 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Journal of Culture Collections, Vol. 6, No. 1, 2009, pp. 69-75 Identification of halophilic bacteria from fish sauce (Nam-pla) in Thailand Somboon Tanasupawat1*, Sirilak Namwong1, Takuji Kudo2 and Takashi Itoh2 1Department
of Microbiology, Faculty of Pharmaceutical Sciences, Chulalongkorn University,
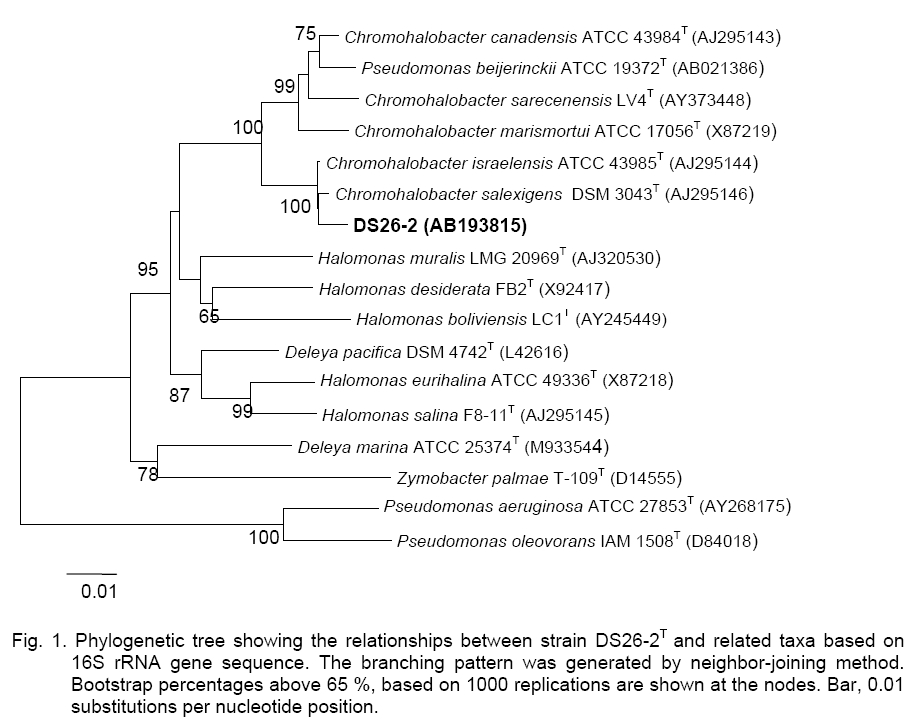
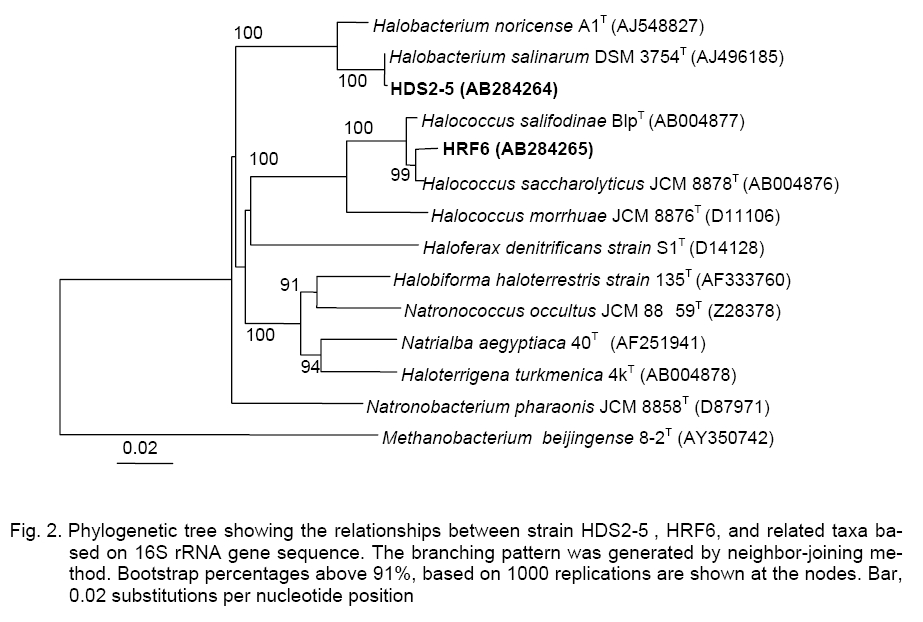
Bangkok 10330, Thailand; Code Number: cc09009 Summary Four strains of Gram-negative, rod-shaped, moderately halophilic bacteria, Group A, and eleven strains of strictly aerobic, extremely halophilic rods (10 strains, Group B) and coccoid (1 strain, Group C) were isolated from fish sauce fermentation (nampla) in Thailand. The 16S rRNA gene sequence analyses of the representative strains indicated that DS26-2 (Group A), HDS2-5 (Group B), and HRF6 (Group C), were closely related to Chromohalobacter salexigens KCTC 12941T, Halobacteirum salinarum JCM 8978T, and Halococcus saccharolyticus JCM 8878T with 99.3, 99.9, and 99.0 % similarity, respectively. Group A strains were identified as C. salexigens, Group B as H. salinarum, and Group C strain was H. saccharolyticus based on their DNA-DNA relatedness. Group A strains grew in 3–25 % (w/v) NaCl. Ubiquinone with nine isoprene units (Q-9) was a major component. The DNA G+C contents ranged from 63.1 to 64.2 mol %. Group B and Group C strains grew optimally in the presence of 25-30 % NaCl. The tested strains of Group B contained major menaquinone with eight isoprene units (MK-8). DNA G+C contents ranged from 63.3 to 64.7 mol %. Group C strain had MK-8(H2) as a predominant menaquinone. The DNA G+C content was 63.2 mol %. Key words: Archaea, Chromohalobacter salexigens, fish sauce, halophilic bacteria, Halobacteriumsalinarum, Halococcussaccharolyticus Introduction The moderately and extremely halophilic bacteria occur to thrive in the salted environment, i.e. in salt lakes, soda lakes, saltern, crude solar salts and proteinaceous productswith the capacity to balance the osmotic pressure of the environment and resist the denaturing effects of salts [5, 8, 14]. The moderately halophilic bacteria i.e., Lentibacillus, Filobacillus, Tetragenococcus, Chromohalobacter are able to grow over a wide range of salt concentrations and grow optimally in media containing between 3 and 15 % (w/v) NaCl [1, 2, 19]. On the other hand, the halophilic archaea i.e., Halococcus and Halobacterium require at least 1.5 M NaCl (optimal growth at 20–25 %, w/v NaCl) for growth [11, 39]. In Thailand, Fish sauce contains high concentration of salt (25–30 %, w/v NaCl), thus microorganisms found during fish sauce production are generally classified as halophilic bacteria [16].The strains of Lentibacillus salicampi, L. jurispiscarius [19], L. halophilus [30], Filobacillus sp. RF2-5 [10], Halobacillus