was gradually changed from the neutral to the acidic region. It is likely that low pH facilitates the generation of inactivating species from the excess H2O2 in the reaction mixture. These findings are generally consistent with earlier studies on the pH-dependence of peroxidase inactivation. Resorcinol-dependent inactivation of lactoperoxidase increases as reaction pH changed from neutral to the acidic range (Divi & Doerge, 1994). Generally, peroxidases are inactivated by extremes of pH, with inactivation more pronounced in the alkaline range than in acidic conditions (Lemos et al., 2000; Mizobutsi et al., 2010). The findings suggest that the balance required between high H2O2 concentrations needed for efficient catalytic conversion and the need to minimise enzyme inactivation can be reached by carrying out reactions at about neutral pH. It will be interesting to know how pH affects the stability of the reaction product and the specificity of the reactions. Modulation of H2O2-dependent inactivation of HRP by redox mediation Redox mediation involves the transfer of electrons from the primary peroxidase oxidation substrate to a secondary substrate via a purely chemical, non-enzymatic reaction (Goodwin et al., 1997). This process also facilitates the conversion of substrates that are poor substrates for peroxidases since they can be oxidised via redox mediation with a primary substrate that is efficiently oxidised by the peroxidase. Redox mediation offers an interesting promise as an alternative route to bioconversion with oxidase enzymes. However, the role of redox mediation in peroxidase inactivation has not been fully studied. This is significant since the highly reactive primary oxidation product has the potential to inactivate the enzyme. The extent and dynamics of suicide inactivation of peroxidases are sensitive to the presence and the nature of the donor substrate (Valderrama et al., 2002; Olorunniji et al., 2009). In order to see if redox mediators could protect peroxidase from destruction by its own substrate, we investigated the effect of promethazine, a known efficient redox mediator (Goodwin et al., 1997; Olorunniji et al., 2000), on horseradish peroxidase inactivation by H2O2 under conditions of catalytic conversion. The first order rates of peroxidase inactivation by H2O2 in the presence of increasing concentration (0 to 0.1 mM) promethazine were studied (Figure 3A). The first order rate of inactivation (kobs) shows a biphasic dependence on promethazine concentration (Figure 3B). There was an initial decrease as promethazine concentration increased from 0 mM to 0.06 mM. This was followed by a steady increase in kobs as promethazine concentration was raised to 1.0 mM. The simplest explanation for this is that lower concentrations of promethazine react with the reactive hydroxyl radical generated by H2O2 in the inactivation process. However, higher concentrations of promethazine lead to the formation of promethazine radicals (Galzigna et al., 1997), which could attack the enzyme and increase the inactivation rate. These results show that promethazine, and indeed any efficient redox mediator, can protect HRP from suicide inactivation if an optimal concentration of the redox mediator is included in the reaction mixture. Yet, these putative ‘protectors’ can potentially be oxidised by the enzyme, and cause inactivation via a donor substrate radical-mediated mechanism (Muraoka & Miura, 2003; Olorunniji et al., 2009). Effect of buffer concentration on catalytic turnover by HRP Results presented above showed that significant changes in pH can sffect the rate of irreversible loss of peroxidase activity. High buffer concentrations are often used to prevent such potential changes in pH during enzyme-catalysed reactions. Hence, we examined if changes in buffer concentration has any effects on inactivation rates. We started by measuring the ABTS/ H2O2 peroxidase activity of HRP in reaction buffers containing 50 mM and 200 mM buffer ions. The time course of ABTS oxidation by HRP shows that the enzyme is more active at lower buffer concentration (Figure 4A). Previous studies have shown that the rate constants for HRP reactions decrease at higher buffer concentration (Alpeeva et al., 2003). We reckoned that the lower activity at 200 mM buffer concentration could be due to inactivation of the enzyme under the reaction conditions that were used. Hence, we studied the first rate of inactivation by 2 mM H2O2 (Figure 4B) and 5 mM H2O2 (Figure 4C) in reaction buffers containing 25, 50, 100, and 200 mM buffer ions. Increasing concentrations of buffer caused increase in inactivation rates of HRP by H2O2. This inhibition was more pronounced when H2O2 concentration was increased from 2 mM to 5 mM. We speculate that the acetate buffer ions used in these experiments react with H2O2 as would a normal substrate to generate reactive inactivating species. The oxidation of carboxylic acids by HRP to generate haem-modifying carboxylic radicals has been reported (Huang et al., 2004). The observed effects of ionic strength on catalytic rate and enzyme inactivation suggest that the desired increase in conversion rate by peroxidases is closely linked with the undesirable inactivation of the enzyme. Changes in buffer ionic strength appear not to alter the conversion/inactivation balance. Further investigations will look at the effects of different buffer ions on the catalytic and inactivation process. Optimizing catalytic conversion in the face of enzyme inactivation The findings here suggest that the balance required between high H2O2 concentrations needed for efficient catalytic conversion and the need to minimize enzyme inactivation can be reached by carrying out the reactions at about neutral pH. The observed effects of buffer concentration on catalytic rate and enzyme inactivation show that the desired increase in conversion rate by peroxidases is closely linked with undesirable inactivation of the enzyme. These factors have to be carefully balanced to achieve optimal effects. While the search for the guiding principles in reaching this balance continues, determination of optimal conditions still needs to be achieved via empirical studies. References
Copyright © 2011 Klobex Academic Publishers The following images related to this document are available:Photo images[bk11017f2.jpg] [bk11017f1.jpg] [bk11017f3.jpg] [bk11017f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}