 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Journal of Culture Collections, Vol. 6, No. 1, 2009, pp. 52-60 Comparative study on characterization of three staphylococcal isolates from varied origin Poulomi Nandy1, Soumen Roy1, Ashoke Ranjan Thakur3 and Shaon Ray Chaudhuri1,2* 1Department of Biotechnology,
West Bengal University of Technology, BF – 142, Sector – 1, Salt Lake,
Kolkata 700064, India; Code Number: cc09007 Summary Two strains of Staphylococcus isolated from urine and sputum samples were compared with a laboratory strain of Staphylococcus. Since all three grew on the same media specific for S. aureus, they were studied for their relatedness by morphological, biochemical and molecular characterization. Variation in properties among the isolates pointed towards their different identity which was confirmed through 16S rRNA gene sequence based phylogenetic analysis. The isolate A from urine was found to be closely related to S. haemolyticus, while isolate P from sputum was S. aureus and isolate S from NICED culture collection was closest to S. epidermidis. Thus the Hicrome aureus agar (Himedia M1468) was not specific for the species S. aureus and could only screen at genus level. The antibiotic profile of the strains was evaluated using a spectrum of 18 antibiotics. The lecithinase activity was retained only in freshly isolated strains and totally ceased in subsequent subculturing. Key words: Staphylococcus aureus, lecithinase, strain characterization, molecular analysis Introduction Urinary tract infections are caused by a variety of bacterial species, the most common being Escherichia coli. Although coagulase negative Staphylococci are among the dominant organisms colonizing the urethra and periurethra in males and females they typically account for less than 10 to 20 percent of all incidences of UTI [5, 15]. Virulence and pathogenicity of Staphylococci are closely related to a wide range of extracellular enzymes and toxins linked with their ability to colonize within the host and to induce lysis of phagocytic cells [16]. Staphylococcus aureus produces one or more enzymes with lipolytic activity but differences between strains have already been reported. Hicrome Aureus agar is a selective medium commonly used for the isolation of S. aureus. The presence of egg yolk [1, 2, 11] in this medium as an added supplement permits the detection of lecithinase reaction [8] due to lipolytic activity of Staphylococcus. This activity appears as a zone of precipitate in the medium surrounding the colonies followed by an iridescent film and a zone of clearing. Lecithinase is a type of phospolipase which is associated with virulence of bacteria [4, 13, 14]. This enzyme usually acts on the animal cell membrane by insertion into the membrane (forming a pore that results in cell lysis) or by enzymatic attack on phospholipids, which destabilizes the membrane [17]. Staphylococci are Gram positive spherical bacteria that occur in microscopic clusters resembling grapes. They can be identified on the basis of a variety of conventional physiological as well as biochemical characters. Staphylococcus is catalase positive, which makes the catalase test useful to distinguish them from other bacteria. S. aureus has also been reported to produce protease, lipase and DNase [18]. A few strains of S. aureus are resistant to all clinically useful antibiotics except vancomycin, and these vancomycin-resistant strains are increasingly being reported. The term MRSA refers to methicillin resistant Staphylococcus aureus. Methicillin resistance is widespread and most methicillin-resistant strains are also showing resistance to multiple other antibiotics. A plasmid associated with vancomycin resistance has been detected in Enterococcus faecalis which can be transferred to S. aureus in the laboratory, and it is speculated that this transfer may occur naturally (e.g. in the gastrointestinal tract) [18]. Emerging vancomycin resistance is probably due to this genetic transfer. In UK, only 2 % of all S. aureus isolates are sensitive to penicillin with a similar picture in the rest of the world, due to the penicillinase (a form of β-lactamase). S. haemolyticus is reported to be resistant to multiple antimicrobial agents including penicillin [10], ampicillin, methicillin, streptomycin, gentamicin, tetracycline, ciprofloxacin. It is sensitive to vancomycin, however the emergence of S. haemolyticus strains with decreased susceptibility to vancomycin is also being reported [12]. This study was an attempt to distinguish two strains of Staphylococcus, the one obtained from urine and another from sputum through detailed microbial characterization in comparison with a laboratory strain of Staphylococcus. Materials and methods Isolation of microorganisms. Two bacterial strains were isolated one each from urine (isolate A) and sputum sample (isolate P), by using Hicrome Aureus agar (Himedia M1468), and Egg Yolk Tellurite Emulsion (Himedia FD046) added as a supplement(as per manufactures protocol), following incubation at 37 ºC for 48 hours under aerobic condition. Another Staphylococcus strain (strain S) was taken from the National Institute of Cholera and Enteric Diseases (NICED) culture collection for comparison and it was also cultivated in the same media. Well isolated colonies were picked up and inoculated into sterile Luria Bertani broth. Morphological characterization. The colony morphologywas monitored, and the Gram nature of the isolates were determined by observing the Gram stained slides of the isolates under 100X magnification of an Axioplus fluorescence microscope by Zeiss (HB50/AC). Biochemical characterization. The presence of enzymes like lipase, protease, oxidase, catalase, lecithinase as well as DNase were tested as described elsewhere using HIMEDIA reagents [9].Additional tests, namely determination of the metabolism type (O/F), susceptibility to lysostaphin [6], lysozyme, haemolysis activity, alkaline phosphatase [7] and test for coagulase [19] were also conducted. Further tests were performed for detecting extracellular as well as intracellular lecithinase activity. Extracellular lecithinase activity using cup assay.
