 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ciência Florestal, Vol. 15, No. 4, 2005, pp. 343-357 PADRÃO E PROCESSO SUCESSIONAIS EM FLORESTAS SECUNDÁRIAS DE DIFERENTES IDADES NA AMAZÔNIA ORIENTAL SUCCESSIONAL PATTERN AND PROCESS IN SECONDARY FORESTS OF DIFFERENT AGES IN THE EASTERN AMAZON Maristela Machado Araújo1, Joanna Marie Tucker2, Steel Silva Vasconcelos3, Daniel Jacob Zarin4, Wilson Oliveira5, Patrícia Delamônica Sampaio6, Lívia Gabrig Rangel-Vasconcelos7, Francisco de Assis Oliveira8, Roberta de Fatima Rodrigues Coelho9, Débora Veiga Aragão5, Izildinha Miranda10 1Engenheira Florestal, Dra., Professora Adjunta do Departamento de Ciências Florestais, Centro de Ciências Rurais,
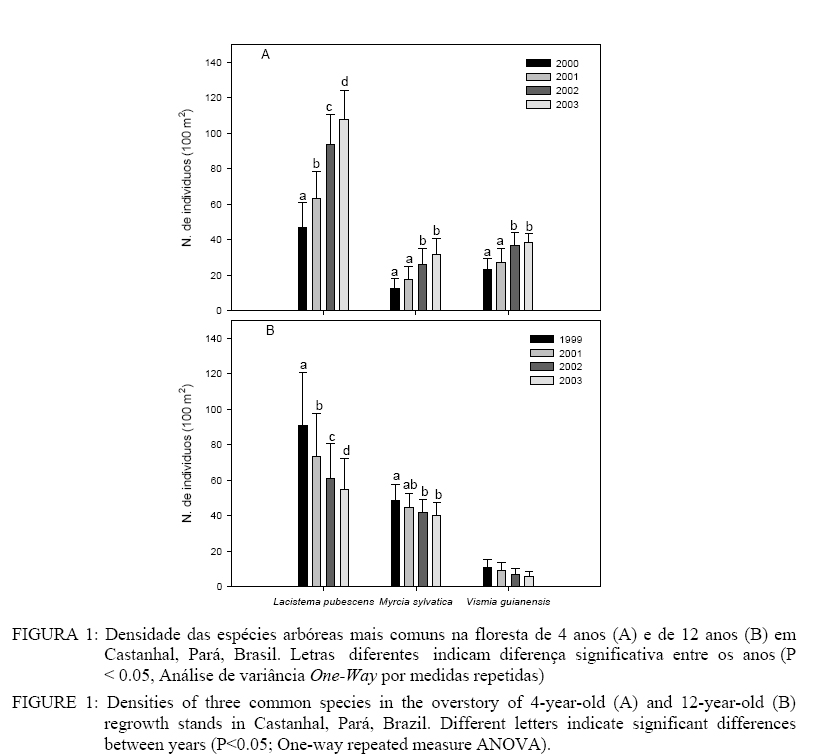
Universidade Federal de Santa Maria, CEP 97105-900, Santa Maria (RS). maristela.araujo@smail.ufsm.br Recebido para publicação em 26/04/2005 e aceito em 16/11/2005. Code Number: cf05034 RESUMOA maioria dos estudos de sucessão em florestas secundárias na Amazônia avalia sítios de diferentes idades, que representam uma cronoseqüência sucessional, em vez de monitorar um único sítio por vários anos pelo inventário contínuo. Este estudo comparou a composição e estrutura florísticas de espécies arbóreas com diâmetro a 1,3 m de altura (DAP) ≥ 1 cm, em sítios com 4 e 12 anos na Amazônia Oriental, e avaliou a mortalidade e o recrutamento em ambos os sítios baseados em dados de inventário contínuo durante 4 anos de estudo. As áreas de estudo foram abandonadas após múltiplos ciclos de uso agrícola de 7 a 10 anos, desde ~1940. Ambos os sítios são dominados pelas espécies arbóreas Lacistema pubescens e Vismia guianensis, com densidade de indivíduos, diâmetro, altura, área basal e riqueza de espécies significativamente maiores no sítio de 12 anos. A densidade de indivíduos, ao longo do tempo, foi crescente no sítio de 4 anos e decrescente no de 12 anos; o diâmetro, a altura e a área basal aumentaram nos dois sítios. No sítio de 4 anos, foi constatada uma taxa de recrutamento líquido crescente entre 2000-2001 e 2001-2002, que diminuiu entre 2002-2003, indicando redução gradual na colonização. No sítio de 12 anos, foi observada alta mortalidade líquida (13 e 11%), sobretudo nas duas primeiras avaliações, indicando o processo de autodesbaste. A combinação dos métodos de cronoseqüência e inventário contínuo aumenta substancialmente o entendimento do desenvolvimento sucessional. Palavras-chave: Amazônia; cronoseqüência; sucessão florestal; floresta secundária. ABSTRACT Most published studies of secondary forest succession in the Amazon examine stands of different ages that represent a successional chronosequence, rather than monitoring a single stand over the long-term. This study compares floristic composition and structure of tree species with diameter at 1.3 m height (DBH) ≥ 1 cm in a 4-year-old and a 12-year-old re-growth stand in the Eastern Amazon, and examines mortality and recruitment occurring within both stands based on repeated sampling carried out annually for four years. The study areas were abandoned after multiple agricultural cicles that lasted 7 to 10 years, beginning in ~1940. Both stands are largely dominated by the same tree species Lacistema pubescens and Vismia guianensis, with significantly higher stem density, diameter, height, basal area and species richness in the 12-year-old stand. In the 4-year-old stand there were measured an increase in annual net recruitment during the first two data collection periods but relatively lower net recruitment during the last evaluation period, indicating on-going but gradually weakening colonization. There were registered a high net mortality during the first two data collection periods in the 12-years-old stand with a relatively lower net mortality during the last evaluation indicating rapid self thinning. When used in combination, the chronosequence and the longitudinal approaches significantly strengthen the understanding of successional development. Key words: Amazon; chronosequence; forestry succession; secondary forest. INTRODUÇÃO A maioria dos estudos sobre o desenvolvimento de florestas secundárias na Amazônia tem sido conduzida partindo das cronoseqüências sucessionais, representadas por uma série de sítios de diferentes idades, mas