 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Journal of Culture Collections, Vol. 6, No. 1, 2009, pp. 85-96 Asporogenic mutants of Alternaria cassiae generated by x-ray irradiation Olubukola O. Babalola1 Department of Plant Sciences, Weizmann Institute of Science, Rehovot
IL-76100, Israel Code Number: cc09011 Summary The prevention of
sporulation is an important factor in precluding the spread of transgenic
biocontrol agents beyond desired limits; asporogenic deletion mutants could
prevent such spread. Alternaria
cassiae is a biological control agent for controlling sicklepod (Cassia
obtusifolia) and coffee senna (Cassia occidentalis). The NEP1
transformed fungus is more virulent than the parent. A. cassiae forms microcolonies on 0.9 g/l sodium
deoxycholate plates. The spores have a D10 value of 50,000 rads when
subjected to X-ray beams. Five putative mutants were achieved and 3 proved to
revert. Mutants xAlt541 and xAlt524 are stable with no spore-forming
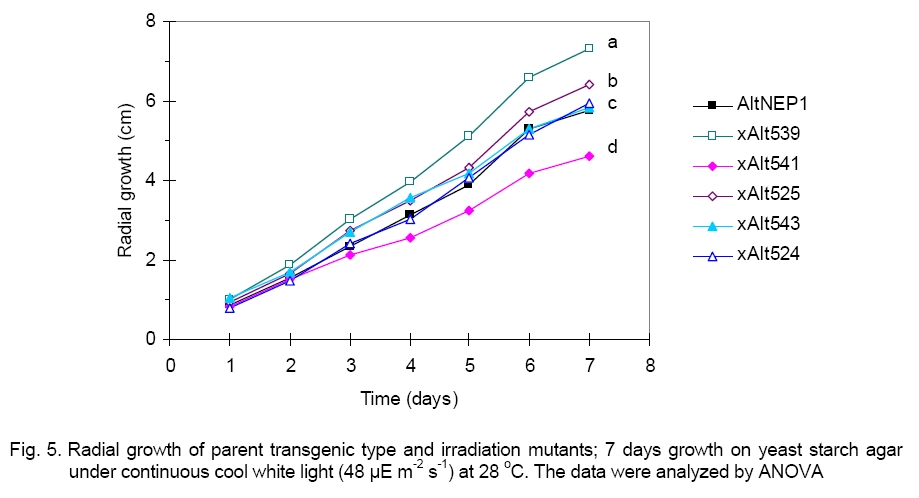
capability. The xAlt541 mutant grew more slowly ( Keywords: mycelia, NEP1, spore Introduction The prevention of sporulation is the critical factor in preventing the spread of transgenic biocontrol agents beyond desired limits. Fungal mycelia have proven to be better inoculating agents than spores with fungi controlling Orobanche and Abutilon [2, 9], thus spores are not needed in these cases. Spores are dispensable structures, inessential for normal growth, and indeed, inessential for storage and propagation since the sexual cycle provides an alternative source of spores [8]. Therefore, the aim of these studies is to use irradiation to generate asporogenic mutants of Alternaria cassiae. A. cassiae spores are large, darkly pigmented, and surrounded by a thick, two-layered wall [24, 40]. Each spore is divided by transverse and longitudinal septa into multiple cells, with size ranging from 20 to 25 µm in width [24] and from 75 to 200 µm in length, with long filiform beaks [40], which do not exceed one-third of the spore length. A. cassiae spores have multiple nuclei each with prominent nucleolus [24]. In culture on YSA media, colonies of A. cassiae appear slate gray, and are cottony in texture. Different colors are produced on different media. A. cassiae Jurair and Khan, an important filamentous fungal phytopathogen of economic importance in weed biological control, is used as a test fungus. A. cassiae is a biological control agent for sicklepod (Cassia senna L.) [14, 24, 29] and coffee senna (Cassia occidentalis L. # CASOC). It had also been reported on cowpea (Vigna unguiculata) [25]. Sicklepod is also a major biotic constraint to production of soybean (Glycine max) [41], peanut (Arachis hypogaea), and cotton (Gossypium hirsutum) [11]. Many of the early investigations were focused on the potential of mycoherbicides on target weeds. Now, there is a need to develop asporogenic mutant strains as researchers have identified that the mycoherbicides must be restricted to the target weed [9]. Safety concerns raised include the persistence of the fungi in the environment and the possibility of the transfer of a hypervirulent transgene from the biocontrol agent to a pathogen of a crop. Failsafe mechanisms proposed to prevent spread and mitigate introgression of transgenic hypervirulent biocontrol fungi include asporogenic deletion mutants of the transgenic hypervirulent fungi, which would prevent the spread of the biocontrol agent and environmental persistence [13, 32, 33]. The important characteristic of ionizing radiations is the localized release of large amounts of energy [15]. The energy dissipated per ionizing event is about 33 eV, which is more than enough to break a strong chemical bond; for example, the energy associated with a C=C bond is 4.9 eV [15]. Spores, like all living matter, are sensitive to the actions of radiation and though spores are more resistant to stress conditions than vegetative cells, no