 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
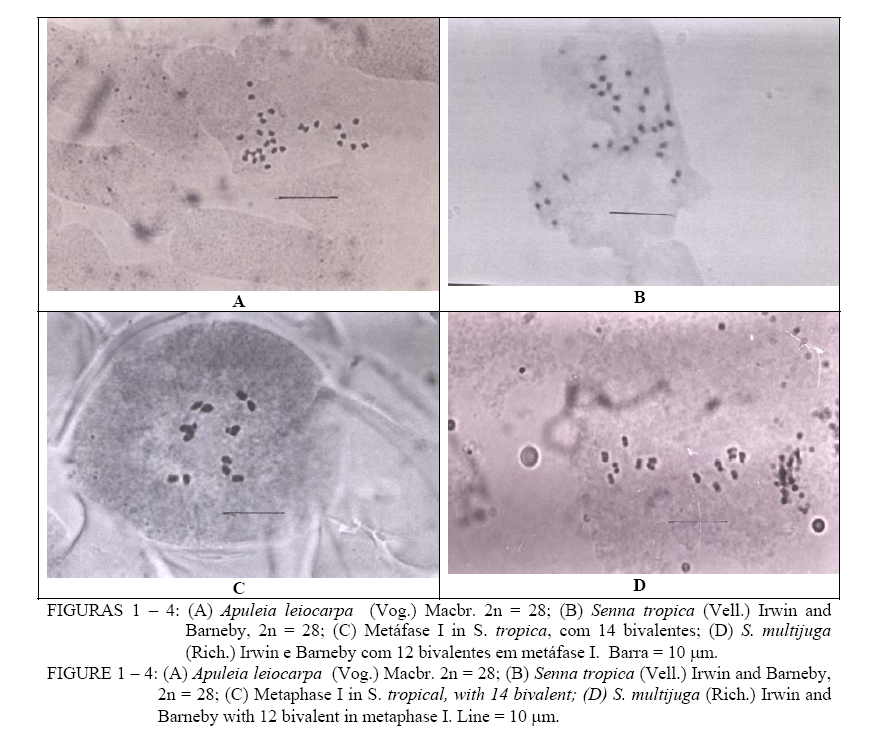
Ciência Florestal, Vol. 15, No. 3, 2005, pp. 241-248 CITOGENÉTICA DE ESPÉCIES ARBÓREAS DA SUBFAMÍLIA CAESALPINIOIDEAE – LEGUMINOSAE DO SUL DO BRASIL CYTOGENETICS OF CAESALPINIOIDEAE – LEGUMINOSAE TREE SPECIES FROM SOUTHERN BRAZIL Elaine Biondo1, Sílvia Teresinha Sffoggia Miotto2 , Maria Teresa Schifino-Wittmann3 1. Bióloga, Msc., Doutoranda pelo Programa de Pós-Graduação em Botânica, Universidade Federal do Rio Grande do
Sul, Rua João Telles, 453/32, Bairro Bom Fim, CEP 90035-121, Porto Alegre (RS). elainebiondo@hotmail.com Code Number: cf05023 RESUMO
Determinação do número de cromossomos e análise do comportamento meiótico são excelentes contribuições aos estudos de relações taxonômicas e padrões evolutivos dentro de grupos de espécies vegetais. Espécies de leguminosas arbóreas da subfamília Caesalpinioideae, ocorrentes na Região Sul do Brasil, têm sido pouco analisadas do ponto de vista citotaxonômico. Este trabalho teve por objetivos determinar o número de cromossomos e analisar o comportamento meiótico de onze espécies arbóreas dessa subfamília. O número de cromossomos na maioria das espécies analisadas foi 2n = 28 cromossomos (x = 14). Foi encontrado 2n = 24 (x = 12) em Senna multijuga (L.C. Rich.) H. S. Irwin & Barneby e Schizolobium parahyba (Vell.) Blake,e 2n = 26 (x = 13) em Peltophorum dubium (Spreng.)Taub. Os núcleos apresentaram padrão arreticulado em todas as espécies e o comportamento meiótico foi regular em seis espécies estudadas. Sugere-se ampliação das coletas e análises citogenéticas em mais indivíduos e espécies de forma a gerar informações adicionais que permitam conclusões mais abrangentes sobre este grupo tão importante. Palavras-chave: Leguminosae; cromossomos; meiose. ABSTRACT Chromosome counts and meiotic behaviour analysis are excellent contributions for studies on taxonomic relationships and evolutionary patterns in plants. The tree legume species of the subfamily Caesalpinioideae have been poorly analysed cytogenetically. This work aimed at determining the number of chromosomes and analysing the meiotic behaviour in 11 species of Caesalpinioideae. Basic chromosome numbers in most of the analysed species were 2n = 28 (x = 14). Senna multijuga (L. C. Rich.) H. S. Irwin & Barneby and Schizolobium parahyba (Vell.) Blake presented 2n = 24 (x = 12) and Peltophorum dubium (Spreng.) Taub. 2n = 26 (x = 13). In all the species nuclei presented an areticulate pattern and a regular meiosis in the six species studied. Further collections and cytogenetic analyses with a major number of individuals and species are suggested, in order to provide additional data for a more comprehensive study on this group of plants. Key words: Leguminosae; chromosome; meiosis. INTRODUÇÃO A família Leguminosae, com cerca de 18 mil espécies (Polhill et al., 1981), tem ampla distribuição no mundo, sendo consenso entre leguminólogos que seja dividida em três subfamílias: Papilionoideae, Mimosoideae e Caesalpinioideae. A subfamília Caesalpinioideae compreende 150 gêneros e 2.700 espécies, com distribuição cosmopolita (Polhill et al., 1981; Judd et al., 1999). Dentre as subfamílias, Caesalpinioideae é a menos estudada e entendida (Herendeen,2000), tendo sido considerada por Burkart (1987) como a mais primitiva, da qual derivaram as subfamílias Papilionoideae e Mimosoideae. Está dividida em cinco tribos: Caesalpinieae, Cassieae, Cercideae, Detarieae e Macrolobieae, sendo que a última não tem representantes no sul do Brasil (Lewis e Polhill, 1998). Atualmente, uma grande quantidade de estudos biosistemáticos vem sendo realizada com esse grupo, com indicativos de que Caesalpinioideae não é um grupo natural, compreendendo tribos parafiléticas (Käss e