 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Biokemistri, Vol. 23, No. 3, Dec, 2011, pp. 129-135 Original Article Sphingomyelinase inhibitory and free radical scavenging potential of selected Nigerian medicinal plant extracts Francis M. Awah1,4,5, Peter N. Uzoegwu1, Patrick Ifeonu1,6, Julius O. Oyugi2,3, John Rutherford2, XiaoJian Yao2, Frauke Fehrmann2, Keith R. Fowke2, and Michael O. Eze4,5* 1Tropical
Diseases Research Unit, Department of Biochemistry, University of Nigeria,
Nsukka, Nigeria; Received: 11 September 2011 Code Number: bk11018 ABSTRACT Ceramides from sphingolipid breakdown, and other sphingolipid metabolites, mediate cellular signalling in infectious and other diseases. Therefore, inhibitors of sphingomyelinases (SMases), hold promise as prospective therapeutic agents. Considering the potential therapeutic utility, this in vitro study explored the sphingomyelinase inhibitory, and free radical scavenging potential of five Nigerian medicinal plant leaf extracts, purported to have efficacy against diseases, including HIV/AIDS. The extracts’ sphingomyelinase inhibitory potencies were assessed colorimetrically and theirfree radical scavenging capabilities were assayed by the ability to quench 2,2‐diphenyl‐1‐picrylhydrazyl (DPPH) radical and superoxide anion (O2.‐) radical. Considering their IC50 (μg/ml) values, the extracts inhibited the biochemical activity of sphingomyelinase in a dose-dependent manner, relative to imipramine the standard inhibitor (IC50 38.5 ± 2.4 μg/ml). With Aloe vera as least inhibitory, inhibition increased as follows: Aloe vera (Asphodelaceae) (1132 ± 10.8) <Senna siamea (Fabaceae) (992.2 ± 11.2) < Azadirachta indica (Meliaceae) (984 ± 7.4) < Landolphia owariensis (Apocynaceae) (146.3 ± 9.4) < Stachytarpheta angustifolia (Verbenacae) (100.3 ± 8.7). DPPH radical scavenging relative to ascorbic acid standard increased as: A. indica < A. vera < S. siamea < S. angustifolia < L. owariensis; and superoxide anion quenching, relative to standard rutin increased as: A. vera < S. angustifolia < L. owariensis < S. siamea < A. indica.These results showed thatthe most potent SMase inhibitor was S. angustifolia; whereas, for DPPH radical scavenging and superoxide inhibition, the most potent of the five extracts were L. owariensis and A. indica respectively.These extracts deserve further investigation into their biological effects. KEYWORDS: Sphingomyelinase inhibition; free radical scavenging; medicinal plants INTRODUCTION Sphingomyelinases (SMases) hydrolyze sphingomyelins yielding ceramide and phosphocholine (Barth et al., 2012; Finnegan et al., 2004; Finnegan et al., 2007; Fox et al, 2006; Smith and Schuchman, 2008; Hofmeister et al., 1997). Ceramide, a unique two-tailed amphipathic lipid (two hydrophobic chains on one polar head) has been found to play profound roles in the normal as well as in the pathophysiological states of the individual (Castillo et al., 2007; Grassme et al., 2003) and in the organism under toxicological insult (Ichi et al., 2009; Rebillard et al., 2008). Thus, as has been reviewed (Fox et al., 2006; Ogretmen and Hannum, 2004; Smith and Schchman, 2008) ceramide is a major mediator in inflammation (Philipp et al., 2010), growth and development, differentiation and apoptosis (Jones et al., 1999). It has also shown an effect in neurological disorders such as Alzheimer’s disease and Niemann-Pick disease. In cancer, its pro-apoptotic potency causes tumour-suppressor and antiproliferative effects on various cancer cells (Ogretmen and Hannum, 2004; Reynolds, et al., 2004). There are also reports revealing its effects on atherosclerosis and heart failure (Adamy et al., 2007), endotoxic shock (Józefowski et al., 2010) and ischemia-reperfusion injury (Llacuna et al., 2006). These roles emanate from the ability of ceramide to function as a metabolic second messenger in a plethora of cell signalling events (Barth et al., 2012; Finnegan et al, 2004; Fox et al., 2006). The molecule also does partition specifically and physicochemically into membrane lipid microdomains called “lipid rafts” (Grassme et al., 2003; Ogretmen and Hannum, 2004; Smith and Schuchman, 2008), thereby creating the potential