 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Journal of Culture Collections, Vol. 6, No. 1, 2009, pp. 42-51 Study of antibacterial activity of Lactococcus lactis against spoilage psychrotrophic bacteria isolated from refrigerated raw milk Mélika Mankaï1*, Olfa Ben Moussa1, Chockry Barbana1, Imen Dhouib1, Mnasser Hassouna1, Nuno Bartolomeu Alvarenga2 and João Canada2 1Unité de Recherche « Sciences et
Technologies des Aliments », École Supérieure des Industries
Alimentaires de Tunis (ESIAT), 58 Avenue Alain Savary 1003 El Khadhra – Tunis,
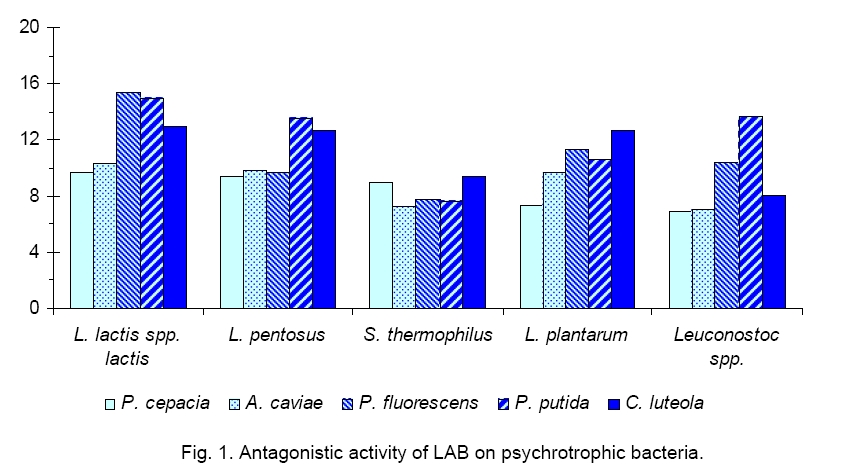
Tunisia Code Number: cc09006 Summary Effect of Lactococcus lactis ssp lactis inoculated in UHT skim milk on the proteolytic activity of psychrotrophic strains was studied. The titration of a purified non nitrogenous fraction from refrigerated UHT milk samples, previously inoculated by 106 psychrotrophic strains, showed that Pseudomonas fluorescens and P. putida were the most proteolytic species. This result has also been confirmed using polyacrylamide gel electrophoresis of casein fractions extracted from refrigerated and contaminated UHT milk samples. All extracellular proteases of the studied germs revealed a decreasing affinity towards κ, β, and αs1-caseins. This bacterial proteolysis of the UHT milk caseins samples, voluntarily contaminated by each of the studied psychrotrophic strains, was significantly reduced following the addition of 106 CFU/ml Lactococcus, which caused, after 9 days of refrigeration, an appreciable reduction of the number of psychrotrophic germs from a minimum of 100 to 1000times, depending on the tested strain. Our results suggest that the inhibitory effect of lactic acid bacteria on psychrotrophs is due to acidification or/and bacteriocin like production. Key words: Lactococcus lactis, psychrotrophic bacteria, UHT milk Introduction Many microorganisms can spoil milk and dairy products [16, 19]. During milk storage by refrigeration psychrotrophs can proliferate. These bacteria hydrolyse milk proteins and lipids through the secretion of a variety of heat stable extracellular proteases and lipases, which cause different defects in dairy products [24] as curdling in UHT [11] or pasteurised milk [7], various off-flavours [9], abnormal texture and reduced cheese yields [28]. It has been shown that proteinases and lipases from psychrotrophs can be inactivated by thermal treatment between 50 °C and 65 °C. For cheese obtained directly from raw milk, this solution cannot be admitted, for this reason alternative solutions to inhibit psychrotrophs have been considered e.g. use of lactic acid bacteria (LAB); activation of the lactoperoxidase system (LPS) that needs the presence of 2 factors, hydrogen peroxide and thiocyanate to develop its anti-microbial function. The main functions of lactic acid bacteria (LAB) are the transformation of lactose to lactic acid, flavour synthesis and direct contribution to the final texture of dairy products [22]. Diverse strains of LAB are commonly used as starters in the transformation of a variety of cheeses. These bacteria participate in the proteolysis during cheese ripening especially in hydrolyzing short and medium peptides into amino acids precursors