 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Journal of Culture Collections, Volume
2, 1997-1998 pp. 51-59
TAXONOMIC INVESTIGATION OF MONASCUS PURPUREUS 94-25 STRAIN Tanya Rasheva1*, Jean – Noel Hallet2 and Anna Kujumdzieva1 1The Sofia University, Biological Faculty,
Department of General and Industrial Microbiology, 8 "Dragan Tzankov" st., 1421
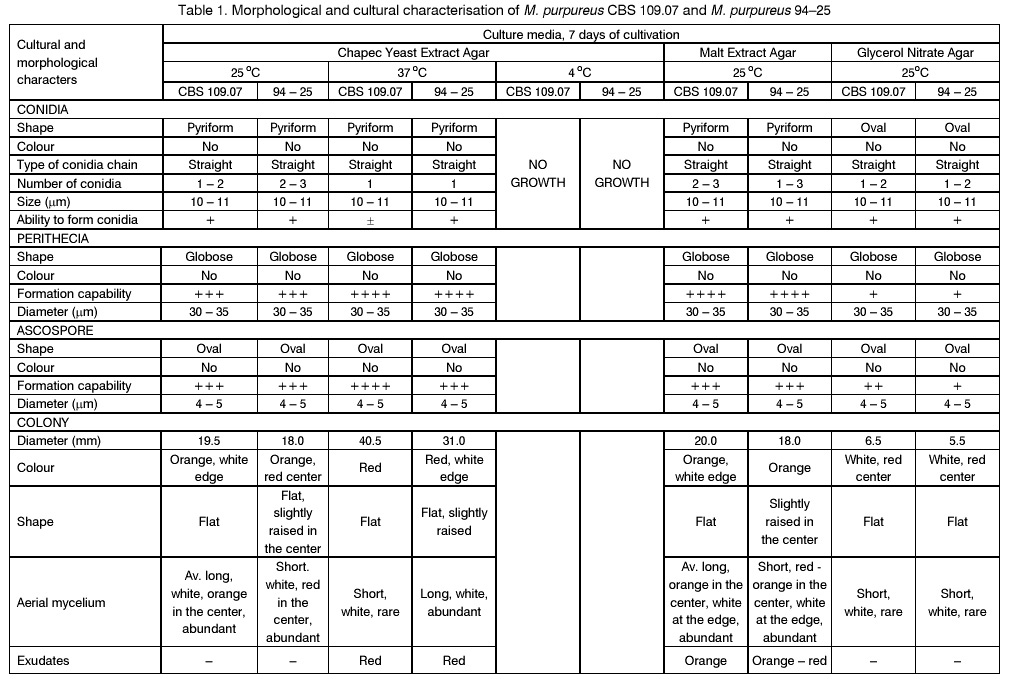
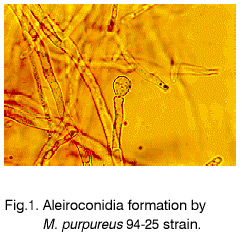
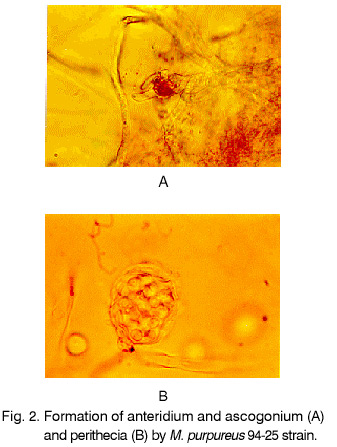
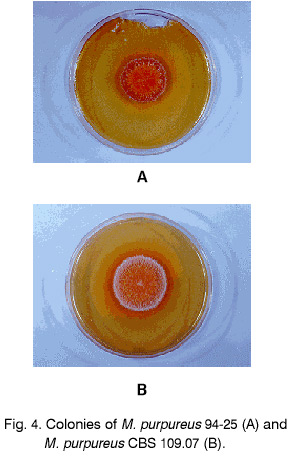
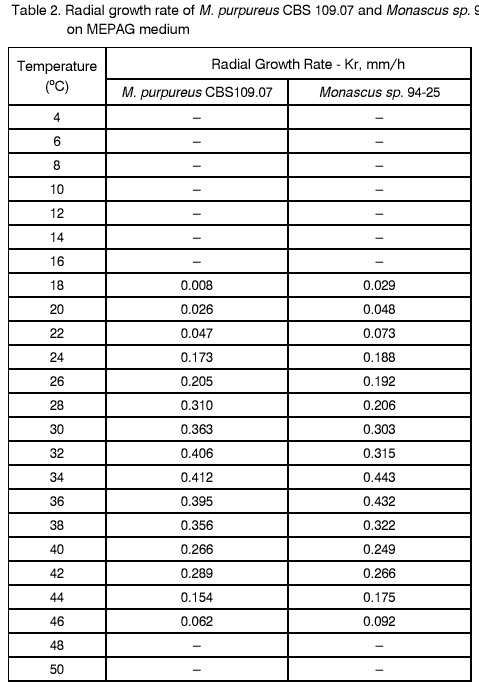
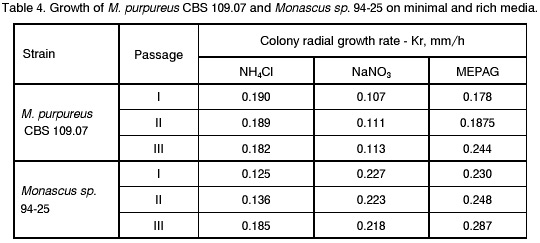
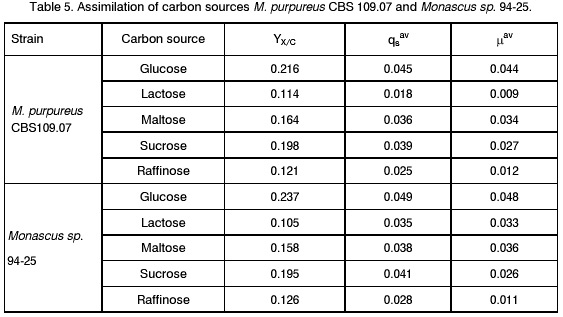
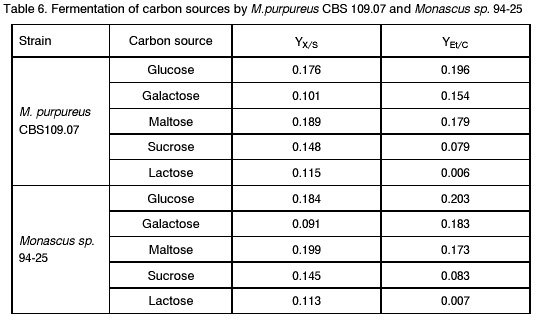
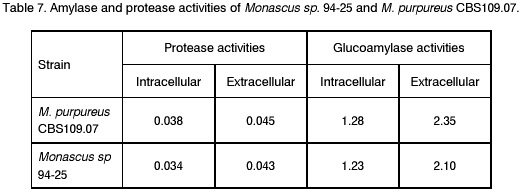
Sofia, Bulgaria; Code Number: cc98009 Summary Monascus sp. 94-25 strain was isolated from red rice with the purpose of red pigments submerged production. Morphological characterisation on the taxonomically important for the genus media, showed that the strain produced cleistothecia with oval ascospores and aleiroconidia. Comparison with a referent strain Monascus purpureus Went 109.07 was made and it was proved that there were no considerable differences between both strains. Monascus sp. 94-25 was a prototroph and had optimal growth temperature 34°C. Investigation of the fermentation and assimilation capacity of both strains was performed. It was found that both of them assimilated well glucose, while highest fermentation ability was observed when grown on galactose. Similarly to the referent strain Monascus sp. 94-25 could grow on starch and protein containing natural substrates. Regarding the morphological and biochemical investigations the newly isolated strain 94-25 could be considered as Monascus purpureus. Introduction Monascus fungi are well known as producers of a family of structurally related hexaketide pigments which are yellow and red in colour. They have been used in Asia for many centuries to colour and flavour food and beverages [4, 12]. These natural colorants are of practical interest because of the difficulties in obtaining red pigments safe for use in food industry [5]. Different strains belonging to genus Monascus are used in pigment production [1, 9]. Isolation of new strains is still of particular interest because of necessity to obtain microorganisms with suitable kinetic characteristics for submerged cultivation. The aim of this investigation is to determine morphological, biochemical and some kinetic characteristics of Monascus sp. 94-25 strain isolated with the purpose for submerged production of red pigments. Materials and Methods Microorganisms. Wild strain Monascus sp. 94-25 was isolated from red rice on MEPAG medium containing (g/l): glucose - 20, peptone - 10, malt extract - 5, agar - 22, pH 6.0 at 30°C. As a referent strain Monascus purpureus Went CBS 109.07 was used. Both strains were maintained on MEPAG medium. Taxonomic investigation. The experiments