 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Journal of Culture Collections, Vol. 6, No. 1, 2009, pp. 106-111 Serological characterization of paprika mild mottle tobamovirus strain P101 Angela Yordanova1* and Elisaveta Stoimenova2 1National Bank for Industrial Microorganisms and Cell Cultures, P.O.Box
239, 1113 Sofia, Bulgaria; Code Number: cc09013 Summary The serological properties of the Bulgarian strain P101 of paprika mild mottle virus (PAMMV) were investigated. Counter immunoelectrophoresis and double diffusion test were applied. The serological relationship with tobacco mosaic virus (TMV), tomato mosaic virus (ToMV), tobacco mild green mosaic virus (TMGMV), pepper mild mottle virus (PMMoV) and the Dutch strain P11 of PaMMV was studied using antisera against P101 and ToMV. The Bulgarian strain P101 was serologically very closely related to the PaMMV strain P11 originated from the Netherlands. The performed immunodiffusion tests with homologous and heterologous antisera allow P101 to be arranged in different series of serological relationship with typical tobamovirus strains. PaMMV is more related to TMGMV and PMMoV than to TMV and ToMV. Key words: paprika mild mottle virus, tobamoviruses, immunodiffusion, mmunoelectrophoresis, serological relationship. Introduction Paprika mild mottle virus (PaMMV) is a member of the Tobamovirus group. It is spread in Eurasian and has a comparatively narrow host range [1].Rast first reported for isolate P11 found in Capsicum annuum, the Netherlands [8]. Later, it was defined as a separate virus [4]. In Bulgaria, in 1979 tobamovirus strain named P101 was isolated from greenhouse pepper cv. Bruinsma Wonder [7]. Subsequently, it was identified as PaMMV by means of biological, serological, electrophoretic and molecular investigations [9, 13]. Strain P101 does not differ from the rest tobamoviruses in the virion morphology, stability in sap, dried and frozen leaves and preparation as well as in thermal inactivation point. The ratio A260/A280 of a purified preparation is 1.3. Strain P101 has very close electrophoretic mobility to PaMMV P11 [10, 11, 13, 14]. The Bulgarian strain is from pathotype P1 and overcomes the L1 resistance gene of C. annuum cultivars but at least 28 accessions of C. chinense are resistant to P101 and susceptible to P11. Strain P101 induces symptomless infection in tomato in contrast to P11 that is in-capable of infecting this plant species [13, 14]. Similar biological properties were established for the Japanese isolate (PaMMV-J), which occasionally infected tomato plants although with poor effectiveness due to low levels of both replication and cell-to-cell movement [6]. The Bulgarian strain of PaMMV also differs from P11 by the reaction of Nicotiana tabacum cv. Samsun N’N’ and N. sylvestris (N’ gene) [13, 14]. Several investigations of the serological properties of strain P101 bymeans of homologous or heterologous antisera have been carried out. Different tobamoviruses have been used for these comparative studies [9, 12, 13]. To date there is not a systematized serological characteristic of the strain. The purpose of the present work was a more complete serological characterization of the Bulgarian PaMMV strain P101 and investigation of itsrelationshipwithothertobamoviruses by immunodiffusion methods. Materials and Methods Viruses. The following viruses and strains were used: tobacco mosaic virus (TMV) - U1 and Sm; tomato mosaic virus (ToMV) - GM-0; tobacco mild green mosaic virus (TMGMV) - U2; PaMMV – P101 and P11, pepper mild mottle virus (PMMoV) - GP2 and GP3. All strains are from the collection of the National Bank for Industrial Microorganisms and Cell Cultures, Bulgaria [2]. Virus propagation and purification.The strains
were propagated as follows: strains U1 and Sm in N. tabacum cv. Samsun N’N’ (resistance
gene N’ from N.sylvestris); GM- Counter immunoelectrophoresis. It was performed in 0.9 % agarose gel in 0.75 M veronal buffer, pH 8.6, at 150 V, 10 °C for 2.5-3.5 hours. Antisera against P101 and ToMV diluted 1:3 – 1:10 were used [3, 16]. The virus preparations in concentration of 1 mg/ml were applied at different times. The gels were stained with Coomassie Brilliant Blue R-250 [15]. Double
diffusion test. Radial diffusion
was made in 0.7 % agarose gel in 0.075 M veronal buffer, pH 8.6,
containing Results and Discussion The serological relationship of the Bulgarian PaMMV strain P101 with other tobamoviruses was studied by means of counter immunoelectrophoresis and double diffusion test. A homologous antiserum as well as an antiserum against ToMV was used. The obtained results are summarized in Table 1, where the strains are provisionally designated as standard and tested antigens. Table 1. Serological relationship* of PaMMV P101 with typical tobamovirus strains.
