
|
Journal of Culture Collections
National Bank for Industrial Microorganisms and Cell Cultures
ISSN: 1310-8360
Vol. 3, Num. 1, 2002, pp. 84-91
|
Journal of Culture Collections, Volume 3, 2000-2002,
pp. 84-91SEROLOGICAL AND ELECTROPHORETIC CHARACTERIZATOIN
OF THE NECROTIC STRAIN CMV-NB OF CUCUMBER MOSAIC VIRUS Angela Yordanova1*, Dimitrinka Hristova2 and
Elisaveta Stoimenova3 1National Bank for Industrial Microorganisms
and Cell Cultures, 1113 Sofia, P. O. Box 239, Bulgaria;
2Plant Protection Institute, 2230 Kostinbrod, Bulgaria;
3Institute of Genetics, Bulgarian Academy of Sciences, 1113 Sofia, Bulgaria Code Number: cc02013
SummarySerological and electrophoretic properties of Bulgarian
isolate CMV-NB causing tomato fruit necrosis were investigated. Its serogroup
affiliation was studied by means of indirect ELISA with polyclonal and
monoclonal antibodies as well as by immunodiffusion and immunoelectrophoresis
with sera against subgroups I and II of CMV. Strain NB belongs
to subgroup I but distinguishes from the other tested strains from this
group in counter and rocket immunoelectrophoresis especially. The electrophoretic
mobility of capsid protein and whole virions of NB was also compared with
those of other virus strains. Strain NB was
characterized with very bigger mobility of virus particles in electric
field. It explained differences established in immunoelectrophoretic analysis.
The obtained results supposed presence of mutations in CMV-NB, which leaded
to increasing negative surface charges of virions. Introduction Cucumber mosaic virus (CMV) is a type representative
of cucumovirus group. Its icosahedronic 30 nm diameter virions contain one
kind of capsid protein with molecular weight approximately 24 kDa. CMV possesses
three-component genome and a subgenome RNA [5]. Some strains also contain
another, fifth satellite RNA. It could significantly affect the induced symptoms
and could cause, for example, tomato necrotic disease [12, 13, 29]. CMV is
spread worldwide and it is presented by plenty of strains, which are divided
into two main serogroups or subgroups. The groups are denoted in a different
way depending on the methods used: DTL and ToRS, U and N, I and II, WT and
S etc. [3, 21, 22, 23, 24]. CMV is of great importance for the Bulgarian
agriculture, having big influence on tomato yield especially. A series of
research on the strain variety have been performed using indicator test,
different serological methods and PCR [4, 9, 11, 15, 19, 27, 33]. From tomato
plants cv. Carmelo, which have shown stem and fruit necrosis, was isolated
a strain denoted as CMV-NB [29]. This strain contains a satellite RNA, which
results in attenuation to a different degree of the symptoms in some plant
species and also in well-exhibited necrosis on tomato stems and fruits [28]. The aim of the present research was a further
characterization of the necrotic strain CMV-NB by means of serological and
electrophoretic methods. Materials and Methods Viruses. The test object was the CMV-NB
strain (NBIMCC 2126). The viruses used for control were: 1) Bulgarian CMV
strains isolated from cucumber (Uo, 1k-92), tomato (MB, 146D), pepper (PB,
108P) and tobacco (TB, 131T) [28]. 146D and 108P strains belong to subgroup
II and the rest ones to subgroup I [11, 19]; 2) American strains isolated
from bean (7, subgroup Ia), pepper (26, subgroup Ib) and Vinca major (3,
subgroup II) [2]; muskmelon (Fny, subgroup I) [25]; Campanula rapunculoides (GR,
subgroup II). All the above-mentioned strains belong to the NBIMCC collection. Virus multiplication. The CMV strains
were propagated and maintained in Nicotiana tabacum cv. Nevrocop 1146
and tobacco cv. HT (containing N gene from N. glutinosa). Leaves with
well exhibited systemic or local symptoms were used for further investigation. Virus purification. The applied method
was the one of Lot et al. [18] with some modifications. After double differential
centrifugation the viruses were suspended in 0.01 M K-Na phosphate buffer,
pH 7.2, containing 5 mM EDTA. The concentration was determined spectrophotometrically. Antisera (AS). Four Bulgarian sera produced
by the team were used as follows: homologous to subgroup I CMV strains
- Uo [34], MB and PB and homologous to subgroup II - 146D [10]. Their titres
in agar diffusion test were 1:8192, 1:512, 1:1024 and 1:4096 respectively.