Wink, 1996; Doyle et al., 1997; Herendeen, 2000; Bruneau et al., 2001). Dentre as quatro tribos ocorrentes na Região Sul do Brasil, duas são consideradas parafiléticas: as tribos Caesalpinieae e Cassieae (Bruneau et al., 2001). Nessa região, pela análise de exsicatas, coletas a campo e revisão de literatura, observou-se que ocorrem 16 gêneros e cerca de 58 espécies nativas (Bortoluzzi, 2003, comunicação pessoal). Um grande número de espécies florestais nativas faz parte dessa subfamília. Muitas delas são utilizadas como ornamentais e na arborização urbana como Cassia leptophylla, Senna multijuga, Senna macranthera, Parkinsonia aculeata (Santos e Teixeira, 2001); outras são utilizadas na medicina popular como Bauhinia forficata (Lorenzi e Abreu, 2002; Silva et al., 2002); na recuperação de áreas degradadas (Resende e Kondo, 2001; Ademir Reis, comunicação pessoal),como é o caso de Senna macranthera e Senna multijuga, espécies que propiciam relações ecológicas interespecíficas importantes. Muitas possuem madeira de elevada importância econômica, como o pau-brasil (Caesalpinia echinata Lam.) (Cardoso et al., 1998) e a grápia, Apuleia leiocarpa (Auler, 1997; Marchiori, 1997; Backes e Irgang, 2002). A última é uma espécie muito explorada (Ruschel et al., 2003) e, provavelmente, está ameaçada de extinção. Além disso, as espécies arbóreas nativas distribuem-se em diferentes formações vegetais, com importância ecológica inquestionável nestes diferentes ecossistemas. Informações sobre cromossomos são relevantes em estudos sistemáticos e evolutivos, abrangendo, desde a simples contagem, até detalhes da citogenética molecular que são a fronteira da pesquisa atual (Stace, 2000). Historicamente, quando estudos citogenéticos de espécies arbóreas são comparados aos de espécies cultivadas e/ou nativas com valor agronômico, sua limitação é evidente, restringindo-se a informações básicas sobre sua estrutura genômica e a inclusão dessas espécies em programas de melhoramento e conservação (Schlarbaum, 2000). Dentre as espécies arbóreas, o grupo das gimnospermas é o mais estudado citogeneticamente, destacando-se estudos com coníferas e pináceas (Muratova e Sedelnikova, 2000). Um evento marcante para a citogenética de espécies arbóreas foi a descoberta da origem triplóide de Populus tremula L., sendo esse o primeiro exemplo do potencial das informações citogenéticas no melhoramento dessas espécies (Schlarbaum, 2000). Gêneros como Eucalyptus L’Herit., Pinus L., Populus L. e Juglans L., em que predomina o hábito arbóreo, são os mais estudados do ponto de vista do melhoramento genético e biotecnológico (Studart-Guimarães et al., 2003), evidenciando a falta de estudos botânicos e genéticos com espécies arbóreas nativas. Dentre as espécies de leguminosas com hábito arbóreo e arbustivo, estudadas citogenéticamente no Brasil, são citadas as dos gêneros Caesalpinia L. (Alvez e Custódio, 1989; Beltrão e Guerra, 1990); Erythrina L. (Forni-Martins e Cruz, 1996); Leucaena Benth. (Cardoso et al., 2000; Boff e Schifino-Wittmann, 2003); Sesbania Scop. (Forni-Martins et al., 1994; Forni-Martins e Guerra, 1999) e Machaerium Pers. (Mendonça-Filho et al.,2002). Considerando que no Brasil estão catalogados 188 gêneros de leguminosas, com ocorrência significativa em diversas formações vegetais, sobretudo na Mata Atlântica, com elevado predomínio de representantes arbóreos (Lima, 2000), os gêneros estudados até o momento representam cerca de 2% do total, evidenciando a necessidade de mais estudos com esse amplo grupo vegetal. A relação entre números cromossômicos elevados e diferentes hábitos depende dos grupos taxonômicos que estão sendo analisados. Além disso, altos números cromossômicos ocorrem em espécies florestais. Porém, a freqüência de poliploidia provavelmente está