of aroma compounds, contributing to some sensorial and textural properties of cheeses [26]. Among LAB strains, different species belonging to Lactococcus lactis are widely used in dairy manufacture. The proteolytic system of L. lactis is composed of a cell-wall proteinase and ten intracellular peptidases. L. lactis contributes to final flavours of cheese by synthesis at the ripening stage of a wide variety of branched-chain alcohols and aldehydes resulting from branched-chain amino acids catabolism [10]. On the other hand, various studies have shown that L. lactis is a bacteriocin producer, specifically of Nisin that is used as a natural preservative in some foods. Also LAB are able to produce other antimicrobial substances such as lactic acid, diacetyl, carbon dioxide and hydrogen peroxide [3]. The aim of this work was to study the inhibition of spoilage gram negative psychrotroph bacteria growth by LAB. This study has been performed by inoculation of UHT skimmed milk with different stains of psychrotroph bacteria isolated from refrigerated raw milk and determination of their growth inhibition after addition of L. lactis ssp. lactis by control of casein proteolysis. Materials and Methods Materials. API 20 NE and API 50 CH were obtained from Biomerieux (France). PCA (Marnes-La-Coquette, France) and MRS (Beauvais, France) agar were purchased from Biorad. Nutritive broth was from Biorad (Hercules, USA). Proteins standards used in SDS-PAGE were from Biorad (France). Proteinase k (Carisbad, Germany) and Trypsin (Steinhein, Switzerland) were provided from Fluka. Trichloroacetic acid (TCA) (Darmstadt, Germany) and UHT bovine milk samples were kindly supplied by Thalja S.A. (Tunis, Tunisia). Strains and culture conditions. Bacterial strains used in this study wereisolated from refrigerated raw milk and were identified as Aeromonas caviae (36%), Pseudomonas fluorescens (29%), P. putida (11%), P. cepacia (3%) and Chryseomonas luteola (15%) in our laboratory [18]. They were classified using the API 20 NE. Phenotypic characteristics of these strains were treated with Statistica software version 5.0. (1997). Strains of LAB were isolated from milk and dairy products (50 strains). They were identified based on Gram staining, catalase, oxidase tests and fermentation of 49 carbohydrates (API 50 CH). Both isolates of Gram-negative germs and LAB were subcultured twice (1% inoculum, 24 h, 30 °C) in 15 ml nutritive broth (Biorad) and kept frozen at -20 °C in nutritive broth supplemented with 10 % glycerol. Inoculation of milk. Single P. fluorescens, P. putida, P. cepacia, C. luteola or A. caviae was inoculated in 500 ml UHT skim milk at 106 CFU/ml with and without L. lactis ssp. lactis (106 CFU/ml) and stored at 4 °C during 9 days. Bacterial counts. Bacterial populations were estimated by standard methods (Guiraud, 1998). Psychrotrophs and mesophilic bacteria were inoculated on PCA and incubated respecttively at 7 °C for 10 days and 30 °C during 72 h. LAB were plated on MRS and incubated 72 h at 37 °C. Antibacterial activities. Antibacterial activity of LAB was tested against Gram-negative strains [27]. 