 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Journal of Culture Collections, Vol. 6, No. 1, 2009, pp. 112-121 Two new serum free and protein free cell strains, derived from hep-2 cell line: cultural conditionsand proliferation activity Marian M. Draganov1*, Stanimir H. Fransazov2, Daniel M. Draganov1, Marianna A. Murdjeva2 and Nikola K. Popov1 1University of Plovdiv ”P. Hilendarski”, Department
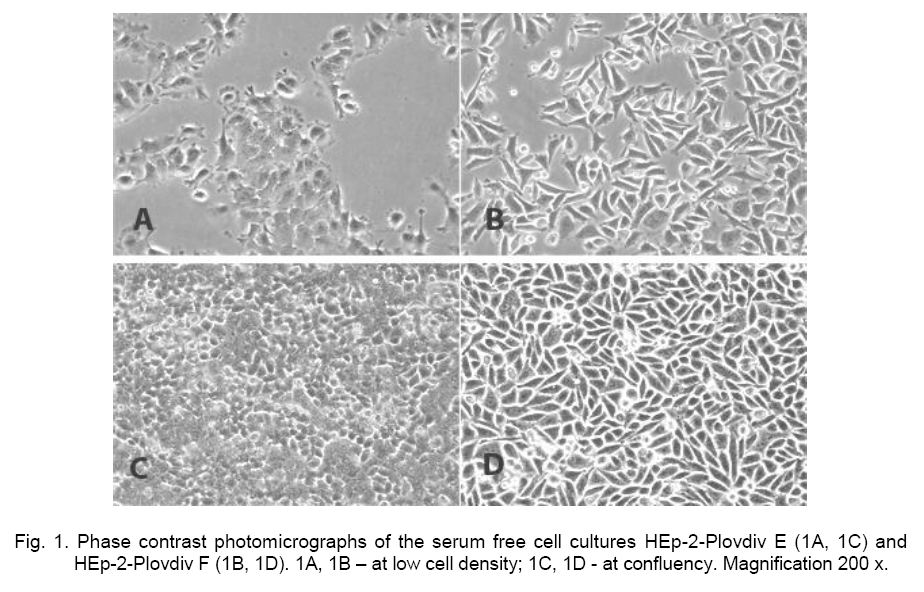
of Biology, Cell Biology Laboratory, 24 “Tsar Assen” Str, 4000 Plovdiv, Bulgaria; Code Number: cc09014 Summary Two new cell strains – HEp-2-Plovdiv E and HEp-2-Plovdiv F were derived from the HEp-2 cell line. They could be cultivated indefinitely in vitro in DMEM/Ham’s F12 1:1medium. These are serum-free and protein-free cultures because they grown only in medium that is chemically defined. Serum-free cultures could be distinguished by morphological characteristics, growth dynamics, sensitivity to trypsinization, proliferation activity and doubling time. The number of cell population’s doublings of the initial carcinoma cell line and the two serum-free cultures was calculated. The dynamics of cell proliferation was investigated and doubling time for HEp-2 cell line (22.6 h), HEp-2-Plovdiv F strain (25.8 h) and HEp-2-Plovdiv E strain (28.4 h) was calculated. DMSO of 7 % was added in the growth medium for storage in liquid nitrogen. Optimized parameters for cultivation and subcultivation of the cell cultures are suggested. Possible ways of application of the new serum free cell cultures, based on preliminary studies for viral cultivation and detection of serum autoantibodies are discussed. Key words: serum free cell cultures, cell line, HEp-2, McCoy-Plovdiv. Introduction HEp-2 cell culture was established in 1952 by Alice E. Moore, Lillian Sabachewski and Helene W. Toolan [8]. The cell line was obtained from tumors induced in young rats after inoculation of human epidermoidcarcinoma tissue of 56 years old man. The isolation of cells in vitro was performed in a mixture of bovine amniotic liquid, embryonic extract, human and horse serum and balanced salt solution [43]. HEp-2 cell line is described in ATTC under ¹ CCL-23. The cell line has got biosafety level 2 (cells contain Papovavirus). Cells of this line contain HeLa marker chromosomes [8]. HEp-2 cell line is one of the most widely-spread cultures and is applied in studies on different viruses – herpes simplex virus (HSV)[4, 31, 42]; measles virus [29]; adenoviruses [24]; monkey immunodeficiency virus [23]; enteroviruses [2]; polioviruses [5]; hepatitis B [35]; arboviruses; VSV – vesicular stomatitis virus (Indiana strain); respiratory syncitial virus; ECHOvirus, Coxsackie, and Newcastle virus [1]. In their life span many microorganisms interact with eukaryotic cells and cause damage and cell death. HEp-2 cells are an appropriate in vitro cell system for studying and diagnosis not only of viruses, but also of bacteria and their products as well - Chlamydiapneumoniae [9, 34]; Listeria monocytogenes [37], Stenotrophomonas maltophilia [16]; Brucella abortus and Brucella melitensis [36]; Streptococcus pyogenes [17, 27]; Staphylococcus aureus [25]; Escherichia coli [46]; Yersinia enterocolitica [41]; Salmonella typhimurium [15]; Campylobacter jejuni [7]; Haemophilus ducreyi[10]; Treponema pectinovorum [44];Trypanosoma cruzi [39]. HEp-2 cell line finds a broad application in medicine for diagnostics of antinuclear antibodies [6, 20, 21] and in cell biology - for studying cell receptors [2]. HEp-2 carcinoma cells are appropriate