sp. SR5-3 [20], Piscibacillus salipiscarius[31], Tetragenococcus halophilus and T. muriaticus [34], Halobacterium salinarum [35],and Halococcus thailandensis [21] were isolated from fermented fish sauce and related fish products. This work deals with the identification of the moderately and extremely halophilic bacteria isolated from fish sauce based on their phenotypic and chemotaxonomic characteristics including DNA-DNA relatedness and phylogenetic properties. Materials and Methods Source of samples and Isolation. Fish sauce samples (nampla) were collected from the factories (Sindhu Samuth Fish Sauce Factory (Squid Brand) LTD. (A), Samutprakarn, Thai Fish Sauce Factory (Squid Brand) Co., LTD. (B), Samutsongkram, and Rayong Fish Sauce Industry Co., Ltd. (C), Rayong Province, in Thailand. The halophilic bacteria were isolated by using spread-plate technique on agar plates of JCM medium No. 169 and incubated at 37 °C for 1-2 weeks. Identification methods. Morphological, cultural, physiological and biochemical characteristics were determined as described by Barrow and Feltham [3] and Namwong et al. [19, 21]; Oren et al. [22]. Acid production from carbohydrate was determined in the medium described by Leifson [15] supplemented with 6.5 % for the moderately and 16.5 % NaCl for the extremely strains. Determination of the antibiotic susceptibility of halophilic archaea was tested as described by Stan-Lotter et al. [27]. The meso-diaminopimelic acid in the peptidoglycan and menaquinones were analyzed as described previously [12]. Polar lipids were determined according to the method of Minnikin [17]. The quantitative analysis of cellular fatty acids was employed with the Microbial Identification System (MIDI Inc.) [25]. DNAs were isolated and purified according to the method of Saito and Miura [23]. The DNA G+C content was determined by the method of Tamaoka and Komagata [28] using reversed-phase HPLC. DNA-DNA hybridization was conducted as described by Ezaki et al. [6] and detected as reported by Tanasupawat et. al. [29].The 16S rRNA gene sequence of the moderately isolate was determined as described previously [26], whereas the archaea strains were determined as described by Namwong et al. [21]. The phylogenetic tree based on 16S rRNA gene sequences was constructed as described by Kumar et al. [13]; Felsenstein [7]; Saitou and Nei [24]; Thompson et al. [33]. The GenBank/ EMBL/DDBJ accession numbers for the 16S rRNA gene sequences of strain DS26-2, HDS2-5, and HRF6 were AB193815, AB284264 and AB284265, respectively. Results and Discussion A total of fifteen isolates were divided into three groups, the moderately rod-shaped halophilic bacteria, Group A (4 isolates) and the extremely rod-shaped halophilic bacteria, Group B (10 isolates) and a sphere-shaped bacterium, Group C (1 isolate, HRF6) based on their phenotypic and chemotaxonomic characteristics, DNA-DNA relatedness including 16S rRNA gene sequence analyses (Figure 1, Figure 2; Tables 1, 2). Group