were performed following the procedure of Hawksworth and Pitt [2]. Both strains were cultivated on the following culture media: CYA (Chapek - Yeast Extract Agar) containing (g/l): NaNO3 - 3.0, K2HPO4 - 1.0, KCl - 0.5, MgSO4.7H2O - 0.5, FeSO4.7H2O - 0.01, sucrose - 30.0, yeast extract - 5.0, agar - 15, pH 6.0. MEA (Malt Extract Agar) containing (g/l): malt extract - 20, peptone - 1, glucose - 20, agar - 15, pH 6.0. G25N (25 % Glycerol Nitrate Agar) (g/l): NaNO3 - 3.0, K2HPO4 - 1.0, KCl - 0.5, MgSO4.7H2O - 0.5, FeSO4.7H2O - 0.01, yeast extract - 5.0, agar -15, pH 6.0. After agar was dissolved, 250 g glycerol to 750 g medium was added. All culture media were sterilised at 121°C. The cultivation was performed in Petri dishes at 25°C for 7 days for all culture media. Additionally cultivation on CYA medium at 4°C and 37°C was carried out. During the cultivation, morphological and cultural characteristics of both strains were studied. Auxotrophy. The auxotrophy of the wild and the referent strain was studied on Chapec – Dox (CD) medium, containing (g/l): glucose - 30, NaCl - 0.5, MgSO4.7H2O - 0.5, KH2PO4 - 1.0, FeSO4.7H2O - 0.01. ZnSO4.7H2O - 0.001, pH 6.0. As nitrogen sources NH4Cl – 2.5 and NaNO3 – 3.0 g/l were used. The media were solidified with 22 g/l Difco purified agar. MEPAG medium was used as a control. The cultivation was performed in Petri dishes with 20 ml solid medium at 30°C for 7 days. Growth temperature. The growth temperatures were determined on MEPAG medium in the range 4 – 50°C. Assimilation of carbon sources. The assimilation ability of both strains was studied in submerged cultures, on CD medium with NaNO3 as nitrogen source and different carbon sources. The cultivation was carried out in 500 ml Erlenmeyer flasks with 100 ml medium, on a rotor shaker at 220 rpm, at 30°C for 7 days. The biomass was harvested by filtration, dry weight of the biomass and the residual concentration of sugar were determined. The parameters mav, qsav and YX/C were calculated according to Pirt [7]. Fermentation of carbon sources. The fermentation capacity of both strains was studied on medium containing (g/l): Yeast nitrogen base (YNB, Difco) - 6.7, sugar – 10, pH 6.0. The cultivation was carried out in bottles with a stopper and a needle at 30°C for 28 days. The residual sugar content was determined following the method of Somogyi [8]. The ethanol formation was analysed according to the method of Herbert et al. [3]. Cell – free extract was obtained after mechanical disintegration of biomass with silica and phosphate buffer (0.05 M, pH 7.8) in ratio 1:3:1 for 1 hour at 4°C. The suspension was centrifuged at 1000 g for 20 min at 4°C and stored at -20°C for enzymatic assays. Determination of protease activity. Investigation of the ability of the strains to assimilate proteins was performed in Erlenmeyer flasks – 500 ml with 100 ml CD medium, containing 1 % of casein as a carbon source. The submerged cultivation