for investigating tumors [45], and for experiments, associated with antitumor therapy [18, 33]. Materials and Methods Cell cultures. HEp-2 cell line was kindly provided by Dr. M. Trajancheva from the Laboratory of Virology, Regional Inspectorate for Protection and Control of Public Health in Plovdiv. The culture has been successfully maintained since1970, and used for virus cultivation in the laboratory by Dr. Mandulov (personal communication). Cells were grown in a closed system in Medium 199 with Hank’s salts and 10 % bovine serum. In our laboratory HEp-2 cells were grown in DMEM/Ham’s F-12 (1:1) medium supplemented with 10 % FCS and antibiotics, and cultivated in a Heraeus incubator (Germany) at 37 °C, 5 % CO2 and high humidity. The monolayer of HEp-2 cells was subcultured after trypsinization with 0.05 % trypsin and 0.02 % EDTA, dissolved in phosphate buffer without calcium and magnesium. The cells were preserved in liquid nitrogen in DMEM/Ham’s F-12 (1:1) containing 20 % FCS and 10 % DMSO. Media and solutions. The cells were cultivated in the following media: DMEM/Ham’s F12 (1:1) (D-8900, Sigma). The media were dissolved, sodium hydrogencarbonate was added, pH was adjusted and sterilization was performed according to the recommendations of the Producer Company. The following reagents and solutions were used: Fetal Bovine Serum (0643, Sigma); Penicillin, Streptomycin (Antibiotic-Co, Bulgaria); trypsin (0152-17-7, DIFCO), EDTA (11280, Serva; E-5134, Sigma), DMSO (D-2650 Sigma), Sodium bicarbonate (S-5761, Sigma). PBS: 0.02 % EDTA in PBS: 0.4 % Trypan blue solution (T8154, Sigma). Cell culture labware. Cells were grown in polystyrene cell culture flasks of 25 or 75 cm2 (Corning; Nunclon, Nunc; Falcon, Becton Dickinson); in 12 or 24-well cell culture plates (Linbro, Flow and Falcon, Becton Dickinson); and frozen in cryogenic vials (430489 Corning, USA). Trypsinization and subculturing of monolayer of serum-free HEp-2-Plovdiv E and HEp-2-Plovdiv F cultures. The serum-free cell cultures of the strains HEp-2-Plovdiv E and HEp-2-Plovdiv F were grown in DMEM/Ham’s F-12 (1:1) at the same conditions of cultivation as those for the serum-supplemented HEp-2 cells. HEp-2-Plovdiv E. Every 5th
or 6th day after setting up the culture the cells were trypsinized
with 0.075 % trypsin and 0.02 % EDTA for 10-15 min and washed once
with PBS by centrifugation for 8-10 min at HEp-2-Plovdiv F.Every 5th or 6th day after setting
up the culture the cells were trypsinized with 0.025 % trypsin and
0.02 % EDTA for 6-8 min and washed once with PBS by centrifugation for
8-10 min at Preservation of cells. Freezing.Cells were trypsinized in a confluent state. After centrifugation for 5-7 min they were resuspended in cold (+4 °C) freezing medium (DMEM/Ham’s F-12 supplemented with 7 % DMSO). The cell suspension with a concentration of 1 – 2 x 106 cells/ml was dispensed in cryotubes (precooled in ice-cold water bath). They were put in a penoplast box in a freezer (Ultra Low, Model 192, Sanyo) at -80 °C for at least 48 h. Then the cells were transferred into a cryocontainer with liquid nitrogen at -196 °C. Thawing. After drawing out the cells from the liquid nitrogen, they were immediately put into a hot water bath at 37-45 °C till entire thawing of the medium. Afterwards the cells were carefully transferred into a preheated at 37 °C culturing medium and kept in an incubator. One or two hours later, half of the medium was discarded and replaced with a fresh aliquot. Development of cell cultures from seeding till confluent monolayer. Cells were