for lateral phase separations in the membrane lipid bilayer. A number of specific and special functions of the membrane are predicated on the presence or transient occurrence of such lateral phase separation events. Such have been implicated in the modulation of HIV infectivity (Finnegan et al., 2004, 2007; Fox at al., 2006) and, in the mouse as well as in mammalian cultured cells, modulation of Pseudomonas aeruginosa infection (Grassme et al., 2003). Involvement of lateral phase separations in membrane-associated processes appears to be universal, and applies in varied ways to various organisms (Eze, 1991). Copious yields of ceramide from the activation of Mg2+-dependent neutral sphigomyelinase (Mg2+-NSMase) have been reported in cultured human neuroblastoma cells and primary cortical neurons. This forms part of the pro-inflammatory effects of the cytokine, tumour necrosis factor alpha (TNF-α) (Barth et al., 2012). This ceramide production is a crucial primary step in the TNF-α – induced superoxide production by NADPH oxidase which leads to damage to neurons via the resulting oxidative stress. Such events also underlie the mechanism of other pro-inflammatory cytokines like interleukin 1 (IL-1) (Grassme et al., 2003; Hofmeister et al., 1997), and IL-6 (Smith and Schuchman, 2008). Ceramides may also participate in the induction of neurodegeneration involved in the aetiology and pathogenesis of cognitive impairment associated with type 2 diabetes mellitus and non-alcoholic steatohepatitis (NASH) (de la Monte et al., 2010). They are implicated as causative agents in the pathogenesis of emphysema in smokers and in animal models as well (Petrache et al., 2005). In addition, ceramide has also been identified as an endogenous pyrogen, a common endogenous metabolite that mediates the rapid phase of the fever response induced by IL-1β or by such exogenous pyrogens as lipopolysaccharides (LPS) of the gram-negative bacteria cell envelope (Sanchez-Alavez et al., 2006). Adamy et al., (2007) reported activation of neutral sphingomyelinase (NSMase) in the rat model of chronic heart failure, and that inhibition of this NSMase, but not acid sphingomyelinase (ASMase), promoted positive outcome in N-acetylcysteine (NAC) therapy of the failing heart. Cells infected with P. aeruginosa produce activated ASMase which protects against the infection, balancing the production of IL-1 with optimum internalization and destruction of the pathogen (Grassme et al., 2003). Enhanced immune activation is synonymous with HIV infection and disease progression (Card et al., 2009; Catalfamo et al., 2008; Koesters et al., 2004; Sousa et al., 2002). In studies of HIV-resistant commercial sex workers, Card and associates (2009) have observed that decreased immune activation is consistent with HIV-resistance. Immune activation entails induction of pro-inflammatory factors such as cytokines (Koesters et al., 2004; Grassme et al., 2003), and oxidative stress (Gil et al., 2003). Enhancement of several indices of oxidative stress has been demonstrated in HIV/AIDS patients (Gil et al., 2003). Ceramide as a pro-inflammatory metabolite (Barth et al., 2012; Grassme et al., 2003) is capable of creating and/or augmenting oxidative stress and inflammatory status in the individual, such as the HIV/AIDS patient (Gil et al., 2003). Thus, antioxidants, as well as anti-inflammatory and other agents that ameliorate these situations are expected to make the patient feel better in health. However, the role of Ceramide and other participating sphingolipid metabolites in HIV infection and AIDS is controversial when viewed in detail. Ceramide inhibits infection in various ways, e.g., facilitating the containment and inactivation of “internalized” HIV [see Fox et al., (2006) for review]. But in the advanced AIDS patient, this lipid contributes to the pathogenesis of HIV-associated dementia (HAD): It is produced via activation of neutral SMase by HIV type 1 gp120, and induces apoptosis and neuropathology through oxidative stress, resulting in HAD (Jana and Pahan 2004). Therefore, there is an emerging rationale that the study of sphingomyelinases may provide some insight into the control of a number