was carried out on a Rotor shaker at 220 rpm, at 28°C for 7 days. The biomass was harvested by filtration, twice rewashed and stored at –20°C. The proteolityc activity was determined in the culture filtrate (extracellular) and in the cell free extract of the biomass (intracellular) according to the procedure of Yasuda et al. [11]. Reaction mixture contained 0.6 ml of 1 % solution of casein, 0.3 ml of 0.1 M sodium acetate buffer (pH 5.5) and 0.3 ml cell free extract. The mixture was incubated at 37°C for 10 min. The reaction was terminated by adding 1.5 ml of 5% trichloracetic acid. The solution was filtered and 0.4 ml 0.5 N Folin reagent was added. After standing at room temperature for 30 min, the absorbency at 750 nm was read and the amount of tyrosine equivalents was estimated. The amount of protein, which causes release of 1 mmol tyrosine per minute, was defined as one Unit protease activity. Determination of glucoamylase activity. For determination of the glucoamylase activity both strains were cultivated in Erlenmeyer flasks – 500 ml with 100 ml CD medium, containing 1 % of starch as a carbon source. The submerged cultivation was carried out on a Rotor shaker at 220 rpm, at 28°C for 7 days. The biomass was harvested by filtration, twice rewashed and stored at –20°C. The glucoamylase activity was determined either in the culture filtrate and in the cell free extract, following the procedure of Yasuda and Kuwae [10]. Results and Discussion The isolated from red rice strain is studied taxonomically according to the procedure of Hawsworth and Pitt [2]. M. purpureus CBS109.07 is used as a referent strain in all experiments. The main morphological and cultural characteristics of the investigated microorganisms are shown in Table1. There is no significant difference between both strains during cultivation on all taxonomically important media. The newly isolated strain forms oval or pyriform aleiroconidia, single or in a chain up to 3 - 4 (Fig. 1). Perithecia can be observed mainly on CYA and MEA media. The microorganism forms actively anteridia and ascogonia (Fig. 2, A). In later stage, colourless or lightly coloured in orange cleistothecia with oval ascospores are developed (Fig. 2, B). The hyphae are septated, 2 – 4 mm width. Some of them are filled up with lipid droplets or pigments (Fig. 3). The colony shape, colour, aerial mycelium and exudate formation between the two strains (Fig. 4) are similar. Determination of optimal growth temperature is done using colony diameter and radial growth rate Kr [7]. Kr is determined in the temperature interval 4 - 50°C. The fungal growth is recorded by measuring the diameter of the colonies in two directions at right angles and taking the means of at least 6 replicates. The results for the two strains are presented in Table 2. There is no significant difference in the values of Kr between both strains at one and the same temperature conditions. In accordance with