seeded in 24-well plates with initial plating density: 0.8 x 104, 1.6 x 104, 2.4 x 104, 3.2 x 104, 4.0 x 104 and 8.0 x 104 cells/cm2 (numbered from 1 to 6). The volume of the media was 1 ml. Every 24 hours the state of the cell culture was observed under an inverted microscope till the 168th hour. The cells were examined and photographed on a Laboval 2 microscope (Karl Zeiss, Germany). Then the cell monolayer was trypsinized and the number of viable cells was determined by Trypan blue test in a haemocytometer. Dynamics of the cell proliferation. Cells were plated in 24-well plates at a density of 3.2 x 104 cells/cm2. Every 24 hours the cells from three wells were trypsinized and pooled into a single cell suspension. Cell number was determined in a haemocytometer. Live cells were detected by Trypan blue test. Statistics and calculations. The data were statistically
processed using the Student’s t-test. The results are presented as mean value
and standard deviation ( The formula used for calculation of “population doublings” in a single passage was: NPD = 3.33 x log10 (N/N0), where NPD is the number of population doublings, N is the number of cells in the growth vessel at the end of the growth period and N0– the number of cells plated in the growth vessel. The “doubling time” was calculated afterSchaeffer [40] using the formula: DT = t x lg2/lg (N/N0), where DTis the doubling time, t – period of time when the culture is in log phase, N0 – the number of cells at the start of the t period and N – the number of cells at the end of the t-period. Results and Discussion Introduction of cells from the HEp-2 cell line in conditions of serum free cultivation HEp-2cells were adapted and cultivated in DMEM/Ham’s F-12 (1:1) medium containing 10 % FCS. After ten successful subcultivations we tried to introduce the same cells into serum free conditions. The same grown medium (DMEM/ Ham’s F-12, 1:1) was used in two variants of cultivation – direct inoculation of cells in serum free conditions (cells were transferred right a-way in a medium without serum), and cultivation by stages’ adaptation. In the second variant the removal of serum was performed through gradualdecrease of serum concentration from 10 % to 0 %. The serum concentration was reduced with 1-2 % at each stage. As a result of the direct inoculation along with cloning, two strains of the initial HEp-2 culture were established and stabilized. They were named HEp-2-Plovdiv E (the form of the cells morphologically is mostly epithelial-like), and HEp-2-Plovdiv F (the largest number of cells are morphologically more elongated, fibroblast-like shaped) (Figure 1, A and B). The two new serum free strains vary in their confluent culture state as well (Figure 1, C and D). For the subcultivation procedure we applied our previous experience with the serum free McCoy-Plovdiv cell line [14]. When trypsin was added directly to the cultural medium, we did not observe cell layer disaggregation as it happened with McCoy-Plovdiv cells [14]. The serum free HEp-2-Plovdiv E cell layer remained intact even after raising trypsin concentration over 0.8 % and prolonging the treatment time above one hour. The cell disaggregation process of both serum free strains was effective in the following sequence: growth medium was removed, culture was washed out with 0.02 % EDTA in PBS solution for a few minutes; EDTA solution was replaced by a fresh one and trypsin was added. Trypsin at final concentrations of 0.5 %, 0.25 %, 0.1 %; 0.05 % and 0.025 %, and EDTA concentrations of 0.08 %; 0.04 %, 0.02 %, 