of major disease conditions. This notion has, therefore, created the need to explore novel functional inhibitors of the activity of these sphingomyelinases as therapeutic agents, as recently published by Kornhuber et al. (2011) for acid sphingomyelinase. Given the merits of sourcing efficacious, inexpensive, and safe medicines for the world’s masses (WHO 2008), we had earlier proposed strategies for studying and characterizing plants of traditional medicine utility (Eze et al., 1993). In line with this, the present in vitro study focuses on the modulation of sphingomyelinase, and the scavenging of free radicals, including superoxide (agents of oxidative stress), by extracts from selected herbs in use by Nigerian traditional medicine practitioners, and for which claims of efficacy against various infectious and non-communicable diseases have been made (see Table 1). MATERIALS AND METHODS Preparation of leaf extracts The leaves of Aloe vera (Asphodelaceae), Landolphia owariensis (Apocynaceae), Senna siamea (Fabaceae), Stachytarpheta angustifolia (Verbenacae), and Azadirachta indica (Meliaceae) were collected, identified, processed, and extracted with 80% methanol; and the extracts were concentrated and stored at 4oC until use (Awah et al., 2012). Chemicals The solvents ethanol and methanol were purchased from EMD Biosciences (Gibbstown, NJ). L-ascorbic acid, and 2,2-diphenyl-1-picrylhydrazyl (DPPH) free radical, were purchased from Fluka Chemicals. Rutin, ethylenediaminetetraacetic acid (EDTA), phosphate buffered saline (PBS), riboflavin, methionine, and the sphingomyelinase enzymatic assay kit were all purchased from Sigma Chemical Co. (St. Louis, MO). Ascorbic acid, rutin, and imipramine were used as standards for comparing the effects of the various leaf extracts respectively as free radical scavenger, superoxide quencher, or sphingomyelinase inhibitor. Enzymatic assay for sphingomyelinase The sphingomyelinase assay is based on the enzymatic hydrolysis of trinitrophenylaminolauroyl (TNPAL)-sphingomyelin to choline phosphate and trinitrophenylaminolauroyl (TNPAL)-N-acylsphingosine, catalysed by sphingomyelinase from a bacterial source. The assay was performed as per the manufacturer’s protocol (Sigma Chemical Co.). Briefly, the reagents for this assay were prepared fresh and referred to as reagents A – F. Reagent ‘A’ was Tris HCl buffer (500 mM) with magnesium chloride (20 mM) prepared in deionized water, pH 7.4 at 37 oC. Reagent ‘B’ was trinitrophenylaminolauroyl-sphingomyelin substrate solution (TNPAL-sphingomyelin). Immediately before use, 0.045 ml (0.05 mg) of the substrate was pipetted into a one-dram glass vial on ice. The solvent was evaporated from the material with nitrogen gas. Reagent ‘C’ was 0.2 % (v/v) Triton X-100 solution prepared in 10 ml in deionized water. Reagent ‘D’ was isopropanol : heptane : H2SO4 solution (40:10:1) (ISOPRO). Solution ‘E’ was heptane while reagent ‘F’ was the sphingomyelinase enzyme solution prepared immediately before use at a concentration of 1.0 unit/ml in cold Reagent ‘A’. One-dram glass vials were labelled test, blank and control. To each of them, TNPAL-sphingomyelin (45 μl) was pipetted and the solvent evaporated from the material with nitrogen gas until the substrate became a yellow film on the glass vial. Immediately, the vial was returned to ice and capped. To the films, 180 μl of reagent ‘A’ (buffer, pH 7.4) and 200 μl of reagent ‘C’ (Triton X-100) were added and the solutions were mixed by swirling to remove the yellow substrate affixed to the glass and equilibrated at 37oC for 2 min. Then, 10 μl of extract solution (10, 20 or 50 μg/ml) was added to both the test and the blank vial, followed by reagent ‘F’ (enzyme solution) to the test and control vials only. The solutions were swirled and further incubated for 2 min and 1500 μl of reagent ‘D’ (ISOPRO) added to each vial. The vials were capped and placed on ice and uncapped immediately, before adding deionized water (800 μl) and heptanes (1800 μl). The caps were replaced and vials shaken vigorously for 5 min by hand and allowed to sit at 25oC. The upper layer was removed and transferred into suitable quartz cuvettes, and absorbance values read at 330 nm. The sphinogomyelinase inhibitory effect of the extracts was calculated as per the formula:
Where A0 and As represent absorbance of the control and absorbance of the test sample respectively. The decrease in SMase activity measured has been treated here as due to inhibition. It is conceivable that part of the decrease could have been due to inactivation which was not assessed in this work. However, this does not alter the conclusions proffered. Free radical inhibition assays Quantitative DPPH radical-scavenging assay Scavenging activity on DPPH free radicals by the extract was assessed according to the method reported by Gyamfi et al. (1999) with slight modifications (Awah et al 2010). In this present work, % inhibition, of DPPH radical-scavenging activity was calculated according to the equation:
Where, as in Awah et al. (2010), A0 is the absorbance of the control, and As is the absorbance of the tested sample. The IC50 value represented the concentration of the extract that caused 50 % inhibition of DPPH radical and was calculated by linear regression of plots, where the abscissa represented the concentration of tested sample and the ordinate the average percent of inhibitory activity from three replicates. TABLE 1 Ethnobotanical data of selected Nigerian medicinal plants investigated
Superoxide radical (O2.-)-scavenging assay This assay was based on the capacity of the extract to inhibit the photochemical reduction of nitro blue tetrazolium (NBT) (Beauchamp and Fridovich, 1971) and the method of Martinez et al., (2001) to determine superoxide dismutase as modified by Awah et al. (2010). The percentage inhibition of superoxide generation was also estimated by comparing the absorbance of the control and that of the reaction mixture containing test sample as per the equation:
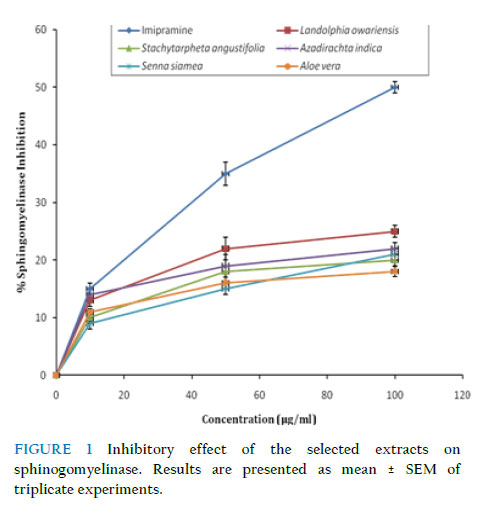
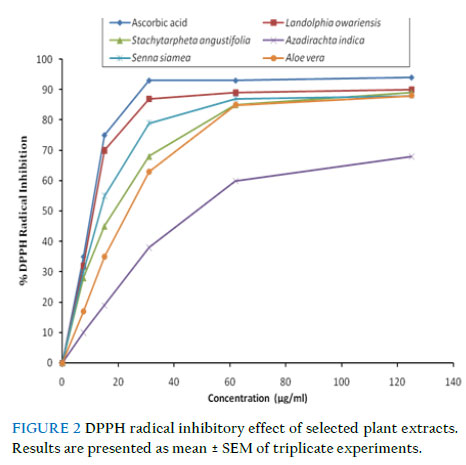
Where A0 is the absorbance of the control, and As is the absorbance of the test sample (Awah et al., 2010). Statistical analysis Data were analysed using one-way analysis of variance (ANOVA) at 0.05 level of significance using the statistical package for social sciences (SPSS) version 17.0 for windows software package. Linear regression plots were done using Microsoft Excel for Windows Vista. All the results are expressed as mean ± standard error of the mean (SEM) (n = 3). Results Sphingomyelinase inhibitory potency of extracts As shown in Fig. 1 and Table 2, the selected plant extracts inhibited sphingomyelinase in a dose-dependent manner compared to imipramine, a standard sphingomyelinase inhibitor. S. angustifolia showed the highest inhibitory potency (IC50 = 100.3 ± 8.7μg/ml) with maximal inhibition of 26 % compared to 49 % for imipramine (IC50 = 38.5 ± 2.4 μg/ml) at a concentration of 100 μg/ml. A. vera had the least inhibitory potency of 18 % at the concentration of 100 μg/ml. Free radical inhibition assays Effect of extracts on DPPH radical All extracts showed significant dose-dependent DPPH radical scavenging capacity (Fig. 2). Among them, L. owariensis was most efficient, inhibiting 92.3 ± 4.5 % of DPPH at a concentration of 125 μg/ml compared to ascorbic acid which inhibited 94.2 ± 3.2 % at the same concentration. As shown by their IC50 values (Table 3), the DPPH radical scavenging efficiency of the extracts increased as follows: A. indica < A. vera < S. siamea < S. angustifolia < L. owariensis. The radical scavenging ability of the plant extracts could be as a result of the presence of secondary metabolites such as phenolic compounds (Awah et al., 2010). TABLE 2 IC50 for sphingomyelinase inhibitory potency of selected extracts
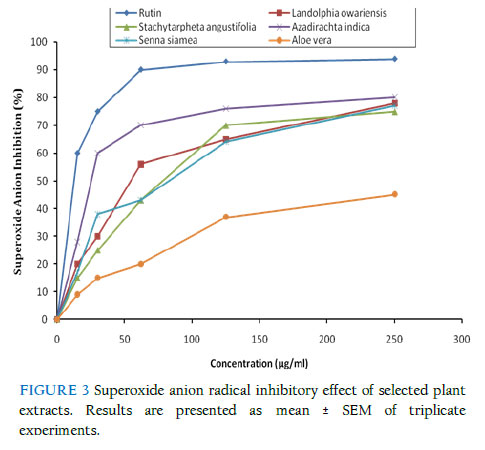