relacionada com a composição florística das formações florestais (Forni-Martins e Martins, 2000). Um gênero de leguminosas com muitas espécies arbóreas e, comprovadamente de origem poliplóide, é Leucaena (Boff e Schifino-Wittmann, 2003). Análises citogenéticas em espécies de leguminosas arbóreas na Região Sul do Brasil são quase-inexistentes. Para a subfamília Caesalpinioideae, apenas Apuleia leiocarpa foi estudada por Auler (1997). A falta de trabalhos deve-se, provavelmente, às dificuldades relacionadas à obtenção do material botânico, especialmente botões florais jovens, já que muitos indivíduos chegam a medir de 30 a 40 m de altura, como é o caso de Schizolobium parahyba (guapuruvú) e Peltophorum dubium (canafístula) (Backes e Irgang, 2002). Este trabalho faz parte de um projeto maior que visa a estudar as Leguminosae – Caesalpinioideae da Região Sul do Brasil do ponto de vista taxonômico, ecológico e citogenético, e teve por objetivos contar o número de cromossomos de 11 espécies arbóreas nativas, e analisar o comportamento meiótico de algumas delas. MATERIAL E MÉTODOS Coleta do material botânico As sementes e os botões florais necessários à realização deste trabalho foram coletados, em sua maioria, nos locais de distribuição natural de cada espécie (Tabela 1). Cada acesso corresponde a um indivíduo, representado, sempre que possível, por uma exsicata depositada no Herbário Instituto de Ciências Naturais (ICN), do Departamento de Botânica da UFRGS, Porto Alegre, RS. Número cromossômico As sementes foram retiradas do fruto, escarificadas com lixa e postas a germinar em placas de petri, à temperatura de 28oC. O tempo de germinação foi, em média, de 5 dias. Quando as radículas apresentavam cerca de 2 cm, foram coletadas e pré-tratadas com solução saturada de paradiclorobenzeno por 24 horas à temperatura de 4oC, para obtenção de maior número de células em metáfase. Após, as radículas foram lavadas e fixadas em Carnoy 9:3:1 (etanol : clorofórmio : ácido acético) por 24h, à temperatura ambiente e, em seguida, estocadas em álcool 70% em geladeira. Para o preparo das lâminas, foi utilizada a coloração de Fuelgen, de acordo com Guerra e Souza (2002); as radículas foram lavadas em água destilada, hidrolizadas em HCl 1 N a 600C por 10 minutos; estas foram novamente lavadas e colocadas no corante por cerca de 30 minutos. A extremidade da raiz foi esmagada com uma gota de carmim propiônico. Foram contadas, no mínimo, quatro células e, no máximo, 38 células por acesso (Tabela 1), com cromossomos visíveis e bem dispersos. As melhores metáfases foram fotografadas. O padrão de condensação de núcleos interfásicos foi definido seguindo-se a classificação de Guerra (1985). Análise meiótica A análise meiótica foi realizada em seis espécies. Para essa análise, os botões florais foram fixados em Carnoy por 24 horas, à temperatura ambiente, sendo transferidos para álcool 70% e conservados à temperatura abaixo de 0oC. No preparo da lâmina, as anteras foram esmagadas e os meiócitos corados com carmim propiônico. Foram analisadas, no mínimo, cinco e, no máximo, 59 células por acesso (Tabela 1). Maior atenção foi dada para as seguintes fases da meiose: diacinese, metáfase I e anáfases I e II, as quais indicam relações de homologia entre os cromossomos homólogos e sua orientação durante a metáfase I e II. RESULTADOS E DISCUSSÃO Foram analisados 29 acessos de 11 espécies (Tabela 1), representando 19% do total de espécies pertencentes à subfamília Caesalpinioideae na Região Sul do Brasil. As espécies analisadas apresentaram números cromossômicos variando de 2n = 28 em Senna oblongifolia, Senna tropica (Figura 2), Apuleia leiocarpa (Figura 1), Cassia leptophylla, Bauhinia forficata e Parkinsonia aculeata (Tabela 1); 2n = 26 em Senna macranthera e Peltophorum