com clima, solo, topografia, uso prévio da terra e potencial de colonização, similares (Saldarriaga et al., 1988; Uhl et al., 1988; Moran et al., 1994; Salomão, 1994; Vieira, 1996; Tucker et al., 1998; Almeida, 2000; Steininger, 2000). Os poucos estudos com inventário contínuo, utilizando parcelas permanentes, se referem a estágios iniciais de sucessão vegetal após a abertura de clareiras em florestas maduras (Zwetsloot, 1981; Uhl e Jordan, 1984; Uhl, 1987). Conforme Pickett (1989), estudos que utilizam cronoseqüências e inventários contínuos são complementares, considerando que a cronoseqüência serve como fonte de informações descritivas para geração de hipóteses, as quais podem ser verificadas em comparação com informações obtidas nas parcelas permanentes. Os estudos de cronoseqüência são rápidos, de custo relativamente baixo, e expressam padrões de mudanças na estrutura e composição florísticas que ocorrem durante o desenvolvimento da floresta. Por outro lado, estudos com inventário contínuo revelam processos sucessionais, como mortalidade e recrutamento ao longo do tempo, que não são evidentes nos padrões observados na cronoseqüência, embora necessitem de mais tempo de estudo e apresentem um custo mais elevado. Assim, apesar de estudos de cronoseqüência dominarem a literatura e possuirem vantagens práticas óbvias, eles também têm limitações significativas, pois, mesmo quando as cronoseqüências são bem selecionadas, é impossível eliminar inteiramente o efeito de variações entre sítios, sobretudo, em relação ao prévio uso da terra. Além disso, a cronoseqüência assume que a variação do clima, entre os anos e tipos de distúrbio, têm impacto uniforme nos sítios de diferentes idades (Pickett, 1989). Na Amazônia brasileira, pesquisadores têm utilizado a cronoseqüência para descrever o desenvolvimento estrutural e florístico (Saldarriaga et al., 1988; Moran, et al., 1996; Vieira, 1996; Almeida, 2000; Steininger, 2000) e o efeito da intensidade do uso da terra sobre o acúmulo de biomassa vegetal (Uhl, 1987; Buschbacher et al., 1988; Uhl et al., 1988; Fearnside e Guimarães, 1996; Moran et al., 2000; Steininger, 2000). Esses estudos mostram que plantas invasoras e outras herbáceas, junto com espécies lenhosas pioneiras, dominam no início da sucessão. Após 10 a 20 anos, as espécies pioneiras envelhecem e morrem causando mudanças da dominância por espécies típicas de estágios mais avançados e aumentando gradualmente a diversidade da floresta sucessional. Estudos sobre a sucessão florestal, com base no inventário contínuo publicado na Amazônia, relatam a elevada ocorrência de espécies de floresta primária (Zwetsloot, 1981; Uhl e Jordan, 1984). Esses estudos indicam que somente os primeiros anos de sucessão são dinâmicos, com intensa invasão inicial de espécies herbáceas e um rápido aumento na densidade de indivíduos, conduzindo ao predomínio de poucas espécies pioneirais lenhosas de vida curta, as quais em poucos anos apresentam elevada mortalidade na área. As publicações com análises de dados em áreas em diferentes estágios de desenvolvimento, provenientes de inventário contínuo, são escassas. Este artigo enfatiza a complementaridade dos métodos de cronoseqüência e de inventário contínuo (sensu Pickett, 1989) no estudo do desenvolvimento de duas florestas sucessionais e adjacentes na Amazônia Oriental, uma inicialmente com 4 anos e a outra com 12 anos. No estudo são focadas três questões: a) Como as florestas de 4 e 12 anos diferem em relação à composição florística e estrutural de espécies arbóreas?; b) Como a mortalidade e sobrevivência de espécies arbóreas diferem nos dois sítios?; e c) De que modo o padrão sucessional inferido pela cronoseqüência concorda com os processos observados com a utilização de inventário contínuo? MATERIAL E MÉTODOS Área de estudo As áreas estudadas localizam-se na Estação Experimental de Castanhal, Universidade Federal Rural da Amazônia, próximo à cidade de Castanhal (1° 19´S, 47° 57´W), Pará. O solo está classificado como Latossolo Amarelo Distrófico – Fase Pedregosa I – Concrecionário Laterítico (Tenório et al., 1999). A área apresenta precipitação média anual de 2000 a 2500 mm, concentrada de dezembro a maio. A média anual da temperatura do ar varia entre 24 e 27oC (Martorano & Pereira, 