this investigation, the minimal, optimal and maximal growth temperatures are determined (Table 3). These results indicate that M. purpureus CBS109.07 and the newly isolated strain grow up to a temperature of 46°C and can be refereed as thermotolerant ones. A study of the strains growth requirements is also made. M. purpureus CBS109.07 and the newly isolated strain are cultivated on minimal (CDA) nutrient media with two different nitrogen sources (ammonium and nitrate) and a rich (MEPAG) one. The growth is estimated by measuring Kr values within three seven-days passages (Table 4). Kr is used as a parameter for a comparison between the nutrient requirements of the strains during the above mentioned conditions. The obtained results indicate that there is no significant difference between both strains on CDA medium with NH4Cl. Nearly twice higher Kr is measured during cultivation of the newly isolated strain on medium with NaNO3. The same difference is observed also during cultivation on MEPAG medium. All these findings indicate that the newly isolated strain possess higher growth capacity on medium with nitrate as nitrogen source. In order to investigate the capability for utilisation of different carbon sources a number of carbon substrates are tested. The results are shown in Table 5. Using different growth characteristics (YX/S, qs and mav), the efficiency of the utilisation of these carbon sources is estimated. Both strains assimilate relatively well glucose (YX/S is about 0.21 – 0.24) and possess close values of qs and mav . Concerning other tested carbon sources, these parameters show slow growth and utilisation. The fermentation capability of both strains is also studied (Table 6). Significant ethanol production during fermentation of glucose, galactose and maltose is observed. The higher capacity for ethanol formation is obtained with galactose as a carbon source. Another important characteristic of Monascus fungi is their capability for production of proteases and amylases [6]. So they are used for obtaining these enzymes by cultivating them on different natural substrates. Investigations of amylase and protease activities are shown in Table 7. There is no significant difference between both strains. On the basis of all morphological and biochemical investigations it could be concluded that the new strain 94-25 belongs to species Monascus purpureus. Acknowledgements These investigations were partially supported by Grant B-406 of the National Science Foundation, Ministry of Education, Republic Bulgaria. References
Copyright 1998 - National Bank for Industrial Microorganisms and Cell Cultures - Bulgaria The following images related to this document are available:Photo images[cc98009t5.jpg] [cc98009t4.jpg] [cc98009f1.jpg] [cc98009t2.jpg] [cc98009t3.jpg] [cc98009t7.jpg] [cc98009t6.jpg] [cc98009t1.jpg] [cc98009f2.jpg] [cc98009f3.jpg] [cc98009f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}