Modified Luria Bertani media plates (tryptone – 1 %, yeast extract – 0.5 %,
sodium chloride – 0.5 %, agar – 1.5 % with egg yolk tellurite
emulsion added as a supplement as per manufacturer’s protocol) were prepared. A
well was punched out in the centre of the plate with the help of a sterile tip.
48 hour old cultures (in LB media) of isolates A, S and P were centrifuged at Intracellular lecithinase
activity using gel diffusion assay. All the three strains were inoculated
in LB broth and it was incubated overnight at 37 ºC. It was
centrifuged at Study of various growth conditions. The growth curve, pH profile, temperature profile and illumination profile were tested as mentioned below. Growth curve. The three isolates were inoculated in LB broth and their growths were monitored for 15 hours in triplicate and the experiment was repeated thrice. The results were plotted graphically with the optical density on the Y axis and the time in hours on the X axis. pH profile. To test the effect of pH variation on the growth pattern of each isolate, they were inoculated in LB broth at pH 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 12 and 14; incubated in a shaker incubator at 37 °C. The growth was monitored after 12, 24 and 36 hours by measuring the optical density value at 660 nm wavelength using spectrophotometer (Beckman Coulter DV – 530, UV Vis spectrophotometer). Temperature profile. Similarly, the strains temperature profile was generated by inoculating the isolates in LB broth, incubating them at 4 ºC, 8 ºC, 20 ºC, 30 ºC, 37 ºC, 40 ºC, 45 ºC and 50 ºC followed by measuring their optical density value at 660 nm wavelength using spectrophotometer, after 12, 24 and 36 hours of incubation. Illumination effect. Growth was also monitored in the presence of continuous source of illumination (13 Watt incandescent lamp from Phillips), complete darkness and alternative cycles of illumination and darkness (each of 12 hours duration). After 24 hours of growth under optimum condition the optical density was measured using spectrophotometer at 660 nm wavelength. Antimicrobial susceptibility testing. The antibiotic resistant characteristics of the three staphylococcal isolates were tested using disk diffusion method. The assay was conducted in triplicate for each organism evaluated. Mueller Hinton Agar (MHA) by Himedia (M173), were used to evaluate each of the microorganisms for antibiotic resistance as per the protocol used elsewhere [9]. The result of clearing zone diameter was interpreted according to the specifications of National Committee for Clinical Laboratory Standards (NCCLS) provided as a chart with the antibiotic kits by Himedia. Molecular characterization. DNA of isolates A, P and S were extracted from