 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ciência Florestal, Vol. 16, No. 1, 2006, pp. 27-43 CURVAS DE ÍNDICE DE SÍTIO PARA POVOAMENTOS CLONAIS DE Eucalyptus saligna Smith PARA A DEPRESSÃO CENTRAL E SERRA DO SUDESTE, RIO GRANDE DO SUL SITE INDEX CURVES FOR CLONAL STANDS OF Eucalyptus saligna Smith FOR THE CENTRAL DEPRESSION AND SOUTHEASTERN MOUNTAIN RANGE, RIO GRANDE DO SUL Helio Tonini1 Paulo Renato Schneider2 Cesar Augusto Guimarães Finger3 1. Engenheiro Florestal, Dr., Pesquisador do CPAF Embrapa Roraima, Br 174, Km 08, Distrito industrial, CEP:
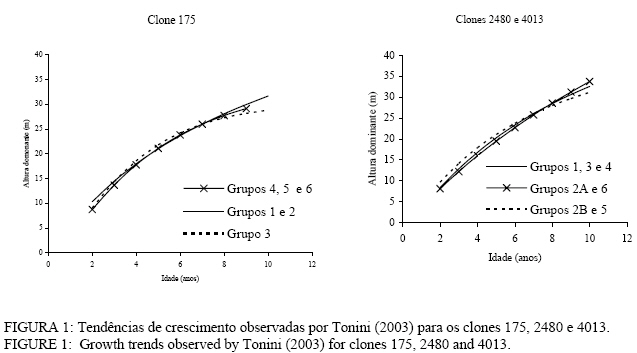
69301-970, Boa Vista (RO). helio@cpafrr.embrapa.br Recebido para publicação em 4/09/2003 e aceito em 11/02/2006. Code Number: cf06004 RESUMO Este trabalho foi desenvolvido com o objetivo de estudar o crescimento em altura dominante de clones de Eucalyptus saligna Smith e construir curvas de índice de sítio fazendo uma comparação com curvas construídas para povoamentos da mesma espécie originados de mudas por sementes. A análise das tendências de crescimento em altura dominante indicaram a necessidade de se construir três sistemas de curvas de índices de sítio monomórficas em função do clone e do tipo de solo. A comparação com as curvas de índice de sítio existentes no mesmo local de estudo, para povoamentos de Eucalyptus saligna originados de mudas por sementes indicou que elas, de maneira geral, não poderiam ser utilizadas para os clones estudados. Palavras-chave: Eucalyptus saligna; clone; índice de sítio. ABSTRACT This work was an attempt to study the growth in dominant height of Eucalyptus saligna Smith clones, to build site index curves and to compare them with curves built for stands of the same species originated from seeds. The dominant height growth trends analysis indicated the necessity of constructing three monomorfic site index curves systems according to clone and soil type. The comparison with the site index curves obtained from stands of the same species originated from seeds available for the same locality indicated that they could not be used for the clones studied. Key words: Eucalyptus saligna; clone; site index. INTRODUÇÃONo Brasil, até o princípio do século XX, o eucalipto foi plantado como árvore decorativa. Desde então, em razão da Companhia Paulista de Estradas de Ferro, surgiram as primeiras plantações com fins comerciais, e o eucalipto passou a ser cada vez mais utilizado como combustível em forma de lenha e carvão. Somente a partir de 1966, com a instituição de um programa nacional de incentivos fiscais, o eucalipto passou a ser plantado em grande escala. Assim, a área plantada saltou rapidamente de 400 mil hectares, existentes antes de 1966, para os atuais 3 milhões de hectares. Atualmente, o setor de papel e celulose tem indiscutível importância nos contextos econômico e social brasileiro, levando o País à condição de maior produtor mundial de celulose de fibra curta, onde as condições ambientais favoráveis associada à evolução nas técnicas de silvicultura, manejo e melhoramento genético fazem com que as florestas brasileiras estejam entre as mais produtivas do mundo. A necessidade de obter florestas cada vez mais produtivas, com madeira de alta densidade, baixos teores de extrativos e outras características que se correlacionam positivamente com a qualidade do produto final, tem levado as empresas brasileiras a investimentos cada vez maiores em programas de melhoramento genético, com a utilização de técnicas como a hibridação e a clonagem. A clonagem é considerada, mundialmente, como a maneira mais eficiente de se produzir madeira em qualidade e quantidade exigidas pelo mercado. Possibilita a produção em massa de madeira com características previamente selecionadas e assegura maior rendimento no processo de produção em todas as suas etapas. Além disso, pode-se obter ganhos expressivos na qualidade do produto final por causa da maior homogeneidade da matéria-prima. No entanto, para prover uma fábrica com essa matéria-prima, é necessário que as florestas proporcionem um suprimento contínuo, que só é possível com o conhecimento da dinâmica do crescimento dos povoamentos florestais. O estudo de crescimento e produção, por analisar a relação entre uma medida de produção florestal em relação a variáveis como a idade, o sítio e a densidade, é a ferramenta mais utilizada para a compreensão dessa dinâmica. Nos modelos de crescimento e produção, a capacidade produtiva do sítio florestal está relacionada à curva de crescimento em altura dominante, que por sua vez, dá origem às curvas de índice de sítio. Com a introdução de materiais genéticos superiores, existe a necessidade de se verificar se as curvas de índice de sítio, construídas para povoamentos originados de mudas por sementes, podem ser utilizadas para clones. Portanto, uma vez que a clonagem é uma realidade com tendência irreversível, existindo áreas cada vez maiores plantadas com esse tipo de material genético, estudos de crescimento e