spore exists that cannot be killed by sufficient dosage. On the other hand, hyphae also are sensitive to the actions of ionizing radiation. Consequently, the idea is to make the fungi asporogenic with little damage to the hyphae. To this end, dose levels were identified which involve various levels of damage to A. cassiae. The intent of treatment is stable control of sporulation. There is a dearth of information about the effect of radiation on fungi. Interest in this area is in connection with irradiation of fungi as it is necessary as a failsafe technique against off-target spread of transgenic fungi. The objectives of the research reported in this study were: (1) to maximize spore production on chemically defined media in preparation for ionizing radiation of the fungal spores and for use in assessing the competence of irradiated spores to germinate and produce spores; (2) to determine the relationship between radiation and survival; and (3) to generate asporogenic fungi mutants which can tolerate normal growth conditions. Materials and Methods Cultivation media, fungi and culture maintenance. Media tested (see Table 1) were used alone and augmented with 1 g/l of either L-phenylalanine or L-tyrosine. The amino acids were added to media as melanin precursors in the hope that they would make it easy to identify asporogenic mutants. The spores of the test fungus, NEP1 transformed A. cassiae (hereafter refer to as AltNEP1), were cultivated at 28 °C under continuous cool white light conditions for 10-14 days on yeast-starch-agar (YSA). A. cassiae was grown on potato dextrose agar (PDA) slants at 28 °C for 14 days and maintained at 4 °C. Spore formation in different culture media and
spore quantification. The agar medium was centrally inoculated with a small plug
( Spores were dislodged from the mycelium of 10-14 day-old cultures in 2 ml sterile distilled water using a rubber policeman. All possible conidial residuals from the plates were rinsed three times with 2 ml sterile distilled water. The rinsing was added to the initial tube, and the final spore suspension was filtered through Nitex® Bolting Cloth (70 µm). Spores were counted on a grid from the surface of a hemîcytometer mounted on a light microscope. Spores per ml of original sample = spore count/dilution factor. Irradiation of spores and dose determination. Five
ml of the spore suspension (103 spores/ml of phosphate buffer, Assay for asporogenic mutants. Batches
of fifty 24-well cluster dishes were prepared with YSA. Single colonies of
putative mutants (visibly different from normal) were subcultured into each well
of the cluster dishes from Petri plates. The single colonies were transferred
to continuous cool white light at 28 °C for 10 days. The
cultures were screened by checking under the inverted microscope with
phase-contrast or under the light microscope for asporogenic mutants.
Asporogenic mutants were picked and subcultured on Observations for mutant strain stability and
mutant characterization. Suspensions from parent strains and irradiated mutant strains were examined
under the microscope. The cultures isolated as asporogenic were finally classified
as asporogenic mutants after 5-7 successive ‘‘generations’’. Each transfer
involved 10 days-culture on agar plates. The hyphae morphology was photographed
4 days after inoculation. Mutants were observed for morphology, mycelia
biomass, pigment formation, and daily mycelia extension. Agar plates were inoculated
centrally with one 5-mm diameter mycelial mat taken from the periphery of
actively growing agar culture by means of a sterile cork borer. The dishes were
incubated at 28 °C. For dry weight determination, five pieces
of mycelial mat cultures Molecular analysis of mutants. The mycelia from asporogenic mutants were recovered from 48 h shake
culture in potato glucose broth (PDB) (Pronadisa) (at 28 °C). The
mycelia were collected and ground in liquid nitrogen with a mortar and pestle.