5 µl of an overnight culture of LAB was put on MRS agar and incubated for 24 h at 30 °C. Later, 5 ml of PCA agar (0.75 % agar) inoculated with 200 µl of an overnight indicator microorganism was poured on MRS spots. The plates were then incubated for 24 h at 30 °C and were subsequently examined for zones of inhibition. Characterization of the antibacterial compounds. LAB cultures broth samples were centrifuged ( Physico-chemical analysis. pH of UHT milk samples was measured. Proteolysis of milk caseins was detected by measuring the levels of trichloroacetic acid (TCA) soluble peptide determined by Kjeldahl method [21]. Polyacrylamide gel electrophoresis (SDS-PAGE) was performed using 15% gels as described by Laemmli (1970) [14]. Preparation of protein samples for SDS-PAGE was conducted according to the Addeo methods [2]. Samples were taken at regular intervals for the determination of total viable counts, psychrotrophics flora, mesophilic lactic flora, trichloroacetic acid (TCA) soluble peptide and for SDS-PAGE. Statistical analysis. Analysis of simple linear correlation coefficients was performed using SPSS 8.0 software (Chicago, USA). Statistical significance level was defined as P < 0.05. Results and Discussion The inhibition of spoilage gram negative psychrotroph bacteria growth by LAB was studied in the present work. The majority of the psychrotrophic strains identified in raw milk were Gram-negative accounting for nearly 80 % of the total psychrotrophic microflora belonging to the two genera Pseudomonas (58 %) and Aeromonas (36 %), independently of the refrigeration storage period[18]. Antagonistic activity of each LAB of five dominant
strains, isolated from raw milk was tested on gram-negative bacteria. LAB
strains used were: L. lactis spp. lactis (62 %), Lactobacillus
pentosus (6 %), L. plantarum (12 %), Streptococcus thermophilus (4 %)
and Leuconostoc spp. cremoris (16 %). LAB gave circular inhibition areas onto the
indicator spoilage strains tested (Figure 1). The diameters of these inhibition
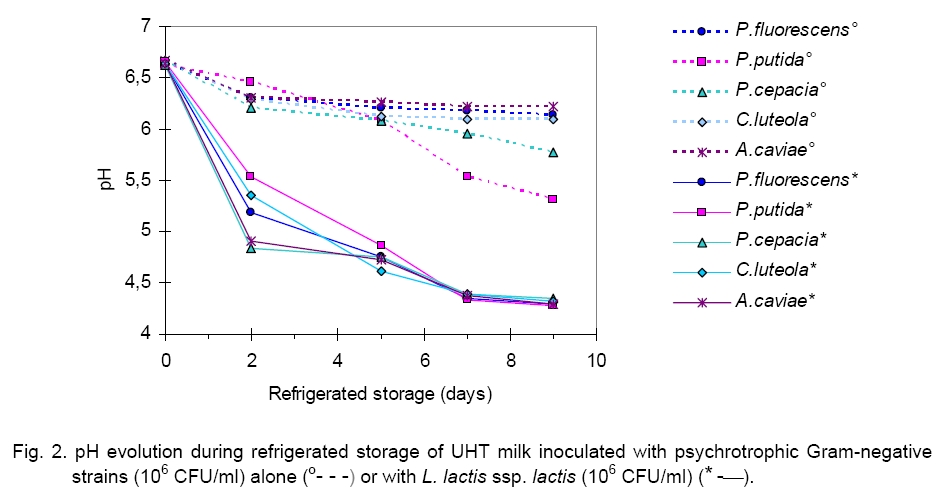
areas were estimated to be between Heating at 80 °C or 100 °C for 15 min did not induce denaturation of antibacterial compounds. Otherwise, it is interesting to note that bactericide activity was found to be stable at pH of 5 and 7 (data not shown). Stability reduction was observed by many authors [20], who reported that inhibitive substances of some LAB strains could be related with both lactic acid and proteinaceous antibacterial compounds [23]. In this study we observed that the pH of milk samples inoculated with the psychrotrophic strains (106 CFU/ml) undergoes a slight reduction. On the other hand, the inoculation with Lactococcus (106 CFU/ml) of raw milk induced an acidification of the medium. The pH decrease primarily takes place during the first two days of storage (1.88 pH units). Beyond this stage and until the 9-th day of refrigeration, the pH of milks remained stable around an average value of 4.3 pH units independently of the tested strain (Figure 2). Recorded reductions are around 0.89 (R = -0.82), 0.51 (R = -0.87), 1.31 (R = -0.98), 0.44 (R = -0.87) and 0.44 (R = -0.95) pH units after 9 days of incubation at 4 °C of UHT milk samples, respectively supplemented with A. caviae, P. fluorescens, P. putida, C. luteola and P. cepacia. These results