1993). A paisagem da região é caracterizada por um mosaico de florestas em diferentes estágios de sucessão, áreas com cultura anual e pastagens degradadas. As áreas estudadas são florestas secundárias desenvolvidas partindo de vários ciclos de derruba e queima da vegetação em pousio e subseqüente cultivo de culturas anuais, o que caracteriza o sistema tradicional de agricultura de subsistência. Conforme relato de moradores locais, as áreas de estudo foram abandonadas após múltiplos ciclos de utilização com pousio, geralmente, de 7 a 10 anos, desde ~1940. As capoeiras estudadas são separadas por um ramal de 7 m de largura que conduz à sede da propriedade. A idade das florestas mencionadas no decorrer do trabalho será aquela referente ao início do estudo em 1999, isto é, “floresta de 12 anos” para a área abandonada em 1987 (~ 2 ha) e “floresta de 4 anos” para aquela abandonada em 1995 (~ 0,5 ha). A floresta de 4 anos apresenta em seu entorno capoeiras em diferentes estágios de sucessão (2 e 8 anos, no início do estudo); a floresta de 12 anos é adjacente a uma mata ciliar em um dos lados e florestas secundárias, em diferentes estágios de desenvolvimento nos demais lados. O primeiro inventário foi concluído em dezembro de 1999 na floresta de 12 anos e julho de 2000 na floresta de 4 anos. Posteriormente, os inventários continuaram sendo realizados anualmente, em julho de 2001, 2002 e 2003. Composição e estrutura A vegetação arbórea foi inventariada em quatro parcelas de 10 x 10 m nas florestas de 4 e 12 anos, onde foram medidos indivíduos de espécies arbóreas com diâmetro a 1,3 m de altura (DAP) ≥ 1 cm. Cada árvore medida foi identificada com plaqueta metálica numerada e identificada por botânico experiente. O material botânico foi coletado e herborizado no herbário da Embrapa Amazônia Oriental. Com o objetivo de avaliar as possíveis diferenças entre número de espécies, altura, diâmetro e área basal nas florestas estudadas, foi realizada uma comparação entre os valores médios por parcela, pela análise de variância com dois critérios de classificação (Two-way ANOVA), com medidas repetidas no tempo. A avaliação da densidade de indivíduos das três espécies mais comuns em ambos os sítios foi realizada por análise de variância com um critério de classificação (One-way ANOVA),com medidas repetidas no tempo. Quando necessário, os dados foram transformados em “ln” para que fossem apropriados ao teste. Foram também construídos histogramas de distribuição de diâmetro e altura para cada floresta. Recrutamento e mortalidade O inventário contínuo em 2001, 2002 e 2003, dos indivíduos com DAP ≥ 1 cm, permitiram o cálculo das taxas brutas de recrutamento e mortalidade, e taxa líquida de recrutamento para populações de espécies e para comunidade, conforme segue: a) Recrutamento Bruto = Rn+1/In b) Mortalidade Bruta = Mn+1/In c) Recrutamento líquido = (In+1-In)/In Em que: Rn+1 = número de novos indivíduos registrados no inventário do ano n + 1; In = número total de indivíduos registrados no inventário do ano “n”; Mn+1 =número de indivíduos presentes no inventário do ano “n” e registrados como mortos no inventário “n + 1”; In+1 = o número de indivíduos registrados no inventário do ano “n + 1”. RESULTADOSA riqueza de espécies arbóreas na floresta de 4 anos foi, aproximadamente, a metade da floresta de 12 anos (p < 0,05, Tabela 1). Durante o inventário contínuo, o número de espécies arbóreas observadas na floresta de 4 anos variou de 21 a 23 e na floresta de 12 anos de 48 a 51. TABELA 1: Densidade de indivíduos, diâmetro, altura, área basal e densidade de espécie (média ±SE) para árvores nas florestas de 4 e 12 anos, observadas em Castanhal, Pará, Brasil. Asteriscos indicam diferença significativa entre os sítios e letras indicam diferença significativa entre os anos em determinado sítio (P < 0,05; Análise de variância Two-way por medidas repetidas). TABLE 1: Density, diameter at breast height, height, species density and basal area (average ± SE) for overstory trees in 4-year-old and 12-year-old regrowth stands, measured in Castanhal, Pará, Brazil. Asterisks indicate significant differences between stands date, different letters indicate significant differences between years within a given stand (P < 0.05; Two-way repeated measure ANOVA).