* spur
formation by standard antigen (+) and tested antigen (++); fusion of the
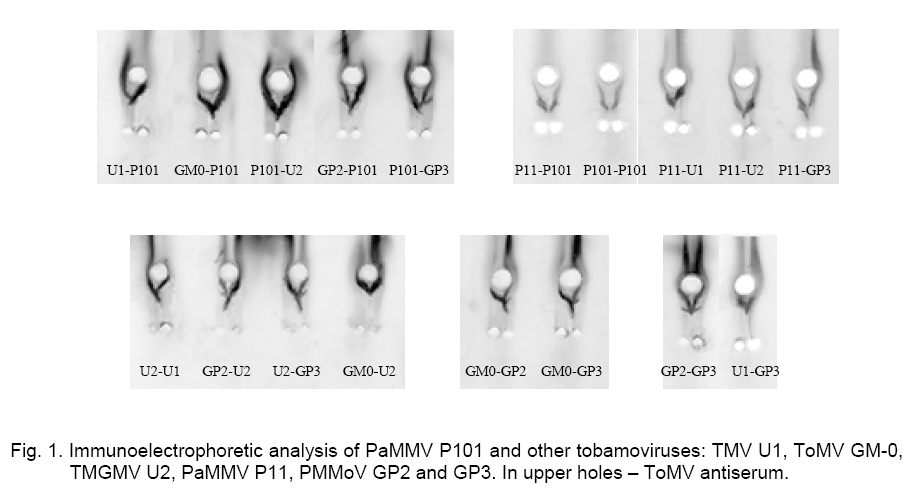
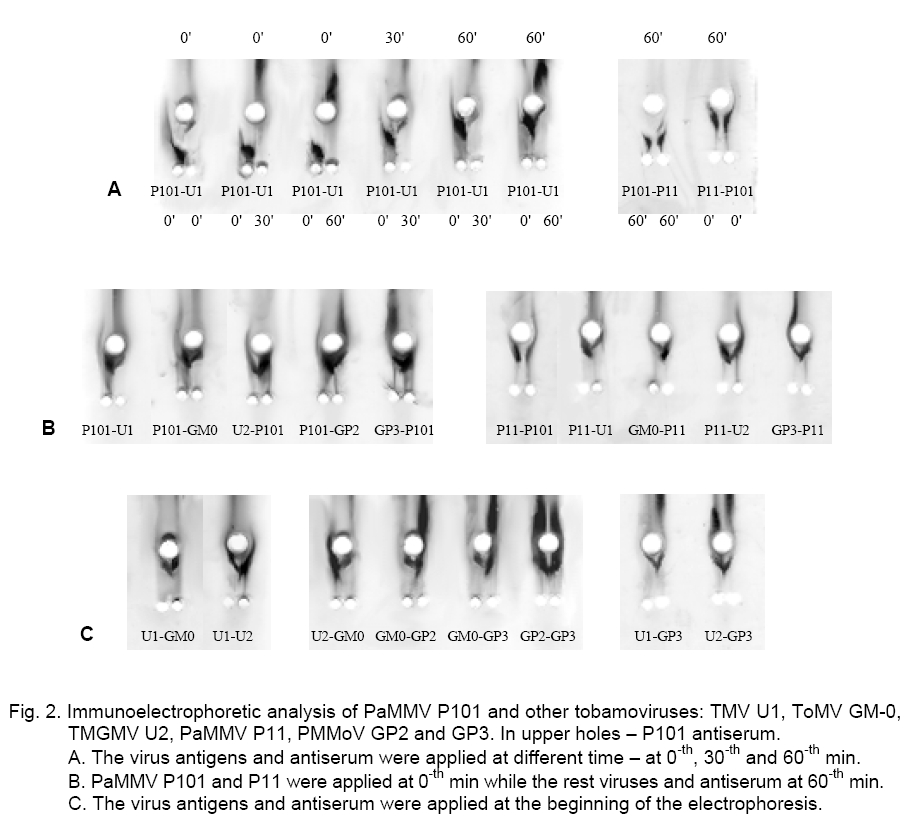
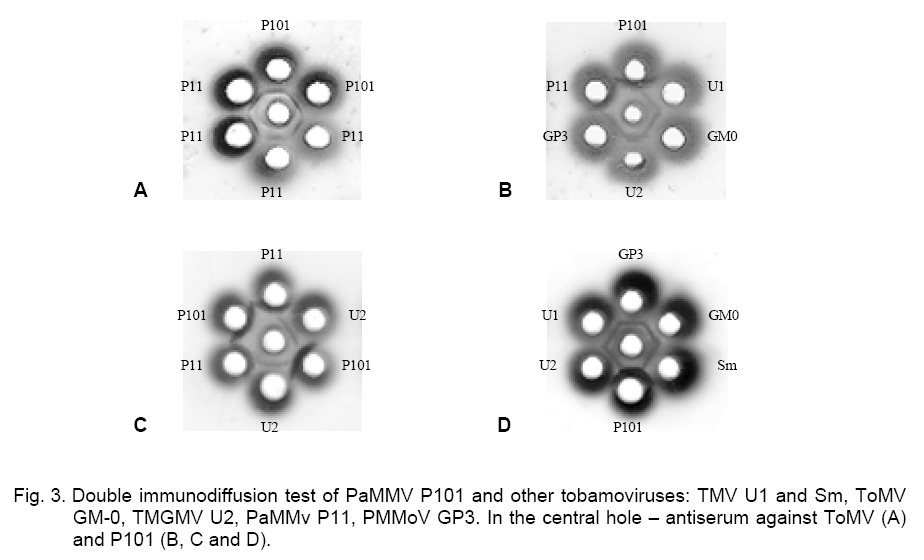
precipitation lines (-). The immunoelectrophoretic analysis with the ToMV antiserum showed that the PaMMV strains P101 and P11 reacted in the same way. They formed spur in respect to TMV and PMMoV – GP2 and GP3, while ToMV and TMGMV produced spur towards P101 and P11 (Figure 1). The ToMV antiserum differentiated clearly TMV, ToMV, PMMoV and PaMMV (Table 1).The used serum did not distinguish ToMV and TMGMV as well as the two PMMoV strains. By testing one against each other, the precipitation lines of P101 and P11 did not touch well but the strains were considered rather serologically identical. Besides that, the PaMMV strains were less related to ToMV than TMGMV and were more closely related to ToMV than PMMoV and TMV. Both PMMoV strains exhibited a second precipitation line probably due to the antiserum reaction with whole virions and capsid protein, which had a different electrophoretic mobility (Figure 1). After counter electrophoresis of P101 and application of the homologous antiserum, it was established that the reaction was very strongly expressed, so the precipitation line was situated near to the start and far from the lines of the other strains. The behavior of P11 was similar. Several variants of electrophoresis were applied, in which the antigens and antiserum were dropped at different times (Figure 2, A). It was found that the application of P101 and P11 at the 0-th min and of the rest viruses and anti-serum at the 60-th min was the most convenient. However, in some cases the electrophoresis termination led to insufficient clearness of the precipitation lines and made difficult the results interpretation. PaMMV strains P101 and P11 formed spur towards TMV, ToMV, TMGMV and PMMoV (Figure 2, B). The precipitation pattern of both strains appeared quite equally but a line touching was not observed even at multi-fold reagents dilutions. Nevertheless, the serological reaction was assumed as a fusion of the precipitation lines. From the rest viruses, U2 produced spur in respect to U1, GM-0 and GP3; GP3 to U1 and GM-0; U1 to GM-0 (Figure 2 C, Table 1). The carried out experiments for the serological relationship