A German antiserum for serogroup N was applied too. ELISA (enzyme linked immunosorbent
assay). ACP-ELISA (antigen coated plate) according to Koening [14] was
performed. 1. With polyclonal antibodies (PAbs) Uo and
146D. The antigen (plant sap) was dropped in dilution of 1:25 in
carbonate buffer, pH 9.6, and incubated for 16 hours at 4°C. The IgG
were applied diluted 1:500 in PBS-T with 2 % PVP and 1 % OA for 2 hours at
35°C. Antirabbit peroxidase conjugate 1:1000 in PBS-T with 1 % OA was
added for 2 hours at 35°C. The used substrate was orthophenylenediamine
and the extinction was read at 492 nm. 2. With monoclonal antibodies (MAbs). MAbs
for subgroup I (DTL), subgroup II (BD9) and not serotype-specific (CF11)
were used [6, 7]. The scheme for ACP-ELISA was the same with one exception
- antimouse peroxidase conjugate was applied. Double radial diffusion according to Ouchterlony
and Nilsson [20]. It was performed in 0.7 % agarose gel in 0.075 M veronal
buffer, pH 8.6, containing 5 mM EDTA. The viruses were applied in the form
of purified preparations (15 mg in a well) while the antisera - in dilution
of 1:2 - 1:4. The gels were stained with Blue R-250 [32].
Immunoelectrophoresis. It was carried
out in 1 % agarose gel in 0.075 M veronal buffer, pH 8.6, containing 5 mM
EDTA, at 150 V and 10°C for 2 – 2.5 hours. The gel staining was the
same as in the immunodiffusion. 1. Counter immunoelectrophoresis. The
antisera were applied in dilution of 1:2 - 1:16 and the virus preparations
were 5 mg each. 2. Rocket immunoelectrophoresis [32]. The gel contained
30 ml Uo antiserum or 35 ml 146D antiserum respectively. The antigens
were also applied as purified preparations.
Electrophoresis of virus particles. It was carried
out under the same conditions as the rocket immunoelectrophoresis without
the application of the antisera. Electrophoresis of virus protein. SDS-polyacrylamide
gel electrophoresis (SDS-PAGE) according to Laemmli was performed [16]. The
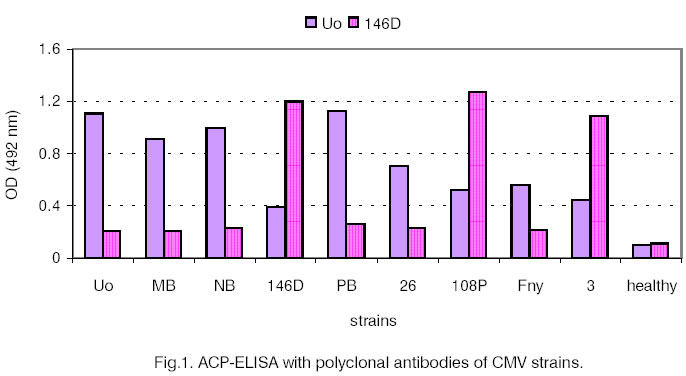
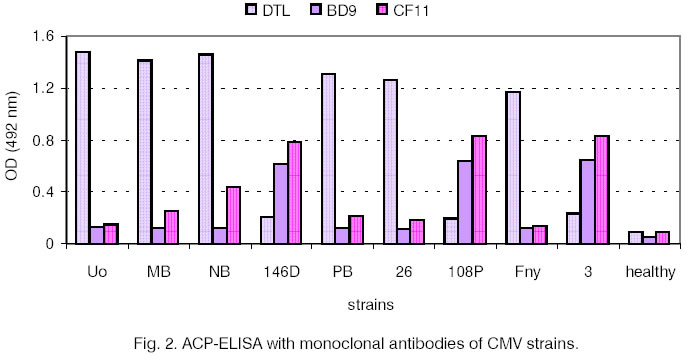
gels were stained with Blue R-250. Results and Discussion For initial serological differentiation of
the virus isolates ACP-ELISA with PAbs and MAbs was performed (Fig.
