 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Journal of Culture Collections, Volume 3, 2000-2002, pp. 3-14 CHARACTERISTICS OF SOIL ACTINOMYCETES FROM ANTARCTICA Penka Moncheva1*, Sava Tishkov1, Nadezhda Dimitrova1, Valentina Chipeva2, Stefka Antonova-Nikolova1 and Nevena Bogatzevska3 1Sofia University, Biological Faculty, Department of General
and Industrial Microbiology, 8 Dragan Tzankov st., 1164 Sofia, Bulgaria; Code Number: cc02001 Summary Forty-seven actinomyces strains were isolated from Antarctic soils – nineteen of them showed antagonistic activity against Gram-positive and Gram-negative bacteria. Six of the strains possessed a broad spectrum of antibacterial activity. Results obtained from the physiological and biochemical analyses including determination of 39 characteristics proved that two of the strains (23 and 29) were similar whereas all the rest differed among each other. Morphological studies indicated that the strains belonged to the genera Streptomyces, Actinomadura and Kitasatosporia. Antibacterial activity of three actinomycetes strains (designed as 29, 30 and 47) was confirmed in batch culture. They were active against clinical isolates from the species Staphylococcus aureus and Streptococcus pneumoniae. The three strains also showed antibacterial activity against the phytopathogenic bacteria Xanthomonas axonopodis pv. glycines, X. vesicatoria, X. axonopodis pv. phaseoli, Pseudomonas syringae pv. tomato and Clavibacter michiganensis, for which no biological means for control, had been developed yet. The broadest spectrum of antibacterial action had the strain 29. The antibacterial compounds produced by these strains probably possessed non-polar structure and consisted of several active components. Introduction Actinomycetes are prokaryotes with extremely various metabolic possibilities. They produce numerous substances essential for health such as antibiotics [4, 10, 22, 23, 25], enzymes [3, 8, 14, 15, 16, 17], immunomodulators [13], etc. In the last decades actinomycetes became the most fruitful source for antibiotics. In the 60s’ and 70s’ of the 20th century 75 to 80 % of all discovered antibiotics derived from the order Actinomycetales, mainly from Streptomyces species. In the 70s’ and 80s’ the ratio and significance of the other, non streptomycete actinomycetes (so called rare actinomycetes) increased up .to 20 % of all microbial antibiotics, and 30-35 % of Actinomycetales species. Considering the practically useful compounds, today about 130 to 140 microbial products and a similar number of derivatives (including semi-synthetic antibiotics) are applied in human medicine, mostly in chemotherapy, and veterinary medicine. Furthermore, some of 15 to 20 compounds are used in agriculture mainly as pesticides, plant protecting agents and food additives. The majority of these compounds, except fungal penicillins, cephalosporins and several bacterial peptides and few others, are also produced by actinomycetes. The high percentage of new compounds derived from new target oriented screening methods is also of actinomycetal origin. In the late 60s’, after the discovery of gentamycin, originated from Micromonospora, the study of non-streptomycete actinomycetes received increasing attention. In general, it was observed that most of rare actinomycetes products had already existed among streptomycetes metabolites. Nevertheless, there were certain types of structure, which occurred more frequently in some rare actinomycetes species than in streptomycetes. Amycolatopsis (formerly Nocardia) and Actinomadura species frequently produced vancomycin-type glycopeptides. The promising antitumor enediyne antibiotics were produced exclusively by rare actinomycetes. The members of new groups of macrolactam and naphtacene-quinone antibiotics were isolated from Actinomadura. The species from this genus frequently produced polyether antibiotics. Micromonospora and Saccharopolyspora strains were relatively rich sources of macrolides. The productivity of Streptomyces strains as antibiotic producers remains unique amongst Actinomycetales strains. The relatively low occurrence of non-streptomycetes species as producers of secondary metabolites is due to difficult techniques required for the isolation of these strains from the environment. Other problem is their complicated preservation and cultivation methods, which frequently require some specific and unusual conditions. These are the main reasons for regarding these microbes as rare organisms and the difficulties for investigations and manufacturing of their products [4]. Some new screening programs have been already developed for discovering of new species or unknown bioactive sub-stances. One of the modern approaches is isolation and screening of microorganisms from relatively unknown or unstudied areas. In this meaning Antarctica is of significant interest. Prokaryotes dominate in many Antarctic ecosystems and play important role in consumption chains and