0.01 % were studied. HEp-2-Plovdiv E and HEp-2-Plovdiv F cells showed different sensitivity to trypsin during the subcultivation procedures. Evaluation was made regarding duration and effectiveness of disaggregation, cell vitality after trypsinization and cell growth after plating. For HEp-2-Plovdiv E the optimal combination was established to be 0.02 % EDTA and trypsin concentration of 0.075 %, and for HEp-2-Plovdiv F – 0.02 % EDTA and 0.025 % trypsin. These parameters of the disaggregating agents brought the cell monolayer to a suspension of single cells after 10-15 min processing. The comparison between trypsinization procedures of the two new serum free cell cultures - HEp-2-Plovdiv E and HEp-2-Plovdiv F, with the McCoy-Plovdiv serum free cell line [12], pointed out several important differences, characteristic for the HEp-2 Plovdiv serum-free cultures: i. compulsory using of chelating agent; ii. higher trypsin concentration; iii. longer time needed for enzyme treatment of the culture. The analysis of results brîught us to the assumption that the cells of the two new serum free cell strains, HEp-2-Plovdiv E and HEp-2-Plovdiv F, built up stronger cell-substrate and cell-cell contacts in comparison with these of the McCoy-Plovdiv cell line [12]. By establishment of these contacts an important function probably have specialized cell molecules like integrins and cadherins. Calcium ions are linked in big amounts to integrin molecules. They play an important role in intercellular communications and adhesion of the cells to their substrate [3; 26]. In in vitro conditions fibroblasts or epithelial cells spread on the cultivation vial wall through building specialized connection domains situated on the lower surface, called focal adhesions. Integrins participate in the construction of these specialized structures [30]. Calcium ions are of extreme importance for the structure and functional activity of cadherins, a big family of glycoproteins that determine Ca2+-dependent cell-cell adhesion and transmission of signals from the extracellular matrix to the cytoplasm [3]. They are observed in adhesions junction and desmosomes [19]. These facts support our observation of disaggregation absence in both serum-free HEp-2 cell lines when chelating agent is not used. Its role is to remove or decrease the amount of bivalent calcium ions, promoting the strength of cell-cell interaction. An important stage in the disaggregation process of the cell culture is to terminate the action of trypsin and EDTA. Under serum free conditions this is achieved through washing out the cells and removal of trypsin. It could be done through several washings or centrifugation in large amount of a washing liquid. Both approaches (alone or in combination) could be applied successfully for the cells of the serum free HEp-2-Plovdiv E and HEp-2-Plovdiv F strains. To guarantee a prolonged preservation of cell cultures we studied the process of cell freezing and thawing. Cells were frozen in growth medium only in the presence of DMSO at different concentrations as a cryoprotectant (Table 1). The results coincide with our previous observations on the McCoy-Plovdiv cell line [13]. We assume that the combination of DMEM/ Ham’s F-12 (1:1) medium supplemented with 7 % DMSO is appropriate for a long term preservation in liquid nitrogen of the serum free HEp-2-Plovdiv E and HEp-2-Plovdiv F strains. Table 1. Effect of DMSO concentration in the preservation medium on the cell survival* of the serum free HEp-2-Plovdiv-Å and HEp-2-Plovdiv Å cultures after thawing.