1 and Fig. 2). The reactions of 8 other strains of CMV are
shown for comparison. When PAbs were applied CMV-NB behaved as a strain from
subgroup I. The extinction values with IgG Uo were more than four times as
high as the ones with IgG 146D. The results obtained after using MAb DTL were
similar to those with IgG Uo. With MAb BD 9 the strains from subgroup I showed
lower values and the ones from subgroup II (146D, 108P, 3 and GR) - high
values. CMV-NB did not differ from the other tested subgroup I strains. The
application of MAb CF11, however, revealed higher extinction values for NB
(0.443) compared to the other strains from subgroup I (Fig.
2) but CF11 was not serotype-specific. Both MoAbs (BD9
and CF11) have been prepared for different CMV epitopes and have been used
in combination for detection of serotype N [6, 7]. The serological relationship between NB and
other CMV strains, belonging to subgroups I and II [11], was also investigated.
Antisera against MB, PB, Uo (subgroup I), 146D and N (subgroup II) were used.
The data obtained from the double radial immunodiffusion, and also from the
counter immunoelectrophoresis (for some of the sera) are summarised in Table
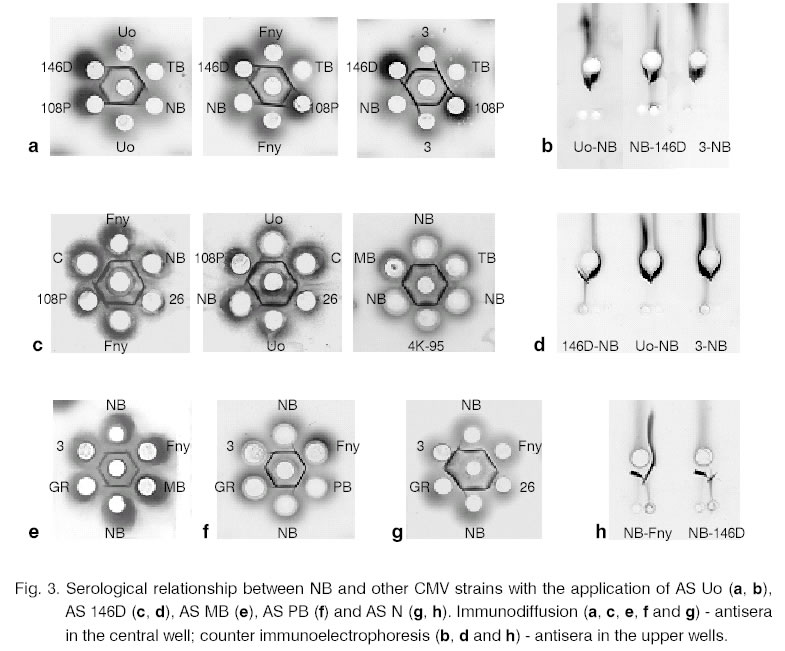
1. Part of the serological reactions is illustrated in Fig.
3.
Table 1. Serological relationship between
CMV-NB and other CMV strains.