A contained isolates KS11-1, DS26-2, KS87-5, and PB12.
All were Gram-negative, cream-pigmented, non-spore-forming rods. Cells measured
0.7-1.0 µm in width and 2.0- 5.0 µm in length and occurred singly or in pairs
on JCM. No. 169 agar plates supplemented with 10 % NaCl. Catalase,
oxidase, urease, and nitrate reduction were positive but indole formation was
negative. Group A isolates hydrolysed arginine and starch but did not hydrolyse
gelatin, casein, Tween 80 and tyrosine. They were capable to grow at
temperature ranging from 10 to Table 1. Differential characteristics of strains in Chromohalobacter,Halobacterium and Halococcus.
*Strains: 1, 4 strains in
Group A; JCM, Japan Collection of Microorganisms, RIKEN BioResource Center, Saitama, Japan; KCTC, Korean Collection for Type Cultures, Biological Resource Center, Korea Research Institute of Bioscience and Biotechnology, Daejeon, Korea. Group B contained isolates HDB1-1, HDB1-11, HDB1-31, HDS2-5, HDB10-5, HIB20-2, HIB60-1, HIS10-4, HIS30-1, and HIS50-2(1). All were Gram-negative, red-pigmented, non-spore-forming rod-shaped archaea. Cells measured 0.7-1.0 µm in width and 2.0-3.0 µm in length and occurred singly or in pairs. Colonies were circular and convex on JCM. No. 169 agar plates. Group C, HRF6 was red-pigmented and sphere-shaped. The cells measured 0.8-1.0
µm in size and appeared singly or in tetrads. The isolates in both two Groups
grew optimally in the presence of 20–30 % (w/v) NaCl. Growth did not occur
in the presence of less than 15 % NaCl. Cells of Group B strains lysed in
distilled water, but Group C strain did not. They grew optimally at Group B strains proved to be positive for catalase and oxidase. Some strains hydrolysed casein and gelatin but did not hydrolyse arginine and tween 80. Growth on single carbon sources was not observed. No acids were produced from L-arabinose, D-cellobiose, D-glucose, lactose, D-mannitol, D-melibiose, raffinose, sucrose, and D-trehalose (Table 1). The strains were susceptible to bacitracin (10 units), novobiocin (5 µg) and rifampicin (30 µg) but resistant to ampicillin (10 µg), chloramphenicol (30 µg), gentamicin (10 µg), kanamycin (30 µg), nalidixic acid (30 µg), tetracycline (30 µg), and streptomycin (10 µg). The representative of Group B strain, HDS2-5 contained menaquinones of MK-8 (95.8 %) and MK-8 (H2) (4.2 %). The major polar lipids presented were phytanyl-sesterterpanyl moieties (C20C25) of phosphatidylglycerol (PG), phosphatidylglycerol phosphate (PGP) and phosphoglycerol sulfate (PGS). Glycolipids were S-TGD-1, and S-TeGD. Group B strains contained two polar lipids, the glycerol diether analogs (C20C25 and C20C20) of PG and PGP. On the basis of 16S rRNA gene sequence analyses, the representative strains of Group B, HDS2-5 