Genomic DNA from mycelia was obtained as stated in Daboussi et al. [10]. To
confirm that the mutants still carry NEP1, Polymerase Chain Reaction (PCR)
detection of NEP1 was carried out. The primers designed to amplify NEP1 were a
forward primer 5’-CCG-ACGGTTGTCAGCCATACAC- Pathogenicity assay. Pure mycelial mats of A. cassiae
( Greenhouse grown Cassia
senna L. plants in 8.5 X 8.5 X For statistical analyses, the damage was rated visually for disease progression at 2 and 5 d post-inoculation using a 0-3 scale [2]: 0 = no infection or a hypersensitive reaction; 1 = moderate infection, plants still alive; 2 = severe infection, plants hardly alive, but one leaf may be alive and the plants can recover; 3 = complete desiccation and abscission of all leaves, dead plants. The results are shown as means with standard errors, and were further statistically analyzed as described below. To retain pathogenicity, the fungi were reisolated from the sprayed plants 2 days after inoculation. Statistical analysis. Fungal characterizations were carried out twice under identical conditions, each with three replica platings. Data generated are reproducible. Means ± standard error are indicated in the text. Analysis of variance (ANOVA) for the ratios obtained and posthoc tests were performed by using SAS version 9.1 [34]. Fisher’s protected least significance test (LSD) test was used to determine the significance of pairwise comparisons between treatments. Statistical significances were judged at the 5 % level. Damage severity from mutants was compared with the parent type by Kruskall-Wallis ANOVA test. Results and discussion Cultivation media, spore germination, and quantification. The influence of medium composition was tested on fungal sporulation (Table 1). The aim was to optimize spore production on chemically defined media in preparation for ionizing radiation of the fungal spores. The sporulation ability of AltNEP1 was studied and compared on agar media. The results showed that the highest number of spores were for-med (1.7 to 4.7 x 10-3 spores/ml) on YSA (Figure 1, Table 1). However, in PDA, a predominantly vegetative mycelium was formed (Figure 1). A. cassiae thrived on PDA. PDA, however, encourages the growth of mycelium rather than spore formation. The fungi were maintained by serial transfer from stale to fresh, solid media. Table 1. Effect of various cultivation media (broth and agar) and the influence of two amino acids on A. cassiae culture pigmentation and sporulation.
The aim of screening different media was to identify the media which could induce the highest spore production. YSA showed the presence of spores for A. cassiae. That was highly desired in the process to check for asporogenic mutants and, when possible, the likely revertants. YSA is a nutrient media, yet it was found effective for the study purpose. A previous study [39] observed a slight increase in sporulation of some fungi strains with yeast extract which was attributed to the vitamins in yeast extract. Yeast dextrose agar is recommended for fungal sporulation [22]. It is known that cultural conditions determine the type of pigment formed [39]. Tyrosine was added to the agar medium. Tyrosinase is the rate-limiting enzyme in melanin synthesis [12] and melanin is formed from tyrosine. Formation of