 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Journal of Culture Collections, Volume 4, No. 1, 2004-2005, pp. 53-60 ISOLATION AND CHARACTERIZATION OF BACTERIOCIN PRODUCING LACTIC ACID BACTERIA FROM A SOUTH INDIAN SPECIAL DOSA (APPAM) BATTER Vijai Pal, Marilingappa Jamuna and Kadirvelu Jeevaratnam* Food Biotechnology Division, Defence Food Research Laboratory, Siddarthanagar, Mysore-570 011, India Code Number: cc05007 Summary Lactic acid bacteria (LAB) commonly used as starter cultures in food are known to produce antimicrobial substances such as bacteriocins, having great potential as food biopreservatives. The present study was aimed at isolating bacteriocinogenic LAB from a traditional fermented food, “Appam”. Twenty five colonies of LAB were isolated and screened for bacteriocin production potential. Ten isolates showed good antimicrobial activity against Gram-positive viz., Bacillus cereus, Staphylococcus aureus, Listeria monocytogenes as well as Gram-negative viz., Pseudomonas aeruginosa, Vibrio parahaemolyticus and Aeromonas hydrophila microorganisms. During characterization of these strains two of them were found to resemble two other strains. One of the bacilli has been reported earlier. Therefore, in this study characterization of 7 isolates through morphological, physiological, biochemical and carbohydrate fermentation tests as well as cell wall protein profiles have been reported. Five of them were cocci, identified as Pediococcus pentosaceous (4 strains) and the other as Tetragenococcus halophilus, while the remaining two being rods and were found to be Lactobacillus plantarum. Introduction Isolation and screening of microorganisms from naturally occurring processes have always been the most powerful means for obtaining useful cultures for scientific and commercial purposes [21]. This certainly holds true for lactic acid bacteria (LAB), which are used throughout the world for manufacture of a wide variety of traditional fermented foods. Since they are involved in numerous food fermentations, known to man for millennia, it is assumed that most representatives of this group do not pose any health risk to man, and are designated as GRAS (Generally Recognized as Safe) organisms. The LAB, generally considered as “food grade” organisms, show special promise for selection and implementation as protective cultures. There are many potential applications of protective cultures in various food systems [10]. These organisms have been isolated from grains, dairy and meat products, fermenting vegetables, and the mucosal surfaces of animals [15]. During lactic acid fermentation these bacteria not only have their effect on food and flavour but they are also known to produce and excrete compounds with antimicrobial activity, such as bacteriocins. Bacteriocins of LAB are considered as safe natural preservatives or biopreservatives, as it is assumed that they are degraded by the proteases in gastrointestinal tract [5]. Bacteriocins are extracellularly released peptides or protein molecules, with a bactericidal or bacteriostatic mode of action against closely related species. The inhibitory spectrum of some bacteriocins also includes food spoilage and/or food-borne pathogenic microorganisms [17]. The discovery of nisin, the first bacteriocin used on a commercial scale as a food preservative dates back to the first half of last century but research on bacteriocins of LAB has expanded in the last two decades, searching for novel bacteriocin producing strains from dairy, meat and plant products, as well as traditional fermented products. Many bacteriocins have been isolated and characterized [5]. Earlier, we have reported the characterization of LAB isolates, a bacillus from appam batter and another from vegetable pickle as well as characterization of the partially purified bacteriocins from the same [11]. Present investigation reports on the isolation and characterization of the other bacteriocin producing LAB from appam (special dosa) batter. Materials and Methods Chemicals. Analytical grade chemicals and dyes were obtained either from SRL, India or SD Fine Chemicals, India while the proteolytic enzymes, molecular weight markers and bacteriological media were obtained from Sigma, USA and Hi-Media, India respectively. Microorganisms. The indicator organisms,
viz., Bacillus cereus, Staphylococcus aureus, Listeria monocytogenes,
Pseudomonas aeruginosa, Vibrio parahaemolyticus and Aeromonas hydrophila
were procured from Microbial Type Culture Collection (MTCC) at Institute of Isolation and screening of LAB for antimicrobial activity. LAB were isolated from the appam batter by appropriate dilutions with saline, plated on MRS (de Mann Rogosa Sharpe) agar and incubated anaerobically at 37 oC for 2-3 days. Twenty five well-isolated colonies were picked up and transferred to MRS broth. They were propagated twice and streaked on MRS agar to check the purity of the isolates and then stored in MRS soft agar (0.5 %) overlaid with 50 % glycerol at -20 oC. These 25 isolates from appam batter were tested for their ability to produce bacteriocins. The isolates maintained in frozen stocks were propagated twice in MRS broth and used for further study. These were inoculated into MRS broth and incubated at 37 oC for 48 h. Cell free supernatants adjusted to pH 5.0 with 2 N NaOH, were concentrated to one tenth of the original volume by flash evaporator, sterilized by passing through 0.22 mm membrane filter (Millipore, India) and evaluated for antimicrobial activity by agar well diffusion method [20] against B. cereus, St. aureus, L. monocytogenes, Ps. aeruginosa, V. parahaemolyticus and A. hydrophila as indicator organisms. Characterization and differentiation of bacteriocin producing LAB. Bacteriocin producing strains were Gram stained and examined microscopically for cellular morphology and Gram-stain phenotype. Catalase activity was tested by spotting colonies with 3 % hydrogen