and Group C, HRF6, were represented the genera Halobacterium and Halococcus, respectively (Figure 2). HDS2-5 was closely related to the H. salinarum JCM 8978T with 99.9 % similarity whereas HRF6 corresponded to Halococcus saccharolyticus JCM 8878Twith 99.3 % similarity. The DNA G+C contents of tested strains in Group B ranged from 63.3 to 64.7 mol % and HRF6 was 63.2 mol %, respectively (Table 2). Table 2. DNA G+C contents and DNA-DNA relatedness of strains in Chromohalobacter, Halobacterium and Halococcus.
ND, Not
determined. Group B strains were closely related among themselves with 90-102.9 % of DNA-DNA relatedness, suggesting that they belong to the sa-me species. On the other hand, all strains showed the high levels of DNA-DNA relatedness to H. salinarum JCM 8978T (87.3 - 101.1 %) as shown in Table 2. The representative strain, HDS2-5 contained C20C25 derivative of several kinds of polar lipid such as glycolipids (S-TGD-1 and S-TEGD) that are characteristics and diagnostic for the genus Halobacterium [8]. There-fore, Group B strains were identified as Halobacterium salinarum [38]. HRF6 contained two major polar lipids, the glycerol diether analogs (2-O-sesterterpanyl-3-O-phytanyl-sn-glicerol, C20C25 and 2, 3-di-O-phytanyl-sn-glycerol, C20C20) of phosphatidylglycerol (PG), phosphatidylglycerol-methylphosphate (PGP-Me) and MK-8(H2) (89.4 %), and MK-8 (10.6 %) that were different from the strain of Group B [8]. HRF6 exhibited high level of DNA-DNA relatedness (88.5 %) with H. saccharolyticus JCM 8878T buthad low level of DNA-DNA relatedness (8.3 %) with H. morrhuae JCM 8876T. Therefore, a sphere-shaped HRF6 (Group C) was identified as H. saccharolyticus [18, 27]. In fish sauce and related products, Lentibacillus jurispiscarius, Filobacillus sp., Halobacillus sp., T. halophilus, T. muriaticus, H. Salinarum, H. piscisalsi, and Natrinema gari [10, 19, 20, 29, 32, 34, 35, 39] were reported. In this study, the moderately halophilic bacteria, C. salexigens and the extremely halophilic archaea, H. salinarum and H. saccharolyticus strains were commonly distributed and played some roles in many stages of fish sauce fermentation. Acknowledgements. We are very grateful to Sindhu Samuth Fish Sauce Factory (Squid Brand) LTD., Samutprakarn, Thai Fish Sauce Factory (Squid Brand) Co., LTD., Samutsongkram and Rayong Fish Sauce Industry Co., Ltd., Rayong Province, for providing the samples. This study was in part supported by the Thai-land Research Fund for a 2001 Royal Golden Jubilee Scholarship as a research grant to S. N. and by Ratchadapiseksomphot Endowment Fund, Chulalongkorn University (2002). References
Copyright 2009 - National Bank for Industrial Microorganisms and Cell Cultures - Bulgaria The following images related to this document are available:Photo images[cc09009f1.jpg] [cc09009f2.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}