overnight grown culture, by using PrepMan Ultra Kit from ABI according to manufacturer’s protocol and sent for partial sequencing of the 16 S rRNA gene. The sequences were subjected to blast analysis. The novel sequence (strain A) was submitted to GenBank. The sequences were subjected to phylogenetic analysis using neighbour joining method. Strains A and S were also sequenced for full length rRNA gene according to the above mentioned protocol using Microseq Full gene Kit from ABI. Results and Discussion Isolation and morphological characterization. Jet black circular colonies of strains A and P surrounded by an immediate opaque zone and an outer clear zone were isolated on Hicrome Aureus agar. Colonies of S were also typically black but did not show lecithinase activity in the same medium. Colonies of A and P had a slimy consistency when touched by an inoculating needle while that of S had a rough appearance with a dry texture. The dry appearance of S can be attributed to the lack of slimy layer production due to long storage under laboratory conditions. Thus glossy appearance of A and P due to extracellular polysaccharide secretion was absent in S. This could also be associated with the lack of lecithinase production which is essential for pathogenic manifestation under clinical condition. Strains A, S and P were Gram positive cocci in clumps, indicating typical staphylococcal nature, but the grape like bunches in A and P were larger in size than those in S. Biochemical characterization. The detailed characterization of the isolates is presented in Table 1. Unlike the reports in the literature [18], all three strains were negative for protease, lipase and DNAse. All three strains were oxidase negative and catalase positive. Organisms positive for the enzyme lecithinase produced typical black coloured colonies in Hicrome Aureus agar (as seen in A, S and P) with a clear zone around it (as seen in A and P). Production of lecithinase was comparatively more in A than P, though the quantity of enzyme production subsequently diminished and ultimately ceased on further propagation for both. There was no detectable lecithinase activity using cell free soup in cup assay. The intracellular lecithinase activity was also found to be absent through gel diffusion assay. One reason that may be accounted for this phenomenon is that lecithinase is an enzyme required for pathogenicity. When growing outside the body under laboratory condition for multiple generations, the need for pathogenic manifestation is lost and so is the enzyme activity. Strain A showed a significant clearing zone around the colonies in the egg yolk media, which could not be reproduced after two generations. In case of strain P, lecithinase activity