Em relação à densidade de indivíduos em 100 m2, a diferença significativa, entre as florestas de 4 e 12 anos, ocorreu somente no primeiro ano de estudo (p < 0,05, Tabela 1). Na floresta de 4 anos, houve aumento significativo da densidade com o decorrer do processo de sucessão, tendendo a estabilização, enquanto na floresta de 12 anos, observou-se o processo contrário, com decréscimo do número de indivíduos ao longo do tempo (p < 0,05, Tabela 1). As espécies arbóreas mais abundantes na floresta de 4 anos foram Lacistema pubescens Mart., Vismia guianensis (Aubl.) Choisy, Banara guianensis Aubl. e Myrcia sylvatica (G. Mey.) DC., representando em média 83% da comunidade (Tabela 2). Na floresta de 12 anos, L. pubescens, M. sylvatica, V. guianensis e Lacistema aggregatum Berg Rusby, representaram em média 65% da comunidade (Tabela 2). As três espécies comuns e mais representativas em ambas as florestas (Lacistema pubescens, Vismia guianensis e Myrcia sylvatica) mostraram o mesmo comportamento em relação ao aumento da densidade de indivíduos na floresta de 4 anos e redução na floresta de 12 anos em todas as avaliações. No entanto, as duas primeiras espécies ficaram melhor representadas na floresta de 4 anos, enquanto Myrcia sylvatica se sobressaiu na floresta de 12 anos (Tabela 2). TABELA 2: Recrutamento e mortalidade total de árvores (DAP > 1 cm) em floresta de 4 anos estudada em julho de 2000 (00), 2001 (01), 2002 (02) e 2003 (03), e floresta de 12 anos estudada em novembro de 1999 (99), julho de 2001, 2002 e 2003, em Castanhal, Pará, Brasil. A área amostral foi 400 m2 em cada floresta. As espécies com abundância relativa menor do que 2% em qualquer um dos sítios foram agrupadas como “outras espécies”. TABLE 2: Recruitment and mortality for woody stems > 1 cm DBH in 400 m2 of 4-year-old regrowth stand at July 2000, 2001, 2002 and 2003; and 400 m2 of 12-year-old regrowth stand at November 1999, July 2001, 2002 and 2003, in Castanhal, Pará, Brazil. Species with relative abundances < 2 percent in both stands are grouped as “other species.”
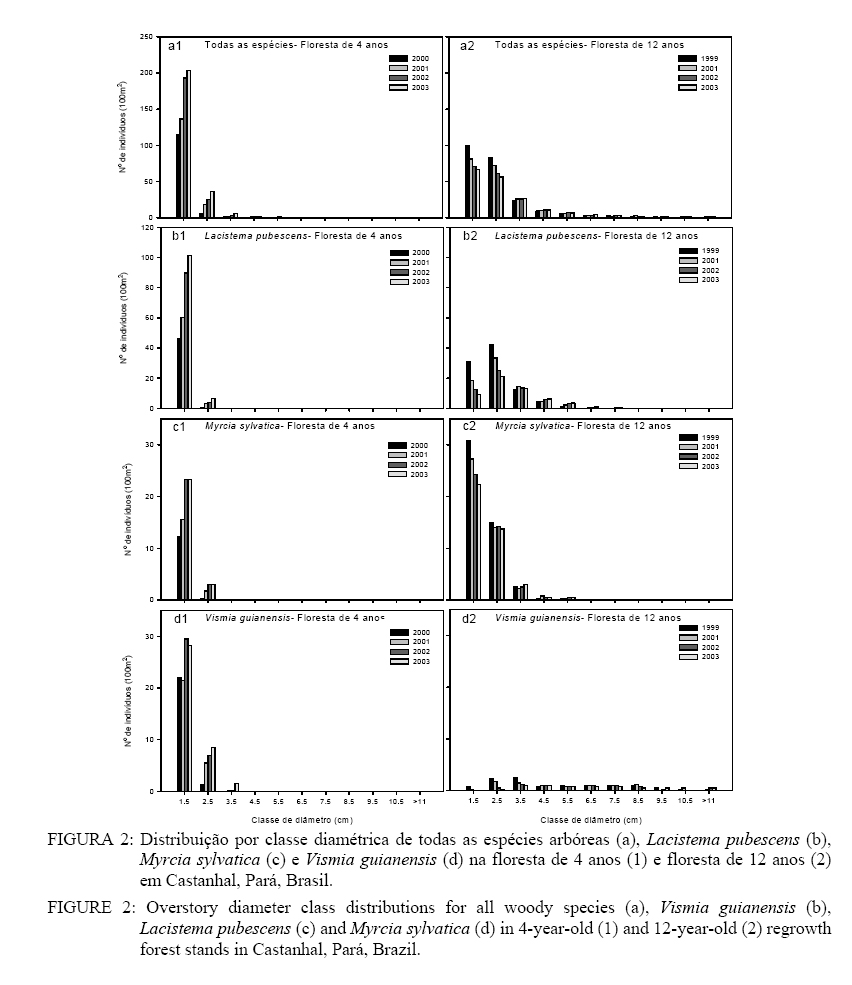
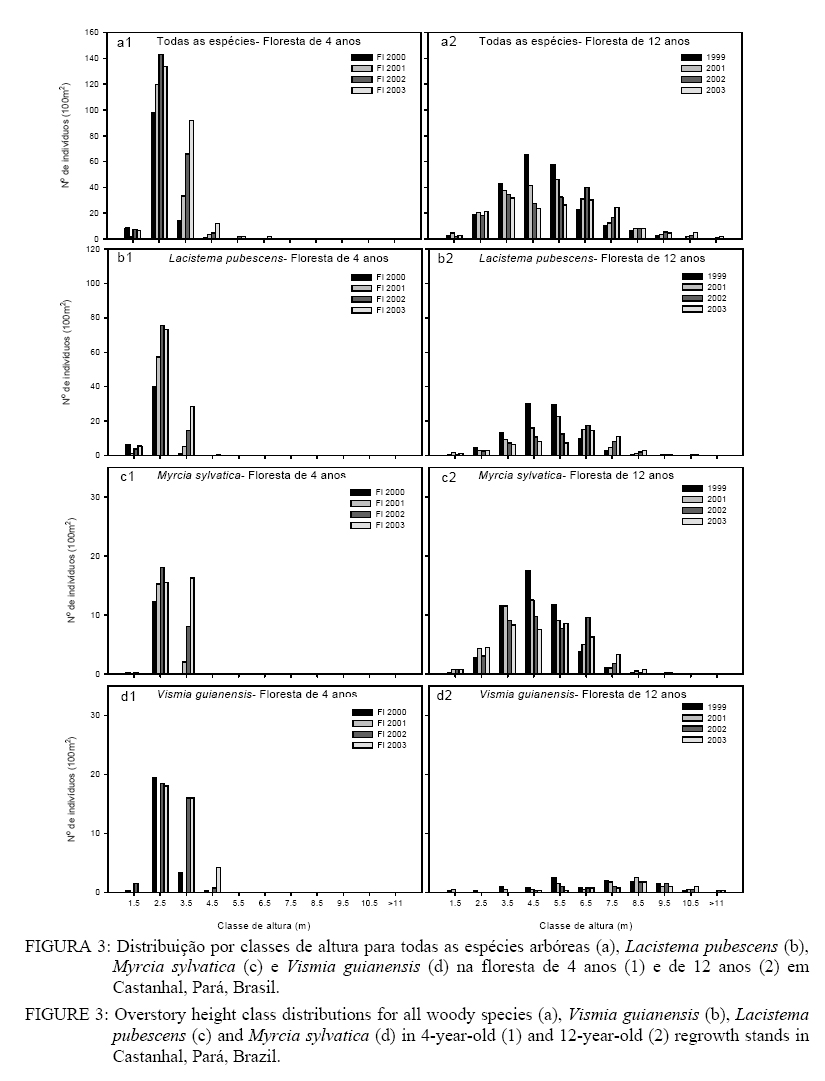
Lacistema pubescens foi a espécie que mais respondeu à sucessão, pois apresentou um aumento significativo na densidade de indivíduos na floresta de 4 anos e redução na floresta de 12 anos, enquanto Myrcia sylvatica e Vismia guianensis apresentaram mudanças menos intensas, em ambos sítios (p < 0,05, Figura 1). As médias de diâmetro, altura e área basal foram significativamente maiores na floresta de 12 anos em relação a floresta de 4 (p < 0,05, Tabela 1). A floresta do 4o ao 5o ano teve incremento significativo em diâmetro (20%) e área basal (90%), embora o incremento em altura tenha sido mais gradativo no decorrer da sucessão. Esse mesmo padrão foi observado na floresta de 12 anos (p < 0,05, Tabela 1). As médias de diâmetro e altura médios foram, respectivamente, 1,6 cm e 2,9 m na área de 4 anos e 2,9 cm e 5,5 m na de 12 anos. A distribuição das classes de diâmetro mostrou-se do tipo “J invertido” e normal nas classes de altura, em ambos os sítios (Figura 2a1 & a2 e Figura 3a1 & a2). No entanto, as três espécies mais comuns nos sítios (Lacistema pubescens, Myrcia sylvatica e Vismia guianensis) mostraram diferentes padrões de distribuição. Na floresta de 4 anos, Lacistema pubescens apresentou uma maior concentração de indivíduos com DAP entre 1 e 2 cm, enquanto na floresta de 12 anos a maior densidade ocorreu na classe de 2 a 3 cm; quanto à altura, a forma da curva foi semelhante nos dois sítios, embora na floresta de estágio mais avançado ocorra distribuída num maior número de classes (Figura 2b1 & b2 e Figura 3b1 & b2). Em ambas as florestas, a maioria dos indivíduos de Myrcia sylvatica apresentou DAP entre 1 e 2 cm. Em termos