microcolonies.To permit the use of higher plating densities (about 50 colonies/plate) than is normally possible when single colonies are desired, 0.9 g/l sodium deoxycholate was added to the agar medium before plating. A. cassiae formed microcolonies on 0.9 g/l sodium deoxycholate plates. The colony forming unit per plate was adjusted to get a countable number of colonies. Dose-response curve. To assess the optimum dose of X-ray irradiation for total asporogenesis of A. cassiae, X-ray doses were sequentially increased and many irradiation doses were investigated (25,000 to 100,000 rads). A. cassiae curve demonstrates clearly that the X-ray survival curve has an initial ‘‘shoulder’’. The initial shoulder region may be attributed to the multinuclei nature of the A. cassiae spores. The existence of a shoulder on the survival curve of cells has been interpreted to imply a multi-target, multi-hit effect of radiation [7]. According to target theory, the mathematical approach to the interpretation of irradiation survival curves [16], survival curves with ‘‘shoulders’’, as shown by A. cassiae (Figure 2), indicate that the killing of certain cells requires one or more hits on each of several critical targets. Such cells are capable of accumulating a certain ‘‘threshold dose’’ before an effect is observed [19]. This was referred to as an ‘‘induction’’ dose (Di), which is defined as ‘‘the point of intersection between lines parallel with the x-axis and the extrapolation line of the linear portion of the survival curve to the y-axis’’ [26]. Some plates with X-ray irradiated spores gave higher propagule counts than the controls. The dose-response curve of A. cassiae to X- rays varies critically with a changing dose rate and it is hard to say conclusively at which dose fungal cultures remain viable but with no spore-forming capability. Energy from X-rays is deposited unevenly in ‘‘spurs’’ and ‘‘blobs’’. This may lead to sites with multiple damages, that is, a combination of a double-stranded break and a base damage. The purpose of these findings is to determine the D10 radiation dose of A. cassiae spores. The spores were irradiated with 25,000 to 100,000 rads units and the irradiated cultures were plated. The log of the number of survivors against the dose for getting the survivor curve was plotted. The spore has a D10 value of 50,000 rads. The sensitivity of A. cassiae to radiation ranges from ≥ 25,000 to 100,000 rads (Figure 2). It seems clear that these spores are quite resistant to X-radiation, since more than half of them survived a dose of 60,000 rads (Figure 2). The present data show the potential of A. cassiae’s resistance to irradiation, the destruction of which follows a statistical pattern. Alternaria has been reported to show resistance to irradiation [31]. The resistance of A. cassiae to irradiation is a result of its pigmentation. A. cassiae is dematiaceous - a fungus having brown or black melanotic pigment in the cell wall. The decrease in the sensitivity of the A. nidulans mutant to UV correlates with the accumulation of a red pigment by the strain, that is, the pigment makes the spores less sensitive to irradiation [18]. Assay of asporogenic mutants. A number of colonies were examined and about 4,000 colonies have been screened. The five mutants isolated have been given the prefix xAlt, for X-ray derived asporogenic A. cassiae. The mutants are xAlt539, xAlt541, xAlt525, xAlt543, and xAlt524. Given that an average