*Cell vitality is estimated after Trypan blue inclusion test. Dynamics of cell morphology from seeding till monolayer formation Development of cell cultures from the time of inoculation until confluent state was studied. Results for the HEp-2 cell line, plated at six different seeding densities (from 0.8 õ 104 to 8 õ 104 cells/ñm2), are shown in Table 2 (A). An inversely proportional correlation between plating density and time necessary to form a confluent layer was established. In cultures at initial cell densities from 1 to 6, with increase of the number of cells inoculated, less time is necessary for the formation of a dense cell layer. In the culture with highest inoculation density (8 õ 104 cells/ñm2), confluent monolayer was observed 24-48 hours after seeding, while for the cultures with lowest initial density (0.8 õ 104 cells/ñm2) it happened after 144th hours. Table 2. Generalized microscopic assessment of the development of HÅp-2, HEp-2-Plovdiv Å, HEp-2-Plovdiv F cell cultures from different initial densities to confluent monolayer.
Legend: IPD, initial plating density; C, confluent
monolayer; SC, subconfluent monolayer; SC+C, subconfluent monolayer with large
confluent islands; C+SC, > 80 % confluent cultures; Table 2 (B) shows the results about cell monolayer formation for the serum free HEp-2 Plovdiv-E cell culture. For all six initial plating densities (from 0.8 õ 104 cells/ñì2 to 8.0 õ 104 cells/ñm2), 24 hours after plating cultures represent populations of single cells, rounded and hardly spread. Forty eight hours after seeding we observed a subconfluent state of the culture with the highest plating density of 0.8 õ 104 cells/ñm2, which became confluent at the 72th hour after plating. Decrease of the initial plating density leads to the need of longer time for the culture to reach a confluent state. For 4.0 õ 104 cells/ñm2 it happened 120 hours after seeding; for 3.2 õ 104 cells/ñm2 after 140 hours; and for 2.4 õ 104 cells/ñm2 as well as 1.6 õ 104 cells/ñm2 – after 168 hours. HEp-2-Plovdiv E cell culture, plated with the lowest initial plating density of 0.8 õ 104 cells/ñm2, remained as a culture of single cells or groups of cells that had never reached the confluent monolayer state for the studied period. Data for the development of cell cultures of the serum free strain HÅp-2 Plovdiv F from different initial cell densities are shown in Table 2 (C). At the lowest plating density of 0.8 õ 104 cells/ñm2 the culture did not develop to confluent and even subconfluent state. Cultures became confluent 168 hours after seeding, when plated at an initial density of 1.6 õ 104 cells/ñm2 or 2.4 õ 104 cells/ñm2. Confluent state was observed after 120 hours at initial cell density of 3.2 õ 104 cells/ñm2, on the 96th hour after plating of 4.0 õ 104 cells/ñm2 and on the 72th hours at the highest density of 8.0 õ 104 cells/ñm2. An interesting finding is that the formation of subconfluent and confluent state of the HÅp-2 Plovdiv F culture is faster in comparison with that of the HÅp-2 Plovdiv E serum free cells. In this aspect, the cell monolayer development of the serum free HÅp-2 Plovdiv F cells resembles the development of the HÅp-2 serum-supplemented cells from a plating density higher than 1.6 õ 104 cells/ñm2. Cell number at the end of the experiment allowed us to determine how much