de altura, Myrcia sylvatica apresentou a maioria dos indivíduos entre 2 e 4 m, na floresta de 4 anos, atingindo alturas maiores na floresta de 12 anos(Figura 1c1, 2c2 e 3c1, 3c2). Na floresta de 4 anos, Vismia guianensis ficou melhor representada por indivíduos com DAP e altura entre 1 e 3 cm, e 3 e 5 m de altura respectivamente. Na floresta de 12 anos, Vismia guianensis ocorreu em maior número de classes de DAP e altura. O recrutamento líquido de indivíduos na floresta de 4 anos nos dois primeiros períodos de estudo, considerando 2000-2001 e 2001-2002, aumentaram 30 e 41% respectivamente. No terceiro período (2002 a 2003), o recrutamento líquido decresceu para 11% (Tabela 2). Esta tendência foi observada em todas as parcelas (Tabela 3). A maioria do recrutamento foi por causa das três espécies mais comuns em ambos os sítios (Lacistema pubescens, Vismia guianenesis e Myrcia sylvatica), além de Banara guianensis, as quais apresentaram tendência similar à da comunidade, isto é, de maior recrutamento entre 2000-2002 e menor entre 2002-2003. Abarema jupumba não apresentou recrutamento, e sim 12% e 13% de mortalidade, no segundo e terceiro períodos respectivamente. Rollinia exsucca (DC. Ex Dunal) A. DC.e Lacistema aggregatum apresentaram recrutamento líquido positivo, enquanto, Casearia arborea (Rich.) Urb. apresentou um recrutamento líquido de 73% apenas entre 2001 e 2002; e recrutamento e mortalidade igual nos outros períodos. A maioria das espécies manteve ou aumentou o número de indivíduos e duas espécies novas ingressaram em 2001 e 2002, respectivamente, Connarus angustifolius (Radlkofer) G. Schellend. e Annona paludosa Aubl. TABELA 3: Recrutamento e mortalidade de árvores em parcelas de 100 m2 na floresta de 4 anos (FS 4) estudada em julho de 2000 (00), 2001 (01), 2002 (02) e 2003 (03), e de 12 anos (FS 12) estudada em novembro de 1999 (99), julho de 2001, 2002 e 2003, em Castanhal, Pará, Brasil. TABLE 3: Recruitment and mortality of stems ≥ 1 cm DBH per 100 m2 plot in the 4-year-old regrowth stand (FS 4) between July 2000 - July 2003; and the 12-year-old regrowth stand (FS 12) between November 1999 - July 2003 in Castanhal, Pará, Brazil.
Na floresta de 12 anos, a densidade de indivíduos declinou 10, 9 e 4% de novembro de 1999 a julho de 2001, julho de 2001 a julho de 2002 e deste até julho de 2003, respectivamente (Tabela 2). Da mesma forma, todas as parcelas mostraram maior mortalidade do que recrutamento (Tabela 3). As três espécies mais representativas na floresta de 12 anos (Lacistema pubescens, Myrcia sylvatica e Vismia guianensis)apresentaram maior mortalidade do que recrutamento. Lacistema aggregatum manteve-se, aparentemente, estável no ecossistema, apesar de apresentar recrutamento líquido positivo entre 2001 e 2002 (entre 13 e 14 anos). A maioria dos recrutamentos ocorreu com espécies menos comuns no sítio, como Tabernaemontana angulata Mart. ex. Müll. Arg.e Heisteria sp., além de Lacistema aggregatum e Cupania scrobiculata L.C.Rich.,que também tiveram indivíduos ingressados (Tabela 4). Rollinia exsucca apresentaou elevada mortalidade. TABELA 4: Espécies lenhosas de florestas secundárias de 4 anos inventariadas (400 m2) em julho de 2000 (00), 2001 (01), 2002 (02) e 2003 (03) e de 12 anos inventariadas (400 m2) em novembro de 1999 (99), julho de 2001, 2002 e 2003, em Castanhal, Pará, Brasil. TABLE 4: Woody species inventory for 4-year-old secondary stand (400 m2), study at July 2000, 2001, 2002 and 2003 and 12-year-old (400 m2), study at November 1999, July 2001, 2002 and 2003, in Castanhal, Pará, Brazil.
TABELA 4: Continuação ... TABLE 4: Continued …