of one asporogenic mutant is expected per ten thousand colonies without
irradiation, a mutation rate of one in a million is expected. With irradiation
at D10 dose, one mutation is expected from 100,000. If there are 10
possible mutation sites for sporulation one in 10,000 is expected. The current
study has concentrated on the mutant derived from A. cassiae. Earlier work on sporulation mutation showed that many
genes control various steps in the sporulation pathways [37, 38]. The latter
could be one of the reasons why the mutants are relatively difficult to get. Mutation
rates per locus per rad obtained from experimental organisms extend over an
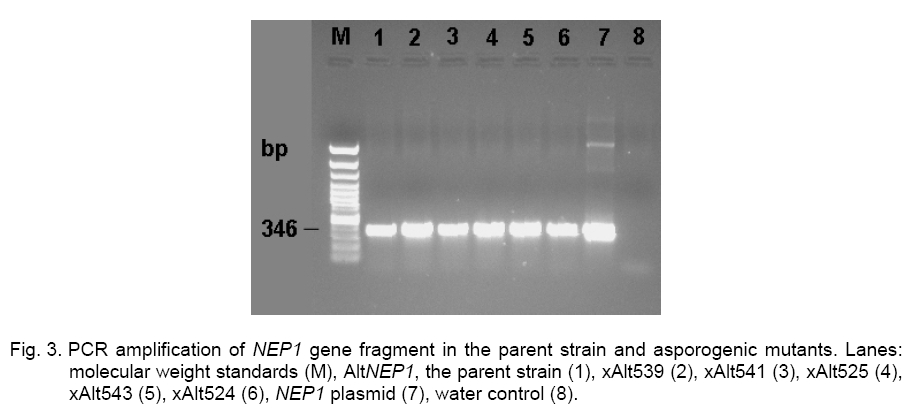
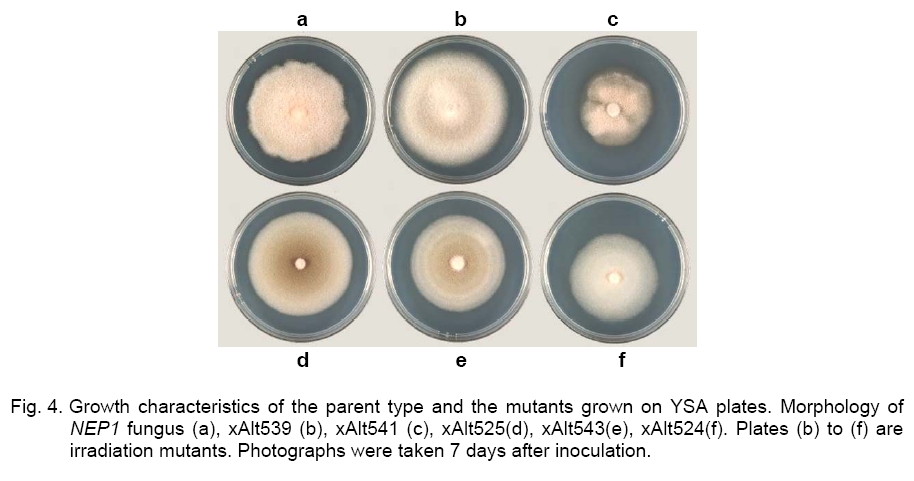
enormous range of three orders of magnitude, being 1 x 10- Observations for mutant strain stability. Three of the cultures, xAlt539, xAlt525, and xAlt543, obtained by X-ray experiments and recorded as mutants, reverted to normal after the third transfer. About half the mutants obtained by X-ray experiments reverted to normal after one or several subcultures [21]. Many single-strand breaks are produced in DNA by radiation but are readily repaired using the opposite DNA strand as a template. Spontaneous morphological changes in filamentous fungi have been reported in phytopathogenic filamentous fungi [17], with a similar correlation between morphological changes. The ability to sporulate is nuclearly controlled but not cytoplasmic. In addition, sporulation is conditioned by at least several nuclearly inherited functions [42]. Several unlinked sporulation gene clusters seem likely to exist [35], so if one of the sporulation genes is unavailable, the presence of other sporulation genes may take over the function of the lost gene. When DNA is irradiated with ionizing radiations, irreversible damage occurs only when a very large number of hydrogen bonds are all broken at the same time [4]. This means there is likely to have been a main-chain scission (a break in each of the adjacent chains less than about 5 nucleotide units apart) caused by the radiation in xAlt541 and xAlt524. The mutants that reverted (xAlt539, xAlt525 and xAlt543) might have had only crosslinks in the DNA molecule. Molecular analysis. Molecular analysis was performed to check whether the NEP1 fragment was not disrupted by radiation treatment. The NEP1 fragment (346-bp) in each of the mutants was intact (Figure 3). The results suggested that the mutation does not occur in loci involving the NEP1 fragment or there was radiation-inducible DNA repair in the fragment. This means that the three- to four-fold reduction of transformation frequency below the spontaneous level caused by low dose irradiation [36] is overcome as the added NEP1 fragment could still be expressed. Mutant characterization. To reveal possible variants in morphology due to radiation treatments, the mutants were characterized. The asporogenic mutants xAlt543, xAlt524, xAlt525, xAlt539, and xAlt541, were isolated from survivors of NEP1 A. cassiae after X-ray irradiation. The colony morphology of NEP1 A. cassiae and those of the mutants are