|
CMV strains
subgroup
|
CMV-NB
|
|
AS MB
|
AS PB
|
AS Uo
|
AS 146D
|
AS N
|
|
Uo
|
I
|
+a
|
+
|
+b/+c
|
+/(+)
|
-/+
|
|
4K-95
|
I
|
+
|
|
|
+
|
-/+
|
|
MB
|
I
|
+
|
|
+
|
+
|
-/+
|
|
146D
|
II
|
++
|
++
|
++, x/+
|
+/+
|
+/+
|
|
PB
|
I
|
|
+
|
|
+
|
-/+
|
|
108P
|
II
|
++
|
++
|
|
+/+
|
+
|
|
TB
|
I
|
|
|
+/+
|
+
|
-
|
|
7, 26
|
I
|
|
+
|
+
|
+
|
-/+
|
|
3
|
II
|
++
|
++
|
++, x/+
|
+/+
|
+/+
|
|
Fny
|
I
|
+
|
+
|
+
|
+
|
-/+
|
|
GR
|
II
|
++
|
++
|
++,x/+
|
+
|
+
|
aspur formation from the control strains (+), spur formation from CMV-NB (++),
fusion of the precipitation lines (-), crossing of the precipitation lines
(x);
bimmunodiffusion;
ccounter immunoelectrophoresis. While tested with AS MB and AS PB the precipitation
lines of NB formed spurs with ones of the strains 146D, 108P, 3 and GR from
subgroup II. On the other hand, the tested strains from subgroup I produced
spurs when NB was placed in adjacent well. The immunodiffusion with AS Uo
also resulted in differentiation between NB and the other strains from subgroup
I. CMV-NB demonstrated a well-expressed spur to subgroup II strains
and, in some cases, crossing of the precipitation lines were observed. The
immunoelectrophoretic analysis with AS Uo revealed different patterns. All
the tested strains formed spurs towards NB. The immunodiffusion with AS N determined NB
as a strain from subgroup I. The precipitation lines fused when in the neighbouring
wells Uo, 4K-95, MB, PB, TB, 7 and Fny were applied. The spurs with subgroup
II strains were observed. In counter immunoelectrophoresis, however, all
the tested strains produced spurs towards NB (Fig.
3). The testing of NB with AS 146D revealed that
the precipitation lines of 146D, 108P, 3 and GR formed well-exhibited spurs
when both methods were carried out. A smaller spur was made from the subgroup
I strains and it was better expressed in the double immunodiffusion. The testing of the CMV control strains of one
to another gave uniform results (Fig. 3). After
placing AS I the precipitation lines of subgroup I strains formed spurs while
after applying AS II the reaction was the opposite (the data are described
in details in separate publications [11, 14]). The obtained results show that CMV-NB belongs
to subgroup I but distinguishes from all the other tested strains from this
group. Depending on the titres of the added sera there are better or worse
expressed serological differences between NB and the other CMV strains. There is a possibility for this strain to belong
to another (third) CMV serogroup. Devergne and Carding describe third serogroup
(Co), which is closer to ToRS (subgroup II) than to DTL (subgroup I) [3].
Some researchers include into third serological group the CMV strains, which
do not differentiate from the representatives of either subgroup I or subgroup
II according to immunodiffusion tests but behave as subgroup II strains in
hybridization and ELISA [31]. The investigated Bulgarian strain NB can not
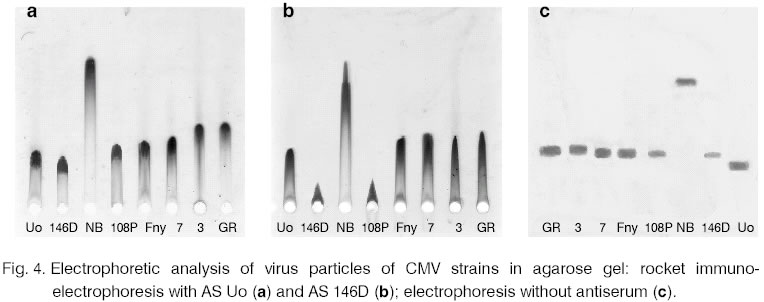
be referred to any of these serotypes. For further characterization of the NB serological
relationship rocket immunoelectrophoresis with AS Uo and AS 146D was performed
(Fig. 4 a, b). CMV strains did not divide into
subgroups at the applied concentrations of the virus preparations. The American
strains from subgroup II (3 and GR) showed weaker relationship (longer "rockets")
with AS Uo but with AS 146D the precipitation figure was as those of Fny
and 7. With the second antiserum the Bulgarian strains 146D and108P from
subgroup II exhibited the strongest reaction. The other viruses did not differentiate
significantly. Both experiments, however, revealed big differences in the
precipitation figures of NB - its "rockets" were twice as long.
The electrophoretic patterns obtained as a result from the tests with both
sera suggest that the weakest relationship exists between NB and the corresponding
strains homologous to these sera. The rocket immunoelectrophoresis data directed
investigations to the testing electrophoretic mobility of the NB virus particles
(Fig. 4 c). These results showed significant
difference between the mobility of NB and the other tested viruses. The NB
strain moved much faster than the other strains, which did not differ significantly.