of P101 and other tobamoviruses by immunodiffusion test showed the following results. Using a heterologous antiserum against ToMV, the strains P101 and P11 were not distinguished (Figure 3, A). The application of the homologous antiserum gave a possibility to observe the presence of a small spur produced from P101 towards P11, by varying the antigen concentrations (Figure 3, C). In this test the reactions of the PaMMV strains were also much more greatly pronounced in comparison to those of the other tobamoviruses, whose precipitation lines were fairly weaker and unclear. P101 and P11 produced spurs towards all other viruses. Strain U2 formed spurs to TMV, ToMV and PMMoV (Figure 3, B, D, Table 1). The performed immunodiffusion tests with the homologous to P101 antiserum allow the Bulgarian PaMMV to be arranged in the following series of serological relationship with typical tobamovirus strains: PaMMV P101 ³ PaMMV P11 > TMGMV U2 > PMMoV GP2, GP3 > TMV U1> ToMV GM-0. When the ToMV antiserum is used, the respective serological order is: ToMV GM-0 = TMGMV U2 > PaMMV P101, P11 > PMMoV GP2, GP3 > TMV U1. The application of different antisera gives a possibility for better characterization of the serological properties of the tobamoviruses and their strain variability. The presented data show that PaMMV P101 is serologically very closely related to the PaMMV strain P11 originated from the Nether-lands. Ruíz del Pino et al. established that the coat proteins of P101 and the Dutch strain P11 differed by two amino acids. They also found 97 % sequence identity in the noncoding 3’-UTR region of both strains because of six nucleotide substitutions leading to structural changes [9]. It could be supposed that similar nucleotide variations are a reason for the little differences in the serological properties between PaMMV P101 and P11 as well as in their electrophoretic mobility [13]. These nucleotide variations as well as additional changes in other virus genes probably determine the differences in the reaction of some plant hosts to both PaMMV strains. References
Copyright 2009 - National Bank for Industrial Microorganisms and Cell Cultures - Bulgaria The following images related to this document are available:Photo images[cc09013f2.jpg] [cc09013f3.jpg] [cc09013f1.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}