could be monitored even after two generations, but it was close to negligible. Absence of any detectable lecithinase activity either in supernatant or cell lysate indicates loss of activity due to disuse. All strains were found to grow in both aerobic and anaerobic portions of the OF medium, which supports earlier reports about Staphylococcus sp. Strain P was positive for both slide and tube coagulase assay. Para nitro phenyl phosphate was used as a substrate for the detection of the enzyme alkaline phosphatase which distinguishes S. aureus (positive) from the phenotypically similar S. haemolyticus (negative). All three strains were sensitive to lysostaphin (to different extents) and resistant to 25 µg/ml of lysozyme that was characteristic for the genus Staphylococcus. Haemolysis was observed only in case of isolate A. Table 1. Biochemical characteristics of three Staphylococcus strains.
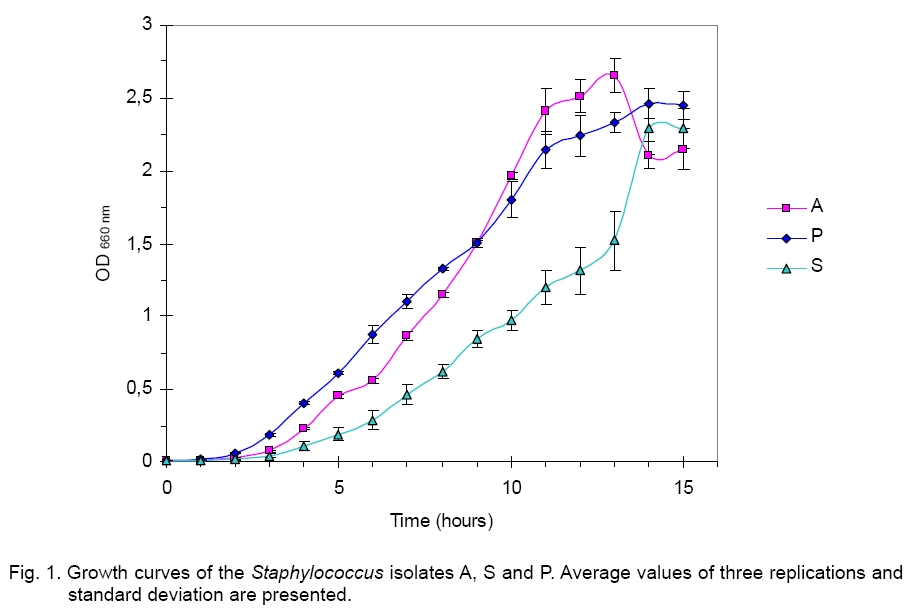
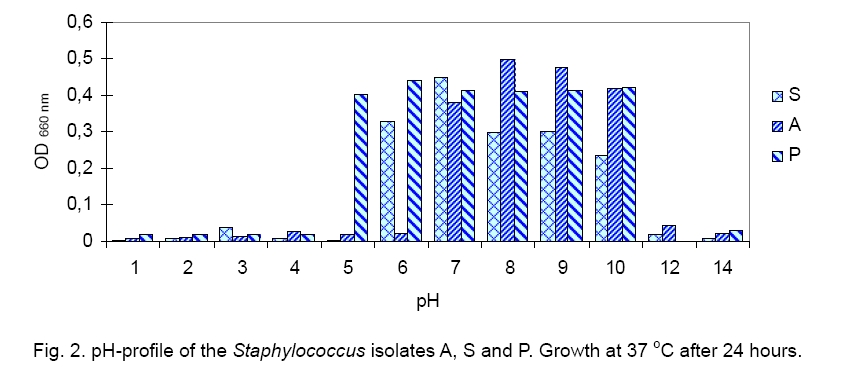
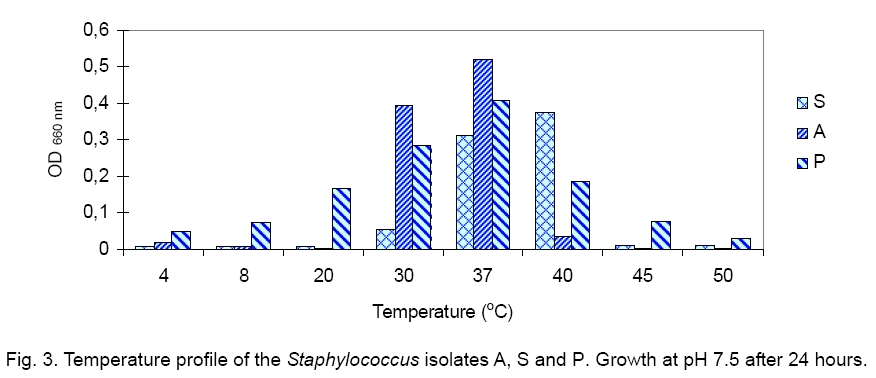
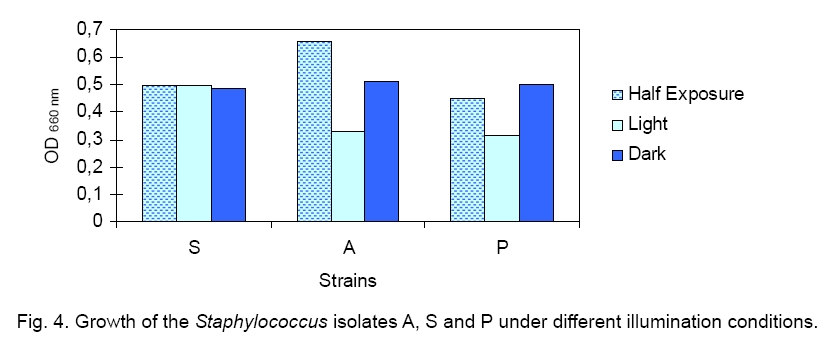
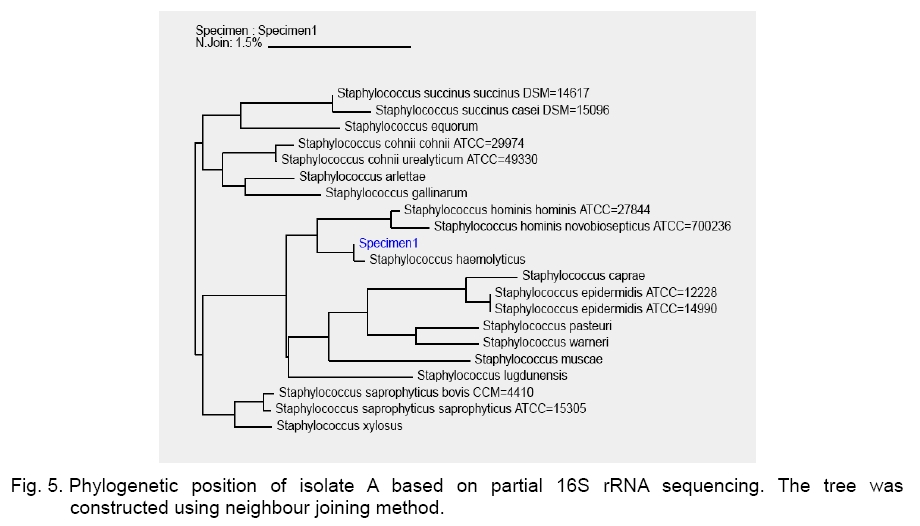
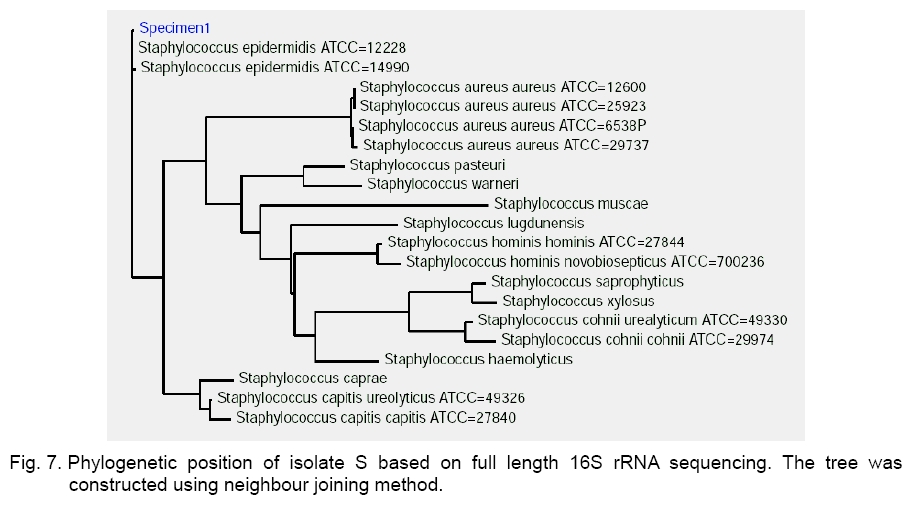
Legend: Performed tests - lipase (Lp), protease (Pr), oxidase (Ox), catalase (Ca) lecithinase (Le); coagulase (Co), DNase and alkaline phosphatase (AP) activities; resistance/sensitivity (+/-) to lysostaphin (Lyn) and lysozyme (Lys); haemolysis (Hae) and O/F test. Growth curve. The growth of strains A, S and P was monitored for 15 hours and there was significant variation in their growth curve patterns as depicted in Figure 1. As per the graph, P has the shortest lag phase while S has the slowest growth rate. The peak growth in case of A is around 13 hours, for P is around 14 hours while for S it is still increasing even at 15 hours. pH and temperature profile. pH and temperature profiling was done to determine the optimum growth conditions as well as the range for growth of the three strains at different temperature and pH. Highest optimum growth in all cases was observed around 24 hours. So, even though the study was conducted after 12, 24 and 36 hours, only the results for 24 hours are depicted here for both pH (Figure 2) and temperature (Figure 3). The result indicates growth of S in pH range of 6 to 12 with optimum growth at 7; growth of A in pH range of 2 to 14 with optimum growth at 8 and growth of P in the range of 1 to 10 with the optimum pH at 6 (Figure 2). The temperature response for S shows growth within 30 °C to 50 °C with optimum growth at 40 °C; for A within 30 °C to 40 °C with optimum growth at 37 °C and for P within 4 °C to 50 °C with optimum growth at 37 °C (Figure 3). Illumination effect. In case of strain S there was no variation in presence or absence of