showed that the most significant reduction of pH was obtained for milk samples inoculated with strains of P. putida. Likewise, when milk samples are inoculated by a mixed culture, composed of a psychrotrophic and a L. lactis ssp. lactis strains, the pH undergoes a more significant reduction. Thus, a diminution of approximately 1.88 pH units in average value is recorded after 2 days of refrigeration at 4 °C of milk independently of the psychrotrophic strain tested. During this stage and until the end of storage, the pH of the milk samples remains stable. Therefore, after 9 days of refrigeration, the pH of the different milk samples reaches the average value of 4.3 pH units. Some authors reported that the addition of 2.5 x 107 Lactococcus/ml to the milk reduced its pH up to the value of 6.54 after 48 h of incubation at 7 °C [8]. Under these conditions, the acidification of milk cannot be responsible for the inhibition of the psychrotrophic bacteria proliferation. On the contrary, in this case, the acidification of milk, being more important, plays a major role in the inhibition of the psychrotrophic bacteria proliferation. Moreover, the results obtained by Desmazeaud showed that the psychrotrophic bacteria cannot grow at pH ≤ 5.4 [12]. We observed that total mesophilic flora revealed a significant development during the refrigeration of raw milk. As it is shown in Table 1 total germs reached, after 9 days of storage at 4 °C, values of about 3 x 107, 2.5 x 1010, 108, 2 x 108, 2.8 x 108 CFU/ml, respectively for milk samples inoculated by P. fluorescens, P. putida, P. cepacia, C. luteola and A. caviae. On the other hand, we observed that the psychrotrophic bacteria content was significantly decreased compared with their population in the UHT milk without L. lactis. Independently of the duration of refrigeration, the linear correlation coefficients were found equal to +0.82 (P. fluorescens), +0.91 (P. putida), +0.96 (P. cepacia), +0.97 (C. luteola) and +0.92 (A. caviae), against respectively +0.99, +0.93, +0.94, 0.94 and 0.90 reported for total flora as shown in Table 2. The concentration of the psychrotrophic bacteria remained high at the end of refrigeration, around an average value of 109 CFU/ml, whatever was the germ inoculated. At these levels, proteases and/or thermoresistant lipases production is important [1].In addition, our results concerning the influence of the addition of lactic strain L. lactis spp. lactis on the development of each of the five psychrotrophic strains showed that total mesophilic flora continues to develop during the refrigeration of milk but at a lower rate compared to that measured in the pilot milk samples not inoculated by the lactic strain. Thus, the number of total germs reached, at the end of 9 days of refrigeration, values about 2 x 109, 2.5 x 1010, 2 x 109, 3 x 109, 109 CFU/ml, respectively for samples of inoculated milk with P. fluorescens (R = +0.50), P. putida (R = +0.63), P. cepacia (R = +0.64), C. luteola (R = +0.52) and A. caviae (R = +0.54). Results obtained show that proliferation of psychrotrophic bacteria increased during the 2 first days of refrigeration, and varied from 3 x 106 to 1.2 x 108, from 2.5 x 106 to 9 x 107, from 1.9 x 106 to 2 x 106, from 3 x 106 to 1.1 x 108 and finally from 3 x 106 to 8.3 x 108 CFU/ml, respectively for samples of inoculated milk with P. fluorescens, P. putida, P. cepacia, C. luteola and A. caviae. Table 1. Microflora development during refrigerated storage of UHT milk inoculated with psychrotrophic strains (106 CFU/ml) with/without L. lactis ssp. lactis (106 CFU/ml).