produção para clones de eucalipto tornam-se cada vez mais necessários. Assim, este trabalho foi desenvolvido com o objetivo de construir curvas de índice de sítio para três clones de Eucalyptus saligna e fazer uma comparação com curvas construídas para povoamentos da mesma espécie originados de mudas por sementes. REVISÃO BIBLIOGRÁFICA Clutter et al. (1983) afirmaram que os métodos para predizer o crescimento de florestas são baseados no conceito de que o crescimento e a produção de povoamentos de uma determinada espécie, ou composição de espécies, são influenciados pela idade do povoamento ou a distribuição da idade em povoamentos inequiâneos; pela capacidade inata de produção da área envolvida; pela suposição de que a capacidade inata de produção do sítio tem sido e é plenamente utilizada pelos tratamentos silviculturais aplicados. A definição de sítio dada pela Sociedade Americana de Engenheiros Florestais citada por Davis (1966), considera-o como a capacidade de produzir florestas ou outra vegetação, sob a combinação de condições biológicas, climáticas e edáficas. Para Spurr (1952), a qualidade do sítio é a soma total dos fatores edáficos, biológicos e climáticos que afetam as plantas, em que o sítio não é um fator, nem todos os fatores, mas a soma dos fatores efetivos entre os quais um ou mais são dominantes. A produtividade de uma área florestal, definida em termos da qualidade do sítio, representa uma medida da quantidade máxima de madeira que a área pode produzir em um determinado momento. Em um determinado macroclima, a qualidade do sítio determina o tipo e a magnitude de riscos e oportunidades que se tem ao manejar um povoamento, sendo que nenhuma decisão silvicultural é válida sem referência à qualidade do sítio (Daniel et al., 1979). A altura dominante é a variável dendrométrica que melhor se enquadra aos requisitos propostos por Burger (1976), para determinar a qualidade do sitio florestal, que são: facilidade de determinação a campo; boa correlação com a produção volumétrica; indica a qualidade do sítio para todas as idades do povoamento e independencia da densidade do povoamento. O índice mais utilizado para determinar a produtividade de uma área florestal é o índice de sítio, que é uma expressão da qualidade do sítio baseada na altura das árvores dominantes e codominantes em uma determinada idade índice (Carmean, 1970). Segundo Clutter et al. (1983), os dados para o desenvolvimento de curvas de índice de sítio são obtidos basicamente de três fontes: medições em parcelas temporárias, medições em parcelas permanentes e reconstrução da relação altura/idade para árvores individuais por meio de análise de tronco. Entre as três fontes, as parcelas permanentes oferecem os melhores dados para o desenvolvimento de curvas de índice de sítio. No entanto, a obtenção dos dados dessa forma é mais dispendiosa, e estes poderão ser utilizados somente após várias remedições periódicas. A análise de tronco proporciona dados com, praticamente, a mesma precisão dos obtidos por parcelas permanentes, com a vantagem de que podem ser utilizados imediatamente. Autores como Johnston et al. (1967), Daniel et al. (1979), Barros et al.(1986), Avery e Burkhart (1994) e Way e Klinka (1996) fazem algumas críticas ao método do índice de sítio. Johnston et al. (1967) afirmaram que a correlação entre o crescimento em volume e o crescimento em altura não é perfeita e citam o trabalho de Mader em 1963 que verificou que o crescimento em altura apresentou melhor correlação com fatores de sítio diferentes dos que melhor se correlacionaram com crescimento volumétrico. Daniel et al. (1979) afirmaram que o índice de sítio mede indiretamente a produtividade em termos de volume de madeira, o que restringe o conceito de qualidade do sítio, pois esta, não é correlacionada com a produção de outros produtos florestais. A densidade e a supressão podem influenciar o índice de sítio, e a introdução de árvores geneticamente superiores torna obsoleta a determinação da qualidade do sítio baseada no índice de sítio pré-existente. Para Barros et al. (1986), o poder de predição da produtividade do índice de sítio é bastante baixo, por ser um método desenvolvido e aplicado inicialmente em regiões onde os sítios apresentavam maior potencial para a manutenção da produtividade, onde o manejo florestal não é tão intensivo quanto no Brasil com solos de baixa fertilidade natural, e onde uma porção considerável dos nutrientes está imobilizada na biomassa. Avery e Burkhart (1994) afirmaram que o índice de sítio não é uma constante, mudando periodicamente em conseqüência de variações ambientais e climáticas. O seu valor para uma determinada espécie não pode ser utilizado para diferentes espécies no mesmo sítio. Outras limitações descritas pelos autores são a dificuldade em se determinar a idade exata de um povoamento, a sua impropriedade para florestas naturais, a não consideração dos efeitos da densidade do povoamento pela seleção de árvores em povoamentos bem estocados que não tenham sido afetados pela supressão passada e o fato de não serem consideradas outras variáveis associadas ao volume do povoamento como o DAP e a forma do tronco. Para Way e Klinka (1996), em conseqüência da compensação ecológica, em que diferentes combinações de fatores ambientais podem resultar em efeitos similares no crescimento das árvores, alguns problemas no uso de curvas de índice de sítio podem ser encontrados. Uma determinada espécie pode ter o mesmo índice de sítio em locais úmidos e ricos em nutrientes e em locais secos e pobres em nutrientes, sendo que esses dois sítios podem necessitar de diferentes práticas de manejo com o objetivo de produzir madeira de grandes dimensões. No entanto, segundo Cunha Neto et al.