light. Optimum growth condition for A was an alternative cycle of 12 hours illumination and darkness. Presence of continuous illumination was found to have an inhibitory effect on growth of P (Figure 4). Molecular characterization. The data of the partial 16S RNA sequence analysisshowed that strain A was 99.89 % similar to S. haemolyticus, indicating it to be a novel strain of S. haemolyticus. Figure 5 depicts its position in the phylogenetic tree constructed by neighbour joining method. The Accession Number of the novel strain A in GenBank is EF644492. The partial sequence for strain P and S showed 100 % similarity with S. aureus. But strain S was quite different in colony morphology as well as at biochemical level from S. aureus, like it did not show characteristic golden yellow colonies on LB agar plate and was also coagulase negative. Isolate P on the other hand showed golden yellow colonies on LB agar plate characteristic for S. aureus and was positive for coagulase, the confirmatory test for S. aureus. The phylogenetic tree constructed using neighbour joining method for isolate P is depicted in Figure 6. In order to be sure about the identity of the isolates, strains A and S were sequenced for full length16S rRNA gene, which was a confirmatory proof of their identity. Strain A showed a 99.9 % similarity to S. haemolyticus as earlier while strain S showed a 99 % similarity to S. epidermidis (Figure 7). Sequence of Strain S was also submitted to GenBank under the accession number FJ416862. Antimicrobial susceptibility testing. Zone of inhibition present around the antibiotic disks was measured and compared with the standard provided in the antibiotic kits by Himedia to determine whether the organism was resistant, intermediate or sensitive (Table 2). Strain P isolated from sputum, which showed morphological, biochemical and genetic characteristics of S. aureus was sensitive to many commonly used antibiotics, which is quite contradictory to the literature data [18, 20, 21], where S. aureus has been reported to accumulate resistant markers for several classes of antibiotics. The antibiotic profile of strain A isolated from urine reveals that the strain might be pathogenic as it is intermediately resistant and also completely resistant to many antimicrobial agents, while S showed a pattern similar to P. All three strains are vancomycin resistant which confirms recently reported facts. Table 2.Antimicrobial sensitivity assay of three Staphylococcus strains.