Table 2. Simple
linear correlation coefficients obtained between spoilage flora inoculated, pH,
non nitrogeneous fraction during storage of UHT milk
at
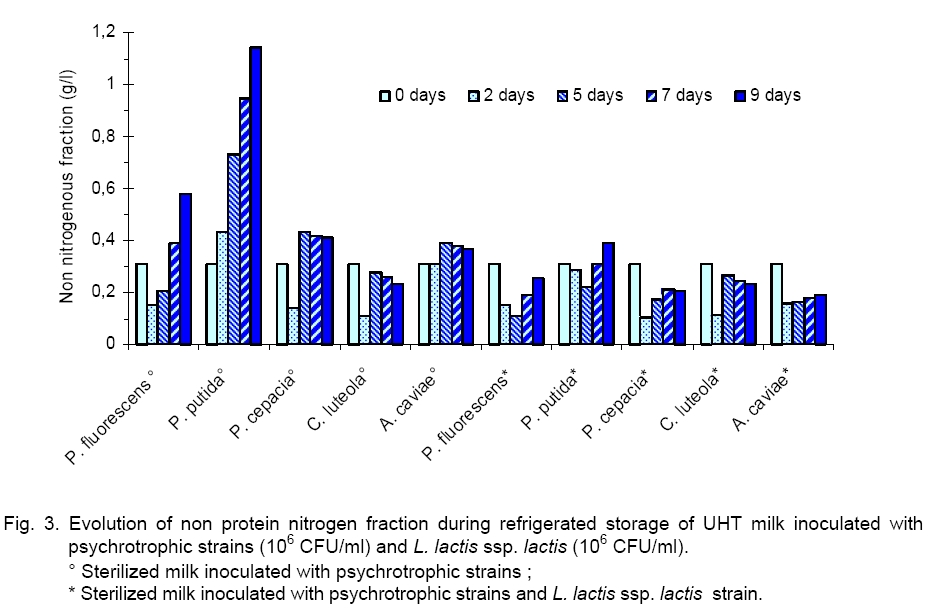
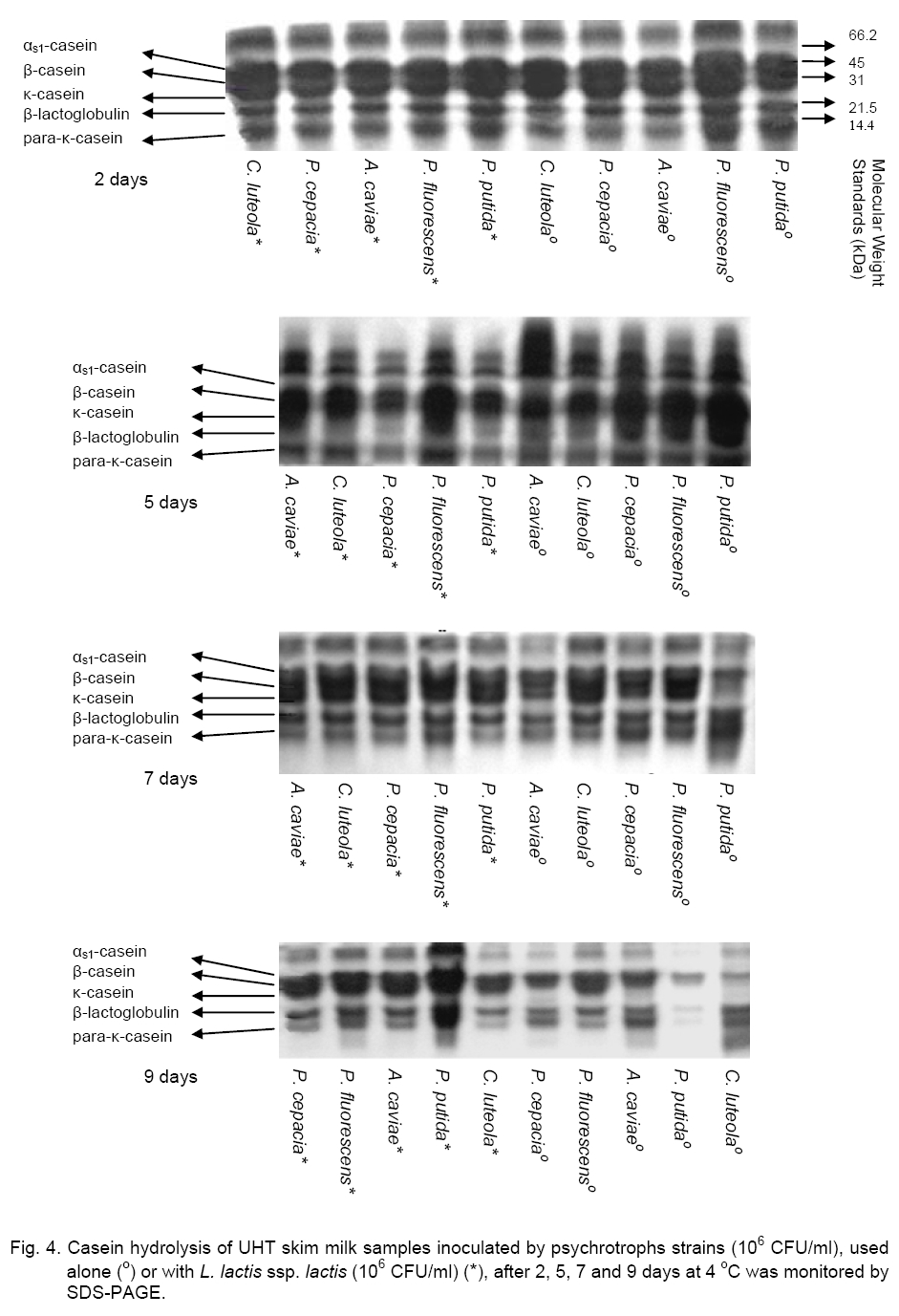
Legend: * level of significance <1 %; ** level of significance <5 %; - sterilized milk inoculated with single psychrotrophic bacteria (106cFU/ml); + sterilized milk inoculated with single psychrotrophic bacteria (106 cFU/ml) and with L. lactis ssp. lactis (106cFU/ml). From this stage and until the end of the storage, except for C. luteola, we observed that the rate of these psychrotrophic germs decreased significantly to reach a value between 105 and 106 CFU/ml according to the tested strain. This result can be explained by an initial cohabitation (maintained for 2 days) of the lactic bacteria and the psychrotophic germs followed by a phenomenon of inhibition exerted by the Lactococcus bacteria on the growth of the psychrotrophic flora, which occurred mainly after the 2-nd day of refrigeration. These observations showed that lactic flora and spoilage flora (Pseudomonas and Aeromonas bacteria) can coexist up to a high level of concentration. In this work, results obtained showed that LAB inhibited the proliferation of the psychrotrophic bacteria by a factor ranging between 102 and 103, depending on the inoculated psychrotrophic strain. These results are in agreement with those obtained previously by Champagne [8] who showed that addition of 2.7 x 107 Lactococcus/ml in sterilized milk reduces P. putida development of about 50 %. Moreover, these authors reported that a concentration of 2.5 x 106 of commercial Lactococcus per ml of milk did not reduce significantly the proliferation of psychrotrophic germs, whereas the addition of 2.5 x 107 CFU/ml decreased clearly the psychrotrophics growth 10-fold. The important decrease of the psychrotrophic bacteria content was related to the acidification of milk observed from the 2-nd day of refrigeration (pH < 4.7). On the other hand, it has been shown that psychrotrophic inhibition depends also on bacteriocin produced by the L. lactococcus ssp. lactis strain as it has previously been described [6]. It has been reported that L. lactis was able to produce Nisin, that is the best known and most studied lanthionine-containing bacteriocin from gram positive bacteria. Nisin has inhibitory effect against some pathogens and spoilage microorganisms, including bacilli, clostridia, corynebacteria, micrococci, pediococci, staphylococci and streptococci [5]. In our study we observed that LAB undergo a significant regular increase throughout the refrigeration of milk. The initial concentrations of LAB varied from 108, 5 x 107, 3.5 x 106, 2.8 x 106 and 1.3 x 106 CFU/ml to reach, at the end of storage, values of 2.6 x 109, 1.5 x 1010, 8 x 109, 3 x 1010 and1.8 x 1010 CFU/ml, respectively