peroxide. Growth was assayed in MRS broth at 10, 15, 37 and 45 oC as well as at pH of 4.4, 5.0, 8.6 and 9.0 incubated at 37 oC. Salt tolerance was tested with 6.5, 10 and 15 % (w/v) NaCl in MRS broth. Production of acid and CO2 from glucose was tested in MRS broth containing Durhams tube, with citrate omitted [18] and in Gibson’s semi-solid tomato-juice medium by the method of Gibson and Abd-el-Malek [9]. Production of ammonia in MRS broth omitting glucose and meat extract, while containing 0.3 % arginine and 0.2 % sodium citrate replacing ammonium citrate, was monitored using Nessler’s reagent. Homo- and hetero- fermentative differentiation test was also carried out according to the method reported by Zuniga et al. [24]. Assays for gelatin hydrolysis and nitrate reduction were performed in accordance with Harrigan [9]. Ability to ferment various carbohydrates was evaluated using MRS broth supplemented with filter sterilized sugar solutions to a final concentration of 1 % w/v and 0.004 % chlorophenol red without glucose and meat extract [18]. The configuration of lactic acid formed from glucose was determined enzymatically using D-lactate and L-lactate dehydrogenase. Antibiogram of LAB isolates. The isolates were inoculated into MRS broth individually and incubated for 24 h. About 25 ml of MRS agar was seeded with the cultures of LAB isolates (106 CFU/ml) mixed well, poured into sterile Petri plates and stored at 4 oC for 1 h to solidify the media. OCTA-discs (8 antibiotics in a single ring) were placed up side down, pressed on the top of the agar plates and kept again at 4 oC for 1 h. The plates were incubated at 37 oC over night. Resistance was defined as the absence of a growth inhibition zone around the discs. Cell wall protein extraction and analysis. LAB isolates were grown in MRS broth and cells harvested and resuspended in 3 ml distilled water (A600nm=2) and centrifuged. Cell wall proteins were extracted from final pellets with 0.5 ml of 0.01 M Tris-HCl, 0.01 M EDTA, 0.01 M NaCl, 2 % SDS, pH 8.0, at 100 oC for 5 min [8]. After treatment suspensions were centrifuged at 11 600 g for 10 min and supernatant fluids were examined using Tris-Glycine SDS-PAGE [14] with stacking gel of 5 % and separating gel of 20 % acrylamide – bisacrylamide. Results and Discussion Ten out of 25 randomly picked colonies exhibited good antimicrobial activity against Gram-positive bacteria, viz., B. cereus, St. aureus and L. monocytogenes as well as Gram-negative bacteria, viz. Ps. aeruginosa, V. parahaemolyticus and A. hydrophila and were studied further. It was observed that two of the strains resembled with other two in all aspects of characterization and hence we were left out with 8 bacteriocin producing LAB. The investigation of 7 LAB is presented in this study. All the 7 strains were found to be Gram-positive while morphologically DFR JJ1, 6, 9, 10 and 11 were cocci and DFR JJ4 and DFR JJ5 were bacilli. Physiological and biochemical properties as well as carbohydrate fermentation pattern of these strains are shown in Tables 1 and 2. On the basis of morphological, physiological and biochemical characters as well as sugar utilization pattern DFR JJ1, 6, 9 and 10 were identified as Pediococcus pentosaceous, DFR JJ11 as Tetragenococcus halophilus, DFR JJ4 and DFR JJ5 as Lactobacillus plantarum. The four strains of P. pentosaceous were Gram-positive cocci, arranged in tetrads and pairs. These strains were catalase-negative, did not produce CO2 from glucose, homofermentative on HHD medium, did not reduce nitrate and produced both L-(-) and D-(+) form of lactic acid (Table 1). These are also the major characters of genus Pediococcus as reported earlier [4, 6, 7]. These four P. pentosaceous strains produced NH3 from arginine and did not hydrolyze gelatin. Only two species of Pediococcus genus, viz., P. acidilactici and P. pentosaceous were able to hydrolyze arginine [3, 6, 7]. Luxurious growth was observed for all 4 strains at 37 oC while it was weak at 10 oC and none could grow at 45 oC (Table 1). P. acidilactici can arbitrarily be differentiated from P. pentosaceous by its ability to grow at 50 oC and survive even 70 oC for 10 min [7, 16]. Carbohydrate fermentation pattern of these strains showed that all the four strains were able to ferment maltose, trehalose, xylose, cellobiose, and galactose but could not ferment sorbitol, mannitol, glycerol and arabinose (Table 2). Ability to ferment maltose is a special character of P. pentosaceous, while P. acidilactici could not ferment it [3, 6, 7]. On the basis of all these properties the four strains were identified as P. pentosaceous, however they were different strains since they distinguished among themselves regarding sugar fermentation pattern as well as their resistance and sensitivity towards various antibiotics (Table 3). T. halophilus strain cells were found to be Gram-positive cocci occurring in tetrads and pairs and exhibited weak reaction for catalase. Presence of pseudocatalase in some strains of Pediococcus and Tetragenococcus lead to catalase-positive reaction [19]. Strain DFR JJ11 did not grow at pH 4.4 but grew luxuriously at pH 8.6 and in 15 % NaCl. It was found to be homofermentative, hydrolyzed arginine and did not produce CO2 from glucose (Table 1). This strain did not ferment arabinose, sorbitol and glycerol but easily fermented sucrose, maltose, mannose and trehalose (Table 2). Garvie [7] and Villar et al. [23] also reported similar results for P. halophilus, which later was renamed as T. halophilus [19]. This strain belongs to family Aerococcaceae which is considered a LAB from a practical food technology point of view [2]. T. halophilus strain was highly resistant to streptomycin and moderately resistant to nalidixic acid, cotrimazine and colistin, while it was sensitive for all other antibiotics examined (Table 3). Table 1. Morphological, physiological and biochemical properties of bacteriocin producing LAB isolates.