the culture had increased growth during the studied period, and to calculate the number of doublings of the cell populations for the three cell cultures. Data are presented in Table 3 (A, B and C, respectively for HEp-2, HEp-2-Plovdiv E and HEp-2-Plovdiv F cells). It is evident that in all samples there was more than three fold increase of the cell population of HEp-2 and HEp-2-Plovdiv F with exception of the samples with the highest plating density. For the HEp-2-Plov-div E cell line (Table 3B) the cell number is lower as compared to that of the other two cell lines and there are fewer than 3 population doublings. For the culture with the highest initial plating density of 8.0 õ 104 cells/ñm2 the population doublings are less than 1.5. This finding is not surprising for the serum-supplemented culture. It could be explained by the fact that in the culture with the highest plating density, the cells most quickly cover the space needed for development and reach a confluent stage, and after that the cell division and the cell number are slowing down. This is determined by more rapid exhaust of the nutritional products from the medium, and an increased amount of secreted metabolites. That changes the medium and the color becomes yellow, which is an indication of acidification. Thus the combination of free space on the substrate for cultivation and the quality of the medium are factors regulating the cell population increase. Besides these facts there is another phenomenon in the serum free HÅp-2 Plovdiv Å and HÅp-2 Plovdiv F cultures, explaining the lower number of cells, and it is the cell death. As a result of this experiment it was established that the serum free cells were capable to develop in vitro starting from a broad range of initial plating densities. However, as the analysis shows, cultures of the lowest initial plating density do not reach the stage of a cell monolayer (the cultures are of “island type”) or the cell monolayer formation is very slow. On the other side, when cultures are plated at a very high density there is no optimal development and cell production. We would recommend for cultivation of the serum free HÅp-2-Plovdiv F and HEp-2-Plovdiv E cultures initial plating density of 1.6 õ 104 cells/ñm2 to 4.0 õ 104 cells/ñm2, which corresponds to a plastic flask with a surface of 25 ñm2 – from 104 to 100 õ 104 cells in total. Thus for subcultivation from a flask of 25 cm2 à dilution of the cell suspension in split ratio 1:4 or 1:8 could be performed. Table 3. HÅp-2, HEp-2-Plovdiv Å and HEp-2-Plovdiv F cells plated with increasing initial cell density in 24 wells plate and cultivated for 168 hours, trypsinized and counted (live cells only).
Legend: NPC, number of plated cells; FPD, final population density; NPDs, number of population doublings. Calculated values for NPD for
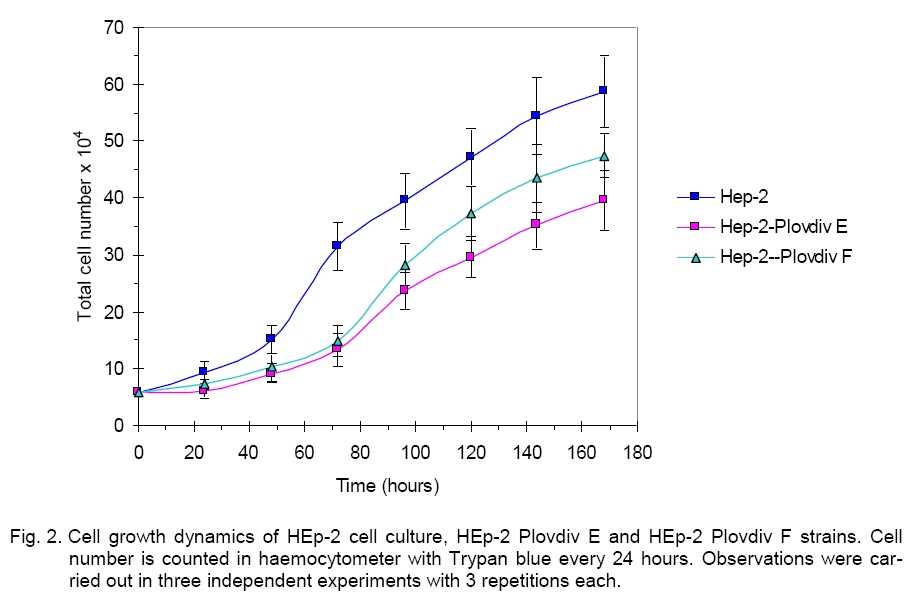