dubium e 2n = 24 em Senna multijuga e Schizolobium parahyba. De Gleditsia amorphoides, foram analisadas poucas células em razão da dificuldade na sua obtenção. Desta, foram observados apenas 2n = 26 em uma célula e 2n = 28 em duas células. Segundo Castronovo (1945), Gleditsia amorphoides apresenta 2n = 28 cromossomos, considerados de difícil análise por serem cromossomos pequenos. Na maioria das espécies analisadas (Figura 1 a 4), os cromossomos são pequenos, com cerca de 2 mm. O pequeno tamanho dos cromossomos de angiospermas arborescentes também foi observado por Mehra e Bawa (1969). O padrão de condensação de núcleos interfásicos é arreticulado. As variações na estrutura de núcleo interfásico geralmente são constantes dentro de espécies e gêneros, com poucos gêneros apresentando variação interespecífica. No caso das espécies e gêneros aqui analisados, não houve variação na estrutura do núcleo interfásico, embora sejam citadas variações entre tribos na família Leguminosae (Guerra, 1985). Todas as espécies analisadas apresentaram comportamento meiótico regular, com formação de bivalentes em metáfase I e segregação regular em anáfases I e II (Figura 3 e 4). Altas freqüências meióticas (mais de 90%) foram observados previamente por Biondo et al. (2003) nas espécies aqui analisadas. TABELA 1: Número cromossômico de espécies arbóreas da subfamília Caesalpinioideae (Leguminosae) coletadas na Região Sul do Brasil. TABLE 1: Chromosomic number of arboreal species of the sub family Caesalpinioideae (Leguminosae) collected in southern Brazil.
Em que: AC = número do acesso; N/ Cél. = número de células; n = número haplóide de cromossomos; 2n = número diplóide de cromossomos; # = sem exsicata; · cultivada. Considerando que o índice meiótico é um complemento para a análise meiótica e um indicador de regularidade (Löve, 1949), pode-se afirmar que as espécies analisadas são estáveis meioticamente com produção de gametas masculinos balanceados. A análise realisada, com raras exceções, indicou estabilidade meiótica e produção de gametas viáveis. Esses são aspectos importantes no caso da inclusão dessas espécies em programas de melhoramento genético e produção de sementes. As espécies das tribos Cassieae e Caesalpinieae apresentaram número básico cromossômico x = 12, 13 e 14, confirmando o sugerido por Irwin e Barneby (1982). A tribo Cercideae inclui espécies que apresentam número básico x = 14 cromossomos (Goldblatt, 1981), conforme observado também neste trabalho. Neste estudo, Senna multijuga apresentou 2n = 24 cromossomos (x = 12). Esse número é observado, sobretudo, em espécies do gênero Caesalpinia (Beltrão e Guerra, 1990; Gonzales et al., 2002). Goldblatt (1988) citou para Senna multijuga 2n = 26 cromossomos, x = 13. Neste estudo, o comportamento meiótico foi regular, com formação de 12 bivalentes na diacinese e na metáfase I (Figura 3). Assim, n = x = 12 é o provável número básico de Senna multijuga. O número básico proposto para Senna macranthera é x = 13 cromossomos (Tabela 1). Bandel (1974) citou número de cromossomos 2n = 24 para essa mesma espécie coletada na Região Sudeste. Constatou-se, portanto, uma variação no número de cromossomos entre populações de uma mesma espécie com diferentes distribuições geográficas. Variações no número cromossômico em Apuleia leiocarpa foram observadas quando foram comparados diferentes locais de coleta. Neste trabalho, Apuleia leiocarpa coletada em Floresta Estacional Decidual apresentou 2n = 28 cromossomos (Figura 1). Auler et al. (1998) encontraram 2n = 26 cromossomos em indivíduos coletados em Floresta Estacional Semidecidual. Para Apuleia leiocarpa também foi sugerida a ocorrência de polissomatia, com número cromossômico variando de 20 a 28 nas células de um mesmo indivíduo (Auler et al., 1998). A