DISCUSSÃO Na Amazônia, Uhl et al. (1988), Vieira (1996) e Peña-Claros (2003) observaram que na sucessão inicial da floresta poucas espécies predominam no ambiente. Entretando, o reduzido número de espécies e a alta densidade de indivíduos na floresta de 12 anos não são comuns quando comparadas com estudos de Vieira (1996) e Almeida (2000) em florestas com idade semelhante na região, embora, Saldarriaga et al. (1988) tenham verificado densidade de indivíduos quatro vezes menor do que foi observado neste estudo. A elevada dominância de espécies na floresta de 12 anos pode ser explicada pela intensidade de uso da terra, a qual se caracterizou por longos e repetidos ciclos de cultivo, durante cerca de 60 anos. Provavelmente, esse intenso uso da terra tenha resultado em esgotamento do banco de sementes do solo e redução da chuva de sementes na área. Sob essas condições, a rebrotação tem uma importante função no restabelecimento dessas florestas na região (Vieira, 1996), portanto, a dominância de Lacistema pubescens, Vismia guianensis, Myrcia sylvatica e Banara guianensis observada neste estudo pode ser atribuída à elevada capacidade de rebrotação dessas espécies. Depois de estabelecidas, tais espécies pioneiras, provavelmente, mantêm sua população por reproduzirem cedo, concordando com os resultados obtidos por Salomão (1994). A baixa riqueza florística na floresta de 4 anos durante todo o período de estudo, quando comparada com a floresta de 12 anos, foi uma incógnita, conduzindo à dúvida se seria possível, no intervalo não-estudado (8 a 11 anos), que o número de espécie aumentasse mais do que o dobro. De acordo com Jacquemyn et al. (2003) o aumento da riqueza de espécies em fragmentos ocorre de acordo com seu grau de isolamento e, apesar da floresta de 4 anos apresentar florestas secundárias no seu entorno, estas também estão em estágio inicial de sucessão, tendo sido submetidas a vários ciclos de cultivo e possível perda de diversidade. Uma hipótese a ser testada é se a floresta ripária e madura, adjacente à floresta de 12 anos, contribuiu para o aumento do número de espécies nesse ecossistema. Em termos de estrutura, a variação no limite mínimo de diâmetro das árvores amostradas, nos diferentes inventários realizados na Amazônia, dificulta uma análise comparativa dos estudos. Assim, para facilitar a comparação de estudos similares com o presente, foi feita uma separação dos indivíduos com DAP ≥ 5 cm inventariados neste estudo. Na floresta de 4 anos, as árvores com DAP ³ 5 cm só foram observadas a partir do 6o ano de sucessão, indicando crescimento relativamente menor quando comparado com outros sítios de semelhante idade (Salomão, 1994; Vieira, 1996; Mesquita et al., 2001). Na floresta de 12 anos, foram observados 12 indivíduos/100 m2 com DAP ³ 5 cm em 1999, aumentando para 20 em 2003; tanto a densidade quanto a área basal foram baixas quando comparadas a outros sítios, também de semelhante idade (Salomão, 1994; Vieira, 1996, Mesquita et al., 2001), provavelmente, em conseqüência da baixa fertilidade do solo associada ao intenso uso da terra. Algumas novas espécies (DAP ≥ 1 cm), presentes na floresta de 12 anos e ausentes na floresta de 4 anos como, por exemplo, Cupania scrobiculata, Guatteria poeppigiana Mart. e Inga rubiginosa, são tolerantes à sombra e, conseqüentemente, apresentam um estabelecimento mais tardio. Espécies de floresta madura tanto de terra firme, como de ambiente com maior influência do lençol freático, também estão presentes no sítio de 12 anos, incluindo Trattinickia burserifolia e Vochysia inundata (Almeida, 2000). Embora as espécies pioneiras continuem dominando a vegetação, a presença de muitas espécies tardias na floresta de 12 anos indica uma fase de transição do desenvolvimento sucessional. Outras cronoseqüências sugerem semelhante transição (Saldarriaga et al. 1988; Almeida 2000; Vieira 1996) e são consistentes com o modelo de tolerância sucessional (Connell e Slatyer 1977), onde grupos