Legend: growth (+), no growth (–), luxurious growth (++), weak growth (w), homofermentative (Ho), very weak (vw), not determined (ND), weak reaction due to pseudocatalase (+1), glucose (Glc), arginine (Arg). Table 2. Carbohydrate utilization pattern of the bacteriocin producing LAB isolates.
Legend: growth (+), no growth (–), delayed fermentation (+*). Both strains of Lb. plantarum, viz., JJ4 and JJ5were found to be Gram-positive, catalase-negative bacilli. Both the strains were homofermentative and did not produce CO2 and ammonia from glucose and arginine respectively. Their growth was weak at 15 oC and pH of 4.4, while both grew luxuriously at 37 oC and pH of 8.6. The two strains produced both L-(-) and D-(+) form of lactic acid (Table 1). Among sugars examined both strains fermented mannitol, melibiose, esculine, galactose, sorbitol, sucrose, raffinose and trehalose but they could not ferment arabinose, glycerol and xylose. Lb. plantarum JJ5 was able to ferment rhamnose, however, the same was not utilized by other strain JJ4 (Table 2). Similar characters for Lb. plantarum have been observed earlier [13, 22]. The behavior of these two strains against various antibiotics was also found to be similar (Table 3). Table 3. Antibiogram of bacteriocin producing LAB isolates determined by antibiotic sensitivity OCTA-discs.
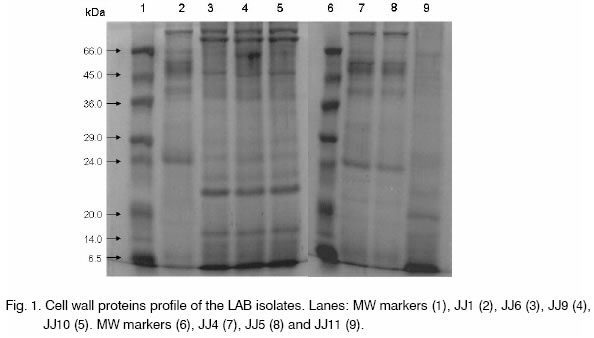
Legend: sensitive (S), resistant (R), moderate (M). Cell wall protein extraction and analysis. Cell wall protein profiles for 7 bacteriocin producing LAB isolates are depicted in Fig. 1. Two major bands of >66 kDa were present in case of DFR JJ6, 9 and 10, while only one band was observed for DFR JJ1, 4 and 5. However, these bands were absent in DFR JJ11. One band of ~66 kDa was prominent in DFR JJ1 and DFR JJ9 but not in DFR JJ6. DFR JJ1 profile shows a band of ~40 kDa while a band of ~38 kDa could be seen for DFR JJ6, 9 and 10. Bands of ~21.5 kDa and 15 kDa were prominent in DFR JJ6, 9 and 10, while absent in DFR JJ1. The profile of DFR JJ4 and 5 showed much resemblance, both being Lb. plantarum, while the strains DFR JJ6, 9, 10 showed similar profile, all being P. pentosaceous, however, DFR JJ1 being P. pentosaceous differed considerably. The profile of DFR JJ11 was found to be differed from others and this was characterized as T. halophilus. In conclusion, eight LAB isolates from appam batter, capable of producing good amount of bacteriocins have been anticipated to have enormous potential for food applications as biopreservatives. Among these strains, characterization of one lactobacillus isolate and its bacteriocin as well as its potential in preserving vegetarian food systems has been reported earlier [11, 12]. Studies on the production, purification and characterization of bacteriocins from other isolates are in progress. Acknowledgements. The authors are thankful to Dr. A. S. Bawa, Director, Defence Food Research Laboratory, Mysore for his keen interest and providing the facilities. References
Copyright 2005 - National Bank for Industrial Microorganisms and Cell Cultures - Bulgaria The following images related to this document are available:Photo images[cc05007f1.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}