Legend: Antibiotics – Ampicillin (A), Cephadroxil (Cq), Chloramphenicol (C), Cloxacillin (Cx), Cephotaxime (Ce), Ceftazidime (Ca), Ciprofloxacin (Cf), Doxycycline Hydrochloride (Do), Gentamicin (G), Metronidazole (Mt), Neomycin (N), Norfloxacin (Nx), Polymyxin B (Pb), Rifampicin (R), Roxithromycin (Ro), Tetracycline (T), Trimethoprin (Tr), Vancomycin (Va). Sensibility – sensitive (S), intermediate resistant (I), resistant (R). Conclusion This study dealt with the isolation and characterization of staphylococcal strains from varied biological origin and their comparison with the standard laboratory strain. From the above study it was clear that chromogenic media available are not as specific as they are claimed to be. They could be relied on only up to genus level. Molecular phylogeny along with biochemical analysis in this case helps in precise identification of the microbe. This study reflects the loss of lecithinase production and thus pathogenicity with subsequent rounds of cultivation outside the body under laboratory conditions for isolates A and P. The same explanation also holds true for the complete lack of activity in case of isolate S. The partial 16S rRNA based molecular analysis showed 100 % similarity of isolates P and S with S. aureus. The difference at the biochemical level (protease, lipase, DNase) with the reported S. aureus, and also among themselves in respect to growth curve as well as antibiotic response clearly indicated that partial 16S rRNA based phylogenetic analysis using neighbour joining method was not the accurate approach for understanding the molecular identity at species level but could reveal the identity only at genus level. This discrepancy may be due to the fact that the region sequenced in this case (about 400 base pairs) did not contain the variable portions of the gene. So, a complete 16S rRNA gene sequence based phylogenetic analysis provides the more precise molecular identity of the isolates and determines strain S as S. epidermidis. This study also reports a novel strain of S. haemolyticus (isolate A) and S. epidermidis (Isolate S) with the GenBank accession numbers EF644492 and FJ416862 respectively. Acknowledgement. The authors would like to acknowledge the financial support of the Department of Science and Technology, Government of India as well as the Department of Biotechnology, Government of India. They would like to thank West Bengal University of Technology for facilitating the computational support provided by Department of Biotechnology, Government of India. The authors would also like to acknowledge NICED Culture Collection Center for gifting us the Staphylococcus strain. References
Copyright 2009 - National Bank for Industrial Microorganisms and Cell Cultures - Bulgaria The following images related to this document are available:Photo images[cc09007f7.jpg] [cc09007f2.jpg] [cc09007f5.jpg] [cc09007f1.jpg] [cc09007f3.jpg] [cc09007f4.jpg] [cc09007f6.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}