de espécies se misturam à medida que o desenvolvimento da floresta ocorre, causando uma mudança gradual na dominância e eventual substituição de espécies. A exclusão de Banara guianensis, Casearia arborea e Abarema jupumba na floresta de 12 anostambémevidencia o processo de sucessão florestal, onde novas espécies mais tolerantes à sombra passam a competir com as pioneiras e substituí-las. A Myrcia sylvatica, por exemplo, é uma espécie mais tolerante à sombra que dominou o sub-bosque em ambos os sítios, enquanto a densidade de Vismia guianensis e Banara guianensis (pioneiras intolerantes à sombra) reduziu consideravelmente no sub-bosque do sítio de 12 anos. Com o processo sucessional, Vismia guianensis e Banara guianensis são substituídas por espécies mais tolerantes à sombra como Ocotea costulata, Myrcia bracteata e Miconia ciliata (Lima, 2003; Coelho et al., 2002). Em adição aos padrões sucessionais evidentes nos dados do sub-bosque, a distribuição da população de indivíduos das espécies comuns no dossel, nas florestas de 4 e 12 anos, também revela uma trajetória distinta (Figura 1 e 3). A distribuição do diâmetro de Vismia guianensis, Lacistema pubescens e Myrcia sylvatica na floresta de 4 anosfoi quase-idêntica. No entanto, a redução da densidade de indivíduos de Vismia guianensis na floresta de 12 anos, em comparação com a de 4 anos, concorda com alguns trabalhos que relatam que Vismia spp., normalmente, persiste 10 a 20 anos após abandono de área agrícolas em outros sítios na Amazônia Oriental (Salomão, 1994) e no Alto Rio Negro (Uhl, 1987). Quando comparado à floresta de 4 anos, a redução da densidade de indivíduos de Lacistema pubescens na classe de diâmetro pequeno, na floresta de 12 anos, reflete no reduzido recrutamento para os estratos superiores, sugerindo que essa espécie, no decorrer da sucessão, apresentará comportamento similar ao de Vismia guianensis. Em contraste com o comportamento de Lacistema pubescens e Vismia guianensis, a concentração de indivíduos de Myrcia sylvatica na classe de tamanho menor, em ambas as florestas, indica o lento crescimento, porém, numa população mais persistente no processo sucessional. Lacistema pubescens, Vismia guianensis e Myrcia sylvatica são responsáveis pela maioria das mudanças sucessionais em ambos os sítios, porém os padrões de recrutamento e mortalidade nos dois ambientes revelam dois diferentes estágios de desenvolvimento, cada um sujeito a diferente dinâmica. A floresta de 4 anos é um exemplo de sítio inicial dominado por espécies pioneiras, enquanto que a floresta de 12 anos representa um estágio de exclusão de indivíduos estabelecidos e inclusão de novas espécies mais tolerantes nas classes de menor diâmetro (sensu Oliver e Larson 1996). Os anos iniciais de sucessão secundária na Amazônia são caracterizados por agressivo crescimento da floresta. A elevada disponibilidade de luz durante os anos iniciais de sucessão secundária favorece a germinação, brotação e crescimento de espécies pioneiras (Uhl e Jordan, 1984; Saldarriaga et al., 1988). Na floresta de 4 anos, o recrutamento líquido de 30, 41 e 11% nos períodos de 2000 a 2001, 2001 a 2002 e 2002 a 2003 respectivamente demonstra a colonização de espécies pioneiras de crescimento agressivo. As quatro espécies mais comuns no sítio, Lacistema pubescens, Vismia guianensis, Banara guianensis e Myrcia sylvatica, representaram em média 83% de todos indivíduos recrutados e 86% de todos indivíduos mortos na floresta de 4 anos durante os 36 meses de mensuração. Na floresta de 12 anos quase-todas as espécies comuns às duas florestas reduziram sua densidade e aumentaram a porcentagem de mortalidade durante quase todo o período de avaliação, indicando o declínio gradual das pioneiras dominantes (Tabela 2). Esse declínio, provavelmente