for milk samples previously inoculated by P. fluorescens (R = +0.65), P. putida (R = +0.79), P. cepacia (R = +0.77), C. luteola (R = +0.77) and A. caviae (R = +0.91). This result suggests that enzymatic activity of psychrotrophic bacteria can favour the growth of the lactic bacteria which can use the peptides amino acids and ammonia accumulated in milk and produced by the psychrotrophic bacteria [25]. In our work, evolution of proteolysis in milk was studied. The profiles of non protein nitrogen (NPN) fractions were measured by the Kjeldhal method in various milk samples inoculated by each of the five psychrotrophic bacteria strains. As it is seen in Figure 3, results showed that during the first days of storage, the NPN fraction contents of the various UHT milk samples, inoculated with P. fluorescens, P. cepacia and C. luteola, underwent respective decreases of about 50.4, 45.5 and 34.7 %. This decrease could be attributed to the use of this source of NPN at the initial phase of their growth for metabolism. On the contrary, in the milk sample inoculated with P. putida, the NPN undergoes a relatively important increase (39.4 %), where-as this content remained almost stable in the milk sample inoculated with A. caviae. These observations suggest that the NPN does not constitute a limiting factor for the growth of the-se two strains. Beyond this stage and until the end of the refrigeration, the NPN content of the milk samples inoculated by the various psychrotrophic strains increased regularly. The total recorded increases are significant, about 276.2, 164.4, 190.7, 117.5 and 18.2 %, respectively for milk samples inoculated with P. fluorescens (R = +0.71), P. putida (R = +1), P. cepacia (R = +0.67), C. luteola (R = +0.06) and A. caviae (R = +0.80).These results suggest that P. fluorescens and P. putida are the most proteolytic psychrotrophic species [19]. Otherwise, the addition of L. lactis ssp. lactis (106 CFU/ ml) to milk samples, previously inoculated by P. fluorescens, P. putida and P. cepacia, caused à considerable reduction of NPN fraction during their refrigeration. The recorded decreeses are respectively of 209.3, 131.8 and 90 % in relation to samples of milk inoculated with the same psychrotrophic bacteria but in the absence of L. lactis ssp. lactis. We observed that the lactic strain did not seem to reduce the caseins proteolysis of milk samples refrigerated and inoculated previously by C. luteola. The relative variations observed for NPN are the same in milk samples inoculated in either the presence or absence of L. lactis ssp. lactis, respectively 110.7 and 20.1 % (Table 2). These results show that the psychrotrophic bacteria inhibition by L. lactis ssp. lactis depends on the tested strain. Electrophoresis (SDS-PAGE) of casein fractions extracted from refrigerated samples of UHT milk, initially inoculated by the five studied psychrotrophs strains, used alone or in mixed culture with lactic bacteria, confirms data obtained previously on the evolution of the NPN in the time course of the milk samples refrigeration. Thus, electrophoretic profiles of caseins of the UHT milk samples, contaminated previously by every psychrotrophic strain, then refrigerated for 9 days at 4 °C (Figure 4), present a considerable enzymatic hydrolysis of different casein fractions (αs1, β and κ), accompanied with the appearance of weak molecular weights deterioration products. In the same way, we noted an increase in the strip intensity corresponding to the β-casein, because products of its deterioration have electrophoretic mobility close to that of β-casein. Otherwise, stronger deterioration of the caseins has been observed for milk samples inoculated by P. fluorescens and P. putida producing higher quantity of deterioration products, after two days of refrigeration as compared to those sowed by P. cepacia, C. luteola and A. caviae. After 9 days of refrigeration of UHT milk samples contaminated by psychrotrophic strains, these proteolytic bacteria hydrolysed β-casein faster than αs1-casein and readily hydrolysed κ-casein to para-κ-casein. On the other hand, for all analyzed milk samples, the β-casein hydrolysis revealed by electrophoretic mobility showed a major band corresponding to γ-caseins. These observations are in agreement with those observed in previous studies by Law [15] who used SDS-PAGE and reported that the first phase of casein degradation by proteinase from a psychrotrophic strain, during the raw milk refrigeration, resulted in the synthesis of para-κ-casein with a concurrent decrease in the electrophorectic major band intensity of κ-casein while αs1-casein was degraded at a slower rate than β-casein. The κ-casein hydrolysis by psychrtrophs’ proteases was faster than that of β-casein, while the αs1-casein remains less degraded. In the same way, Datte and Deeth [11] comparing the action of Pseudomonas proteases, showed that the chymosine attacks preferentially the κ-casein by hydrolyzing specifically only the peptide bond between Phe105 and Met106, producing hydrophobic para-κ-casein and soluble glycomacropeptide. Our results showed that in all analyzed milk samples, the addition of L. lactis ssp. lactis decreased the casein proteolysis initiated by psychrotrophic proteases Lactoserum proteins seem to be resistant to these proteases (Figure 4). Indeed, P. fluorescens and P. putida degrade the α-lactalbumin and the β-lactoglobuline after only 9 days of the cold milk storage (Figure 4). This result contradicts the works of Law [15] that did not observe lactoserum proteins deterioration products. In conclusion, we showed the importance of the antagonist effect of L. lactis ssp lactis against psychrotrophic bacteria in braking proteolysis phenomena that can cause various defects: curdling in UHT [11] or pasteurized milk [7], various off-flavours [9, 16], abnormal texture, and reduced cheese yield [28]. LAB produce antimicrobial substances such as lactic acid. As perspectives we can complete this study to elucidate inhibition of initial contamination of psychrotrophic microflora gram (-) of refrigerated raw milk for cheese manufacturing by the LAB strains selected on their antagonist power. References
Copyright 2009 - National Bank for Industrial Microorganisms and Cell Cultures - Bulgaria The following images related to this document are available:Photo images[cc09006f1.jpg] [cc09006f4.jpg] [cc09006f2.jpg] [cc09006f3.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}