biogeochemical cycles [1, 9, 19]. Among microorganisms isolated from such areas actinomycetes are the least frequently met. In the recent years using molecular genetics methods it was found that the majority of the isolated microorganisms are new species [20]. This expands the possibilities for their use in biotechnology. Aim of the present study was isolation of actinomycetes - producers of antibacterial substances from Antarctic soils. Materials and Methods Microorganisms. Actinomycetes strains isolated from three soil samples, taken in February 2000 from different regions of Livingston Island, Antarctica were the objects of this study. The test microorganisms listed in Table 1 were used to screen antibacterial activity of actinomycetes strains. The phytopathogenic test-bacteria were naturally isolates from ill plants and belonged to the collection of Plant Protection Institute – Kostinbrod. Media and cultivation conditions. For the isolation, cultivation and maintaining of actinomycetes strains selective media - AV agar and starch-casein-nitrate agar [2] were used. Test-bacteria were cultivated in Nutrient agar (Difco), Blood agar (Gibco) and Potato agar (Oxoid). The biosynthesis of antibacterial substances was tested in conditions of dynamic batch cultivation. The active strains were cultivated on a shaker at 25°C and 240 rpm for 120 h. The fermentation media were inoculated with 10 % of inoculum taken from 48th h culture. Both fermentation and inoculation media had the same composition. Isolation of the cultures. The Actinomycetes strains were isolated using Pasteur method. Characterization of Actinomycetes cultures. Morphological, cultural, physiological and biochemical properties of the strains were studied according to methods of Shirling and Gotlieb [24] and Bergey [5]. Table 1. Test-bacteria used for screening of antibacterial activity.
Antibacterial activity. Antibacterial activity of the strains was measured by diffusion method using agar blocks or ethanol mycelium extracts and culture medium filtrates. Analysis of antibacterial substances. Ethanol extracts were used for primary analysis of the antibacterial substances. It was performed by:
Results and discussion 1. Screening of isolated actinomycetes for antimicrobial activity Forty-seven actinomycetes strains were isolated in accordance with differences in the colony morphology. The strains were purified by visual, microscopic and cultivation methods and were maintained on the same media, which was used for isolation, at 4°C. After primary screening for antibacterial activity 19 strains were selected, by which antibiotic activity varied from 14 to 37 mm sterile zone against to referent strain B. subtilis ATCC 6633. The spectrum of antibiotic activity of selected strains was determined using the full set of test bacteria. It was found that 18 strains suppressed in different degree the test microorganisms (Table 2). Most of them were not active against E. coli but repressed the growth of St. aureus, E. faecalis and L. innocua in a significant extent. Eleven strains possessed activity against gram-negative phytopathogenic bacteria but they also expressed antibacterial activity to Gram-positive bacteria (Table 3). It could be suggested that the produced antibiotic substances had a broad spectrum of activity. There was a little information in the literature about antibiotics active against phytopathogenic bacteria [6, 11, 12, 18]. For this reason our interest was focused on the strains, which proved to possess such an activity - numbers 14, 23, 29, 30, 34 and 47. They were cultivated in liquid media on shaker for 120 h. The antibacterial activity of ethanol extracts of the mycelium and culture media filtrates was determined by well diffusion method on agar plates. In these conditions only three strains (29, 30 and 47) confirmed their antibacterial activity. They inhibited the growth of nine from the tested 14 bacterial strains. The antibiotic substances that repressed B. subtilis and Ps. syringae pv. tomato were accumulated both in the mycelium and in the media. The substances repressing St. aureus, E. faecalis were accumulated only in the mycelium, and the ones against the other phytopathogenic bacteria only in the media (Table 4). Most probably, the medium was not suitable for antibiotic biosynthesis from the rest three strains. Optimization of the conditions for antibiotic synthesis of the strains 29, 30 and 47 as well as establishing a suitable media for antibiotic production from stains 14, 23 and 34 will be the aim of further studies. Table 2. Antibacterial activity of selected
actinomycetes strains.
a Test-bacteria: X. axonopodis pv. glycines (1), X. vesicatoria PT (2), X. axonopodis pv. phaseoli (3), X. vesicatoria T (4), Ps. syringae pv. tomato R0 (5), E. amylovora (6, 7), E. chryzanthemii subsp. dianthicola (8), Cl. michiganensis subsp. michiganensis (9). Table 4. Antibacterial activitya of three actinomycetes strains after cultivation in liquid medium.