shown in Figure 4. The irradiation leads to some different outgrowth patterns observed in visible colony diameter, mycelia biomass, pigment formation, growth morphology, and the daily mycelia extension compared with the NEP1 A. cassiae control. Mutant xAlt524 seems to assume the form of a dimorphic fungus as a result of the irradiation treatment. Resulting mutants xAlt539, xAlt541, xAlt525, xAlt543, and xAlt524, which produce no spores, were checked for morphological variants (Figure 4). Diversity of colonial forms, ranging from tiny to vigorous forms, had been reported in Neurospora crassa[23]. Mutant xAlt524 lacks aerial hyphae, pigmentation, and spore production. When xAlt524 was subcultured, it grew no further and could not give rise to a colony. In liquid culture, the mutant was completely autolyzed by 48 h. Mutant xAlt524 displays morphological abnormalities and cell lysis. Accordingly, colonial growth of xAlt524 was scanty. The xAlt524 mutant is assumed to be producing lytic enzymes for self-degradation. The autolysis of filamentous fungi was observed in [27, 30]. The irradiation leads to some different outgrowth patterns because of mutations of the fungal colonies surviving the irradiation treatment. X-ray treatment induces both gene mutations and chromosomal aberrations, including inversions [23]. The NEP1 type produced a rosy brown to peach puff pigment after 5 days of growth on YSA plus tyrosine (Figure 4). The xAlt524 mutant produced a cream and xAlt541 produced dark salmon color. Pigmentations of xAlt543 and xAlt339 were the same, having a Navajo white, dark golden rod, or light salmon mycelial color (Figure 4). A distinctive feature of a Magnaporthe grisea sporulation mutant (abnormal conidium morphology and a reduction in sporulation) was delayed and reduced pigment formation [38]. The xAlt524 mutant lost its pigmentation and so can be readily lysed. Melanized appressoria are resistant to lysis by antagonistic bacteria [20]. Except for xAlt524, all the other mutants retained fluffy mycelia. AltNEP1 has abundant mycelia which were still retained by xAlt539 and xAlt541. The cultures of xAlt543 had moderate but non abundant mycelial density. Mutants xAlt524 and xAlt525 had scanty mycelia (Figure 4). Comparison of
phenotypes of the asporogenic mutants xAlt539, xAlt541, xAlt525, xAlt543,
xAlt524, and the parent AltNEP1
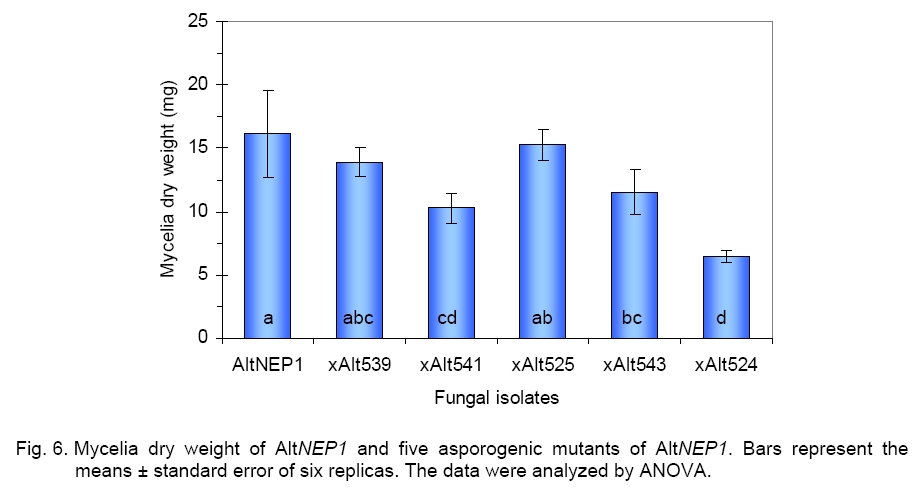
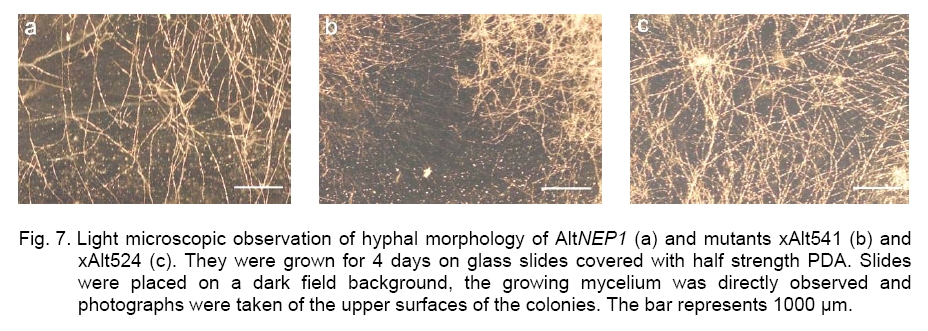
revealed that xAlt541 grew significantly more slowly ( For mycelia dry weight determination, data from six experiments showed that the parent AltNEP1 developed more mycelium than all the mutants (Figure 6). There was a striking significant difference between dry weight of xAlt524 (6.46 mg) and the parent type fungus (16.13 mg). This reduction led to approximately only 40% growth. The other mutants also showed reduced dry weights, respectively xAlt539 (86.29%), xAlt541 (63.55%), xAlt525 (94.48%), and xAlt543 (71.6%). There was no significant difference between AltNEP1 and twoof the mutants (xAlt525 and xAlt539). The hyphal growth and branching of the parent fungus AltNEP1 and the stable mutants xAlt541and xAlt524 are depicted in Figure 7 (a, b and c, respectively). If the fungal colonization ability of the fungus (hyphal growth and branching) is