Effect of extracts on superoxide (O2.-) anion radical All the plant extracts inhibited the formation of reduced NBT in a dose-related manner. As shown in Fig. 3, A. indica showed the maximal O2.- anion inhibitory activity of 80.7 ± 1.4 % at the concentration of 250 μg/ml, followed by L. owariensis (78.6 ± 2.8 %, at 250 μg/ml), while A. vera showed the lowest inhibitory activity (45.8 ± 5.2 %, at 250 μg/ml) compared to rutin (94.0 ± 2.2 %, at 250 μg/ml). The O2.- scavenging effect of the extracts could culminate in the prevention of .OH radical formation since O2.- and H2O2 are required for .OHradical generation. Superoxide anion (O2.–) forms from the inadvertent single electron reduction of molecular oxygen in the mitochondrial (or other related) oxidative electron transport chain (Grivennikova & Vinogradov, 2006). It is also produced in large amounts as a cellular immune response against various types of infection, and in other disease conditions (Eze et al., 1993 for review). Superoxide then becomes the precursor of the other reactive oxygen species: hydrogen peroxide (H2O2), hydroxyl free radical (.OH), hypohalite (e.g., OCl-), and others. DISCUSSION The potencies of the leaf extracts of the following five Nigerian medicinal plants for quenching free radicals (DPPH and superoxide anion radicals), and for inhibiting the enzyme, sphingomyelinase (SMase), have been studied: A. vera (Asphodelaceae), L. owariensis (Apocynaceae), S. siamea (Fabaceae), S. angustifolia (Verbenacae), and A. indica (Meliaceae). In a dose-dependent manner, the five leaf extracts each inhibited SMase, and scavenged free radicals, including superoxide radical, to varying degrees. The most potent SMase inhibitor was S. angustifolia; whereas, for DPPH radical scavenging and superoxide inhibition, the most potent of the five extracts were L. owariensis and A. indica respectively. Scavenging of superoxide prevents its fast (diffusion-controlled) combination with nitric oxide [also produced under pro-inflammatory conditions (Codoner-Franch et al., 2011)] to form peroxynitrite ONOO-, a rather devastating cellular oxidant; thus, saving the cells/tissues from oxidative damage (Castillo et al., 2007; Sandoval et al., 1997). Ceramide, one of the products of SMase activty, is an endogenous pyrogen (Sanchez-Alavez, et al., 2006). It is also a major factor in the cellular signalling pathways of inflammation, oxidative stress (Ichi et al., 2009), and apoptosis (Jones et al., 1999), as well as in all the metabolic and pathophysiological pathways modulated by these states and events in the individual (Barth et al., 2012; Fox et al., 2006). These include pathways regulated by such pro-inflammatory cytokines as TNF-α (Barth et al., 2012), interleukin 1 (IL-1) (Grassme et al., 2003; Hofmeister et al., 1997), and IL-6 (Smith and Schuchman, 2008). Only further testing will determine if the extracts’ radical scavenging and superoxide quenching potencies would be protective against radical/oxidative stress – induced injury. The claims of the herbalists to the effect that any of these herbs can be employed for successful treatment of the various ailments may have a theoretical basis but as of yet remain untested scientifically. The present findings reported herein, therefore, constitute a point at which to begin to design the appropriate experiments for unravelling further details. TABLE 3 Free radical and superoxide anion scavenging potency (IC50)
Acknowledgements This work was supported by the following grants which are hereby gratefully acknowledged: a grant from the Department of Foreign Affairs and International Trade, Canada, to MOE and KRF to support FMA; a Canadian Institute for Health Research (CIHR) grant # MOP 86721 to KRF; a University of Nigeria Senate Research Grant to PNU; a Special Grant from the University of Winnipeg (UofW) Research and Graduate Studies Office to MOE and Dr. Desiree Vanderwel (Chemistry Dept., UofW); and a UofW Major Research Grant (#7437) to MOE. The authors are grateful to the Global College of UofW for supporting Francis Awah’s sojourn in Winnipeg; as well as to Dr. Apollinaire Tsopmo (Dept. of Chemistry, Carleton University, Ottawa, Canada), and Dr. Chibuike Udenigwe (Dept. of Human Nutritional Sciences, University of Manitoba) for useful suggestions. References
Copyright © 2011 Klobex Academic Publishers The following images related to this document are available:Photo images[bk11018f2.jpg] [bk11018f3.jpg] [bk11018f1.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

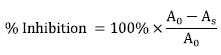
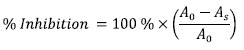
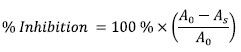
{kind=link}
{kind=link}
{kind=link}