a Activity
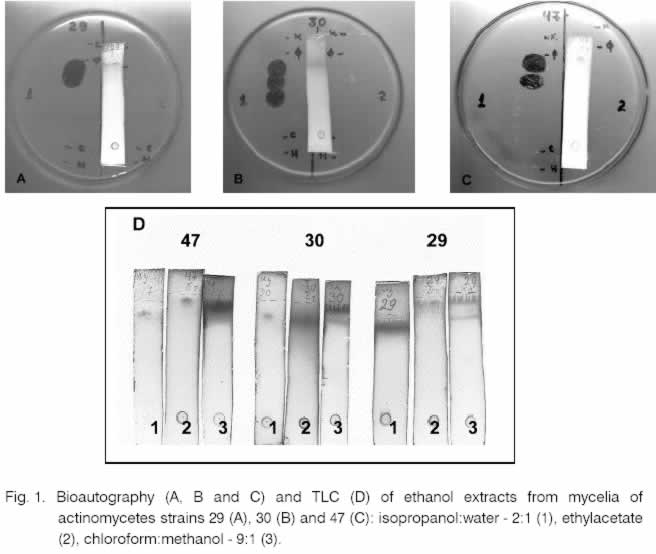
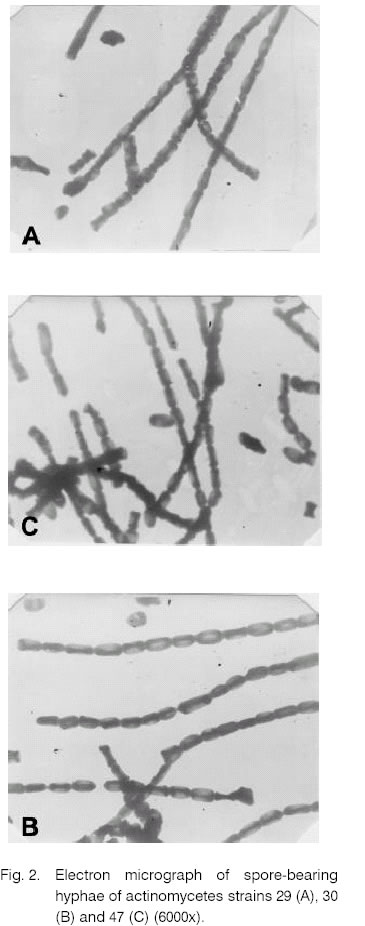
is represented as mm of inhibition zone. The antibacterial activity of the strains 29, 30 and 47 filtrates and extracts was also tested against clinical isolates of the strains St. aureus and Str. pneumoniae (Table 4). The obtained results were interesting because of the frequent occurrence of resistant forms among pathogenic bacteria. 2. Primary studies of the antimicrobial substances produced by strains 29, 30 and 47 An experiment for analyzing the component composition of the antibacterial substances produced by these three strains was performed. The ethanol extracts of the mycelia taken after cultivation for 6 days were analyzed by TLC. The plates were developed in three different systems for separation (ethylacetate, chloroform-methanol and isopropanol-water). 2.1. Bioautography of the ethanol extracts The bioautography of the chromatographic plates showed that the best separation was reached in the system containing ethylacetate (Fig. 1). The bioautography after separation with chloroform-methanol showed that all strains formed single sterile zones. When ethylacetate system was used only strain 29 formed single sterile zones close by the front. The strain 47 and 30 formed two and three zones respectively. This indication showed that the antibacterial activity of these strains was due to antibiotic substances containing two or more bioactive components. Furthermore it is possible that the antibiotic activity of the filtrates of strains 29 and 47 towards phytopathogenic tests may be due to additional component secreted out of the cell. After dyeing of the chromatograms by vanilin-sulphur acid reactive for all three strains single spots were stained which correspond to the closest to the front sterile zones in bioautography. Undeveloped components (by this reactive) probably had more polar character, e.g. their bioactive components had different chemical nature. 3. Characteristics of the strains with antimicrobial activity A great variety of morphological, cultural, physiological, biochemical and other features of the six strains (14, 23, 29, 30, 34 and 47) were studied. 3.1. Morphological features Morphology of the actinomycetes colonies was determined in the selective media, from which they had been isolated. The micro-morphological characteristics were studied by light and electronic microscopy on the 7th, 14th and 21st day cultures in 4 different media (ISP 2, 3, 4, 5). The spore-bearing hyphae of the strains were straight to flexible. The strain 29 formed spore-bearing hyphae of type “open loops, hooks or extended spirals” (Table 5 and Fig. 2). The number of the spores was higher than 20, which indicated they refer to the long chains of spores. Long chains of conidia were formed by strains belonging to various genera. These might include Actinomadura, Streptomyces, Kitasatosporia, Actinopolyspora, Saccharopolyspora, etc [5]. Fragmentation of the mycelium was not observed during the growth of the strains in liquid media. Many actinomycetes strains (Streptomyces, Actinomadura, Kitasatosporia, Termoactinomyces) formed stable filaments [5]. The spores of all strains had smooth surface (Fig. 2). The antibacterial activity of the strains 29, 30 and 47 filtrates and extracts was also tested against clinical isolates of the strains S. aureus and Str. pneumoniae (Table 4). The obtained results were interesting because of the frequent occurrence of resistant forms among pathogenic bacteria. The aerial and substrate mycelium color was determined on media ISP 2, 3, 4, and 5, using Bondartzev color scale [7]. The results are shown in Table 6. According to Nonomura criteria [21] only the substrate mycelium of the strain 14 formed defined pigment on media ISP 3 and ISP 5 i. e. the result was positive. The rest of the strains did not produce distinctive pigment on the substrate mycelium. Table 5. Morphologya of spore-bearing hyphae.