reduced, then the pathogenicity may be reduced. Virulence test. Pathogenicity of the mutants was examined to determine whether the mutants were still virulent despite the mutation effect of irradiation on their sporulation capability. All the mutants caused wilting and necrosis with xAlt525 being statistically (2.67ab ± 0.1) insignificantly different in vigor than the parent NEP1 culture (Figure 8, top panel). To see clearly the infection vigor of the mutants on older plants, xAlt541 and xAlt524 (stable mutants) were sprayed on 2-leaf stage cassia. AltNEP1 (1.5a ± 0.22), xAlt541 (1.33ab ± 0.21), and xAlt524 (1.0b ± 0.00) damage severity was statistically different by two days post-inoculation. However, damage severity from AltNEP1 (2.5a ± 0.22), xAlt541 (2.17a ± 0.31), and xAlt524 (2.0a ± 0.26) was not statistically different by 5 days post-inoculation (Figure 8, bottom panel). The mutants had the capacity to cause disease on cassia plants and the levels of damage severity equaled that of the parent strain over time. Either the sporulation of A. cassiae is not related solely to fungal virulence or the irradiation treatment had not affected some of the components vital for colonization and hence virulence. The present results are in agreement with that of [5] who reported that X-ray sensitive mutants are similar, if not identical, to their wild-type counterparts in other biological functions. Asporogenic fungi could mitigate risks of transgene introgression into related fungi. The mutants have the NEP1 gene and could also still express the gene on target plants. Melanin biosynthesis is an important determinant of pathogenicity and virulence [6, 28]. However, not all pathogenic fungi are melanotic, as seen with xAlt524. Even on YSA plus tyrosine xAlt524 does not form a pigment. Although X-ray irradiation has an effect on fungi mutagenesis, some of the mutants revert and could not be differentiated from the parent isolate. By 5 days post-inoculation, the fungal virulence of the mutants had no statistically significant difference from that of the parent isolate. It is possible that there could be stabilization of the mutated gene in the revertants if they were again irradiated, but this hypothesis requires further research. Results from this study have made it clear that asporogenic mutants could reduce the ecological constraint of the use of mycoherbicides. The mutants could be mass-produced and stored for reasonable periods e.g. > 9 months, [3] prior to field application. Acknowledgements. I thank Inbar Greenspoon for her laboratory assistance, Drs Sagit Meir and Yoav Herschkovit for their research comments and Prof. Jonathan Gressel for human resources. GB Moshe provided helpful advice and guidance in the use of irradiation machines. This research was performed in the Department of Plant Sciences, Weizmann Institute of Science, Rehovot, Israel. This study was supported by International Atomic Energy Agency, Vienna. References
Copyright 2009 - National Bank for Industrial Microorganisms and Cell Cultures - Bulgaria The following images related to this document are available:Photo images[cc09011f7.jpg] [cc09011f2.jpg] [cc09011f5.jpg] [cc09011f6.jpg] [cc09011f1.jpg] [cc09011f3.jpg] [cc09011f8.jpg] [cc09011f4.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}