Only with Uo smaller electrophoretic mobility was observed. This analysis
turned out to be of big importance for the NB testing because in case of
such big differences with the other strains the comparative immunoelectrophoretic
analyses are considered to be unsuitable. This explains the NB behaviour
upon rocket immunoelectrophoresis as well as the different position of the
precipitation lines in the immunodiffusion and counter immunoelectrophoresis
experiments. In the latter all the strains, regardless of their belonging
to a certain group, produced spurs towards NB as a result of their
smaller electrophoretic mobility. In some other papers very small differences
(up to several mm) in the CMV virion mobility are reported. According to Lot and
Kaper the intact virions from subgroup II move faster than CMV-I [17]. In
Hanada experiments, however, the mobility of the virus particles does not
correlate with their serological specificity [8]. In our research the migration
of the virions in electric field is not connected with strain subgroup belonging. The electrophoretic mobilities of the capsid
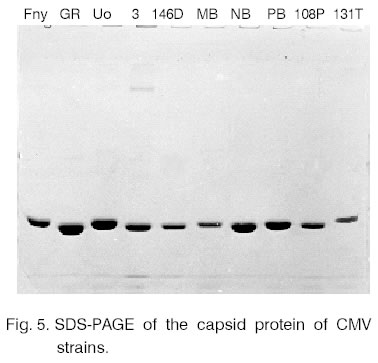
proteins of the NB and the other CMV strains were also compared (Fig.
5). The observed differences were rather strain specific. NB was from
the strains with bigger electrophoretic mobility of the capsid protein. Close
values were observed for GR and 3 (subgroup II) and for PB and Uo (subgroup
I). Literature data for the capsid protein mobility are contradictory. According
to Daniels and Campbell the capsid protein of the isolates from subgroup
II moves faster than the one from subgroup I [2]. According to other authors
the results are the opposite [17]. These differences can be explained with
the usage of different purification procedures. A bigger number of strains
are required for such comparative research. Based on the results of the electrophoretic
analysis it can be concluded that there are single amino acid substitutions
with acidic residues in the capsid protein of the NB strain. They are situated
on the surface of the virus particles. As a result negative charges accumulate
on the intact virion surface, which changes significantly their electrophoretic
mobility in the presence of the applied buffers (alkaline medium). The research of the NB strain shows that it significantly
differs from the other CMV strains. According to ELISA and the immunodiffusion
tests it belongs to subgroup I, as well as according to RFLP-PCR [19] but
it also shows a number of specific serological and electrophoretic properties. The comparison between NB and the other Bulgarian
strain - 146D, isolated from tomato and causing tomato necrosis [11, 34],
is of big interest. Both strains differ significantly not only serologically
and electrophoretically but also with the symptoms they induce in indicator
plants [27, Christova, unpublished data]. There are two tomato necrosis syndromes
caused by CMV: tomato necrosis (or tomato lethal necrosis) and fruit necrosis
[1, 30]. The first syndrome is caused by subgroup II strains while the second
one involves some subgroup I strains, which normally don’t provoke symptoms
on the vegetative parts of the plants or induce weak symptoms. The presence
of satellite RNA is also connected with the fruit necrosis [1]. 146D strain
(subgroup II) provokes the first mentioned above necrosis syndrome while
NB, containing satellite RNA [28] - fruit necrosis. Acknowledgements. The authors express
their gratitude to Prof. Campbell, Prof. Hewlard and Prof. Cheasin, USA,
for the kindly granted strains; to Prof. Rabenstein, Germany,
for the monoclonal antibodies and antiserum N; as well as to Ass. Prof. Yanculova
for the preparation of IgG. The research was financed by Scientific Research
Fund of the Ministry of Education and Science (contracts K-526, CC-616 and
B-603). References
- Crescenzi, A., L.
Barbarossa, D. Gallitelli, G. P. Martelli, 1993. Plant Dis., 77, 28-33.
- Daniels, J., R. N. Campbell, 1992. Plant
Dis., 76, 1245-1250.
- Devergne, J. C., L. Cardin, 1973. Ann.
Phytopathol., 5, 409-430.