ocorreu por causa da senescência natural e condições de sombra estabelecida que diminuiram a capacidade de recrutamento (Whitmore, 1990). As três espécies mais comuns no sítio (Lacistema pubescens, Myrcia sylvatica e Vismia guianensis) representaram em média 85% da mortalidade no período de estudo somente 36% dos indivíduos recrutados. As taxas de mortalidade (6-13%) na floresta de 12 anos são superiores àquelas normalmente registradas para florestas tropicais úmidas em desenvolvimento (1 a 2%) (Condit et al. 1995; Swaine et al., 1987). No entanto, Condit et al. (1995) também registraram mortalidade anual de espécies colonizadoras superior a 10% na floresta do Barro Colorado. Em San Carlo do Rio Negro, Uhl e Jordan (1984) registraram mortalidade em massa de Cecropia ficifolia Warb. ex Snethl., depois de dois anos de crescimento. A comparação da estrutura e composição florísticas das florestas de 4 e 12 anos, tanto na cronoseqüencia como no inventário contínuo, revelou dois sítios dominados por espécies semelhantes. Entre as espécies presentes na floresta de 4 e 12 anos, 46% e 36% respectivamente estão presentes em sítios de idade semelhante na região estudados por Vieira (1996). Quando utilizamos apenas a cronoseqüência não é possível avaliar a real performance das espécies nos ecossistemas como, por exemplo, quando estas começam a ser excluídas, além disso se as florestas de 4 e 12 anos fossem comparadas partindo de dados de um único inventário, acreditar-se-ia que a riqueza florística aumentaria gradativamente, ao longo do processo sucessional e mais do que dobraria o número de espécies aos 12 anos de idade. O inventário contínuo induziu à dúvida se isso irá acontecer por causa da falta de mudanças até o 7o ano. Nesse sentido, o inventário contínuo é que definiu melhor quando a substituição iniciou e as novas espécies apareceram, complementando o entendimento da auto-ecologia das espécies secundárias. Por outro lado, se a análise for limitada ao inventário contínuo em apenas um dos sítios, os dados de mortalidade e recrutamento não revelam as diferenças de composição e estrutura florística entre os dois sítios. Os resultados deste estudo indicam que os métodos de cronoseqüência e o inventário contínuo fortalecem o entendimento do desenvolvimento sucessional em florestas quando analisados conjuntamente. No entanto, quando utilizados separadamente, cada abordagem tem sua desvantagem com relação a onerar os custos, requerer mais tempo de avaliação e não perceber ou interpretar incorretamente informações importantes. Quando combinadas, uma abordagem compensa a deficiência da outra. CONCLUSÕES As espécies predominantes foram as mesmas nos sítios de 4 e 12 anos. Estruturalmente, o autodesbaste inicia próximo aos 12 anos, com redução da densidade. A floresta de 4 anos representa o estágio de iniciação da floresta, com elevado recrutamento líquido de espécies predominantes; o processo de exclusão predomina na floresta de 12 anos, em conseqüência da elevada mortalidade. Os padrões e processos sucessionais são melhores descritos quando se utilizam, conjuntamente, os métodos de cronoseqüência e de inventário contínuo. AGRADECIMENTOS Os autores agradecem a colaboração de Raimundo Nonato da Silva pelo suporte logístico; Alexandre Modesto, Cesar Tenório, Gilson da Silva Sousa, Glebson A. da Silva Sousa, Osório L. Oliveira, Evandro Rodrigues da Silva e Elisângela Santos pela assistência no trabalho de campo. Esta pesquisa foi apoiada pela Estação Experimental de Agricultura da Flórida e financiada pela Fundação Andrew Mellon. REFERÊNCIAS BIBLIOGRÁFICAS
Copyright 2005 - Ciência Florestal The following images related to this document are available:Photo images[cf05034f2.jpg] [cf05034f3.jpg] [cf05034f1.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}