análise de indivíduos de diferentes populações da mesma espécie é muito importante, já que variações significativas de números cromossômicos entre populações são encontradas em literatura (Forni-Martins e Martins, 2000). No caso de Apuleia leiocarpa,o maior número possível de indivíduos e de populações deveria ser analisado, devido à grande importância dessa espécie como componente da Floresta Estacional Decidual bem como pela sua ampla utilização para produção madeireira (Ruschel et al, 2003). A variação no número cromossômico observada nas espécies analisadas é corroborada por outros autores em trabalhos realizados em outras regiões do Brasil. Bandel (1974), ao estudar 35 espécies de leguminosas das Regiões Sudeste e Centro-Oeste, encontrou número de cromossomos 2n = 28 nos gêneros Cassia, Dimorphandra Schott e Parkinsonia; 2n = 24 nos gêneros Apuleia e Cassia e 2n = 20 cromossomos no gênero Pterogyne Tull., sugerindo a existência de variação quanto ao número básico de cromossomos de x = 12 e 14. O número cromossômico básico nas espécies da subfamília Caesalpinioideae parece ser x = 7. A poliploidia começa a estabelecer-se, aparentemente, na evolução da subfamília, para x = 14, com uma linhagem de x = 12 cromossomos (Goldblatt, 1981). A tribo Cassieae pode ser considerada uma tribo com espécies tetraplóides cujo número básico é x = 14, com exceção do gênero Chamaecrista (Breyne) Moench com x = 7. Este é, talvez, derivado por disploidia de um ancestral poliplóide com x = 14. A tribo Caesalpinieae apresenta número básico x = 14 (Goldblatt, 1981). O aumento no número de cromossomos foi um passo importante na evolução de espécies tropicais lenhosas (Morawetz, 1986). Stebbins (1971) sugeriu que espécies com números básicos maiores que x = 12 cromossomos são poliplóides, derivados de ancestrais com baixos números básicos. A mesma sugestão é feita por Goldblatt (1981) para Leguminosae. Corroborando o que esses autores sugeriram, todas as espécies aqui analisadas são poliplóides. Na literatura, são encontrados poucos trabalhos que relacionam altos números cromossômicos com o hábito em Leguminosae – Caesalpinioideae. Ohri et al. (1986) encontraram variações abruptas em conteúdo de DNA em espécies arbóreas de Cassia s.l., quando comparadas com espécies arbustivas e herbáceas. Kumari e Bir (1989) observaram predominância de cromossomos metacêntricos em espécies arbóreas e arbustivas de Caesalpinioideae indianas sem diferenças no número de satélites e cariótipo. No Brasil, análises citogenéticas de espécies arbóreas, arbustivas e herbáceas dessa subfamília, até o momento, são incipientes. Há necessidade de aprofundar os estudos relacionados à estrutura genética e aos sistemas reprodutivos, que ainda são pouco conhecidos na subfamília Caesalpinioideae, para melhor entendimento da sua evolução, sugere-se que novas coletas de material botânico, ocorrente em diversas formações vegetais, sejam realizadas para conclusões mais abrangentes. Enfatizam-se, portanto, a importância e a necessidade dos estudos de caracterização citogenética básica em espécies de leguminosas arbóreas nativas no sul do Brasil. CONCLUSÃO Onúmero cromossômico da maioria das espécies analisadas é 2n = 28. Senna macranthera e Peltophorum dubium apresentam 2n = 26 cromossomos, enquanto que Senna multijuga e Schizolobium parahyba apresentam 2n = 24 cromossomos. O padrão de condensação de núcleo interfásico é arreticulado, e o comportamento meiótico das seis espécies analisadas foi normal. AGRADECIMENTO À CAPES e ao CNPq pelas bolsas recebidas e ao PRONEX pelo subsídio para viagens de coletas. REFERÊNCIAS BIBLIOGRÁFICAS
Copyright 2005 - Ciência Florestal The following images related to this document are available:Photo images[cf05023f1.jpg] |
| |||||||||

{kind=link}
{kind=link}