- Dikova, B . Viral diseases in cucumber. Ph.
D. Thesis (in Bulgarian).
- Francki, R. I. B., D. W. Mossop,
T. Hatta, 1979. CMI/AAB Descriptions of plant viruses, No. 213.
- Haase, A., F. Rabenstein, 1988. Arch. Phytopathol.
Pflanzenschutz, 24, 167-169.
- Haase, A., J. Richter, F. Rabenstein,
1989. J. Phytopathol., 127, 129-136.
- Hanada, K., 1984. Ann. Phytopathol.
Soc.
Japan, 50,
361-367.
- Hristova, D., K. E. San, 1995. Plant
Science (Sofia), 32, 84-89.
- Hristova, D., A. Yordanova,
2001. Plant Science (Sofia), 38, 56-63.
- Hristova, D., A. Yordanova,
V. Mavrodieva, 2002. J. Phytopathol., 150, 334-339.
-
Kaper, J. M., D. Gallitelli,
M. E. Tousignant, 1990. Res. Virol., 141, 81-95.
- Kaper, J. M., H. E. Waterworth,
1977. Science, 196, 429-431.
- Koening, L., 1978. J. Gen.
Virology, 40, 382-398.
- Kovachevski,I., 1965. Cucumber
mosaic virus in Bulgaria.
BAS, 79.
- Laemmli, U. K., 1970. Nature, 227,
680-685.
- Lot, H., J. M. Kaper, 1976. Virology, 74,
209-222.
- Lot, H., J. Marrou, J.
B. Quiot, C. Esvan, 1972. Ann. Phytopathol., 4, 25-38.
- Mavrodieva, V. A., D. J.
Barbara, N. J. Spence, 1998. Ninth Congress of the Bulgarian Microbiologists,
15-17 Oct. 1998, Sofia,
Bulgaria.
Proceedings, vol. 2, 549-552.
- Ouchterlony, O., L. A. Nilsson,
1978. Immunodiffusion and immunoelectrophoresis. In: Weir, D. M. (Ed.), Handbook
of Experimental Immunology, 3rd edn, Oxford: Blackwell
Scientific Publications, 19.16-19.23.
- Owen, J., P. Palukaitis,
1988. Virology, 166,
495-502.
- Palukaitis, P., M. J. Roossinck,
R. G. Dietzgen, I. B. Francki, 1992. Adv. Virus. Research, 41,
282-349.
- Piazzolla, P., J. R. DiazRuiz,
J. M. Kaper, 1979. J. Gen. Virol., 45, 361-369.
- Richter, J., K. Schmelzer,
E. Proll, 1971 Arch. Phytopathol. Pflanzenschutz, 8,
421-428.
- Rizo T. M., P. Palukaitis,
1988. J.
Gen. Virology, 69, 1777-1787.
- Stoimenova, E., 1995. Plant
Science (Sofia), 32, 90-92.
- Stoimenova, E. 1995. J. Culture
Collections (Sofia), 1,.46-52.
- Stoimenova, E., A. Yordanova,
V. Mavrodieva, 1997. Compt. Rend. Acad. Bulg. Sci., 50, 85-88.
- Takanami, Y., 1981. Virology, 109,
120-126.
- Varveri, C., K. Boutsika,
1999. Plant
Pathol., 48, 95-100.
- Wahyuni, W. S., R. G. Dietzgen,
K. Hanada, R. I. B. Francki, 1992. Plant Pathology, 41,
282-297.
- Wallenberg, B., U. B. Andersson,
1978. Immunoelectrophoretic Techniques with LKB 2117 Multiphor.
LKB Application Note 249.
- Yordanova, A., D. Hristova,
1995. J. Culture Collections (Sofia), 1, 42-45.
- Yordanova, A., D. Hristova,
V. Mavrodieva, 1999. Plant Science (Sofia), 36, 230-234.
Copyright 2002 - National Bank for Industrial Microorganisms and Cell Cultures
- Bulgaria
The following images related to this document are available:
Photo images
[cc02013f3.jpg]
[cc02013f2.jpg]
[cc02013f4.jpg]
[cc02013f1.jpg]
[cc02013f5.jpg]
|

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}