each culture are presented. Cell growth dynamics Results from the cell proliferation of HEp-2, HÅp-2-Plovdiv F and HEp-2-Plovdiv E cultures are presented in Figure 2. The study was carried out on cell cultures with initial plating density of 3.2 õ 104 cells/ñm2 for all three cell lines. The analysis of the way of cell development shows that HÅp-2-Plovdiv F and HEp-2-Plovdiv E cell lines are quite similar. After subcultivation, HEp-2 plated cells demonstrated shorter period of adaptation and after 24hours an exponential increase of the culture was observed. The increase in the cell number was highest between 48 - 72 hours after plating. Cell number at given intervals (15.18 ± 2.54 x 104 cells for 48th hour and 31.62 ± 4.28 x 104 cells for 72th hour) allowed us to calculate the doubling time – 22.6 h. Òhe most rapid increase in cell number for the serum free strains was detected in the interval from the 72nd to the 96th hour. Data for this interval of the graphics were 13.22 ± 2.37 x 104 cells and 23.74 ± 3.59 x 104 cells for HEp-2-Plovdiv E, and 14.85 ± 2.94 x 104 cells and 28.31 ± 3.24 x 104 cells for HÅp-2 Plovdiv F. Doubling population time for the serum free cultures was 28.4 h for HEp-2-Plovdiv-Eand 25.8 h for HÅp-2 Plovdiv F. Conclusion After summarizing the data from the study and the analysis of the results, we do believe that two new strains have been developed and established - HEp-2-Plovdiv E and HEp-2-Plovdiv F, serum free cell cultures. They differ in their cell morphology, the state of the confluent culture, the dynamics of the cell growth, the sensitivity to trypsin and disaggregation of the cell layer. Serum, serum-derived or other exogenous products of microbial, plant or animal origin with undefined nature are not used in any single stage of culturing the serum-free cells and their preservation in liquid nitrogen. The complete development of the cultures is ensured only by the chemically defined medium DMEM/Ham’s F-12 (1:1). All this lead us to determine the two new strains as serum free, and the cell cultural systems as free of exogenous proteins. Our previous investigation carried out for several years allowed us to establish the serum free cell line McCoy-Plovdiv that showed convincingly the advantage of the serum free culture compared to the initial serum-supplemented McCoy line [11, 13, 32]. Apart from the new possibility for application of these cells as substrates for antinuclear antibodies testing [47], effect of dental monomers bis-GA and bis-GMA on expression of histo-blood group antigens [38], and cultivation of HIV was investigated (Prof. R. Argirova, personal communication). We do hope that the new serum free HEp-2-Plovdiv E and HEp-2-Plovdiv F strains will be good models for further studies in experimental and clinical medicine. Preliminary results confirmed the possibility these cells to be used for cultivation of Adenoviruses, Herpes simplex virus, and Respiratory Syncitial virus. A study on testing antinuclear and
antimitochondrial antibodies from human sera using these new serum free
cultures has been initiated. The serum free HÅp-2-Plovdiv F culture correlates very well with the cells of the cell
line HEp- The two
new serum free strains are registered as
numbers 8575 and Acknowledgements. The work was supported by two projects: 07B4 - from the Section “Scientific and Developmental Work”, University of Plovdiv “P. Hilendarski”, Bulgaria, and DRI-01-2006 - from Medical University-Plovdiv and National Science Fund, Ministry of Education and Science, Bulgaria. References
Copyright 2009 - National Bank for Industrial Microorganisms and Cell Cultures - Bulgaria The following images related to this document are available:Photo images[cc09014f1.jpg] [cc09014f2.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}