a Morphology: straight (rectus, R), flexible (F), open loops, hooks or extended spirals (retinaculum – apertum, RA). Table 6. Color of aerial mycelium of actinomycetes strains.
3.2. Physiological and biochemical features The results of this study are presented in Table 7. The strains showed similarity in their ability to assimilate 15 carbon sources. Some differences occurred in assimilation of fructose, xylose and saccharose. None of the strains grew on substrates like adonitol, xylitol and isobutanol. Table 7. Biochemical and physiological characteristics of actinomycetes strains.
Degradation of 9 substrates by the strains was tested. The strains did not hydrolyze casein and gelatin. They differed in ability to decompose arbutin, esculin, starch and urea. Acid formation from four sugars (glucose, ramnose, saccharose and xylose) was studied. The strains differed among them-selves in ability to produce acid. Some similarity was observed only concerning the saccharose, which was metabolized from all the strains without acid formation. The strains 29, 30 and 47 reduced nitrates whereas 14, 23 and 34 did not. All strains produced melanoid pigment on both peptone-yeast extract iron agar (ISP 6) and tyrosine agar (ISP 7). The ability of Actinomycetes to grow at three different temperatures (25°C, 37°C, and 50°C) and 6 different concentrations of NaCl in the media was thoroughly studied. All strains grew at 25°C and one of them (strain 30) even with lower degree grew at 37°C too. All strains grew at 1 % NaCl. Most of them were tolerant to 3 % NaCl in the media. Only three strains (23, 29 and 30) grew at 5 % NaCl. There was no growth detected at higher concentrations of NaCl. Genera Actinopolyspora and Saccharopolyspora, to which the strains studied were morphologically close, had NaCl tolerance (10 - 20 % of NaCl) [5] On the base of their physiological and biochemical characteristics the strains were subjected on hierarchical cluster analysis. The clustering is presented by dendrogram in Fig.3. At 40 % similarity three clusters were formed. The first included three strains (23, 29 and 30), the second - two strains (14 and 34) and the third included only one strain (strain 47). At similarity 80 % five clusters were formed - one cluster included two strains (23 and 29 respectively). The strains 14, 30, 34 and 47 formed single clusters. The affiliation of strain 47 to a single cluster in both analyses is a proof for its species differentiation. The same could be true for strains 14, 30 and 34 while 23 and 29 probably are strains of one species. On the basis of determined characteristics of the strains it could be concluded they are close to the genera Streptomyces, Actinomadura, Kitasatosporia. Determination of additional characteristics of the strains and further detailed analysis of the antibacterial substances synthesized by them probably will reveal their taxonomic position as well as the nature of the substances. Acknowledgements. This study was supported by National Science Fund of Bulgarian Ministry of Education and Science (grant CC-1102/2001). References
Copyright 2002 - National Bank for Industrial Microorganisms and Cell Cultures - Bulgaria The following images related to this document are available:Photo images[cc02001f1.jpg] [cc02001f3.jpg] [cc02001f2.jpg]
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

{kind=link}
{kind=link}
{kind=link}