(1994), apesar de algumas limitações, o índice de sítio é o método mais utilizado para definir a potencialidade dos sítios florestais e assim o será até que métodos que conjuguem a soma de vários fatores ambientais possam ser traduzidos de forma numérica, acessível aos usuários do setor. MATERIAL E MÉTODO Localização da área e caracterização dos clonesEste estudo foi realizado em plantios clonais de Eucalyptus saligna, pertencentes à empresa Klabin Riocell S.A, localizada em Guaíba, a 20 km de Porto Alegre, no Rio Grande do Sul. A Klabin Riocell S.A possui 39.554 hectares reflorestados com o gênero Eucalyptus, distribuídos por 24 municípios, localizados nas regiões da Depressão Central e Serra do Sudeste no estado do Rio Grande do Sul. A partir da década de 70, a empresa começou a investir em um grande programa de melhoramento genético, com a seleção de matrizes e estacas, sendo que atualmente possui 8.381 hectares plantados com clones e híbridos. A área plantada com os clones de Eucayptus saligna é de 4.325,8 hectares, totalizando 10,9% da área total. As matrizes tiveram origem em sementes coletadas na Austrália, que foram levadas para Itatinga, SP onde foram semeadas e selecionadas as melhores árvores, principalmente em relação ao crescimento e à qualidade do fuste. Essas árvores foram abatidas e suas brotações originaram as estacas que deram origem aos clones. Características edafoclimáticas Os hortos florestais onde foram coletados os dados abrangem duas regiões fisiográficas distintas: Depressão Central e Encosta do Sudeste. Conforme a classificação de Köppen, o clima geral é caracterizado como do tipo Cfa, ou seja, mesotérmico subtropical, com verões quentes e sem estação seca. Segundo Nimer (1990), na Encosta do Sudeste, a precipitação média anual é de 1500 mm, com máxima precipitação nos meses de julho, agosto e setembro. Na Depressão Central, a precipitação média anual é de 1.750 mm, com máxima precipitação nos meses de abril, maio e junho. Conforme a classificação existente de solos para as áreas da empresa, os solos mais comumente encontrados na região da Encosta do Sudeste são os Neossolos e os Cambissolos e, na Depressão Central os Nitossolos e Argissolos, também ocorrendo Gleissolos e Planossolos, comumente encontrados próximo ao litoral. Segundo Hermann e Rosa (1990), a região da Encosta do Sudeste localiza-se sobre a unidade de relevo denominada de Planalto Sul-Rio-Grandense que se caracteriza por apresentar forma triangular, com altitude ao redor dos 400 m e relevo fortemente ondulado. A Depressão Central faz parte das depressões periféricas da bacia do Rio Paraná e se constitui em uma área sem grandes variações altimétricas, com as maiores cotas situando-se aos redor dos 200 m. Os dados Os dados utilizados neste estudo são provenientes de 608 parcelas permanentes de Eucalyptus saligna distribuídas em 29 hortos florestais distribuídas da seguinte forma: 251 Parcelas plantadas com o clone 2480, 95 parcelas com o clone 4013 e 262 parcelas com o clone 175, todos em primeira rotação com um mínimo de uma e um máximo de quatro medições. A Tabela 1, mostra a distribuição dessas unidades por densidade inicial de plantio, bem como os valores máximos e mínimos observados para idade, diâmetro médio e altura dominante. O processo amostral utilizado pela empresa é a amostragem aleatória estratificada, na qual os estratos são formados pelo horto florestal, espécie, rotação, ano de plantio ou corte e espaçamento. As unidades amostrais utilizadas apresentavam forma retangular com as dimensões de 24 x 18 m (432 m2) e 27 x 18 m (486 m2), sendo a última utilizada em áreas com espaçamentos de 2,7 m entre linhas. No interior de cada unidade amostral, foram medidos o diâmetro à altura do peito de cada árvore, a altura das dez primeiras árvores e a altura das quatro árvores mais grossas, seguindo o conceito de altura dominante definido por Assmann. Construção das curvas de índice de sítio Como base para a construção das curvas de índice de sítio, utilizaram-se as análises de tendências de crescimento em altura dominante feitas por Tonini (2003) e Tonini et al (2004). Nesse estudo, foram identificadas seis tendências distintas na curva de crescimento em altura dominante para os clones estudados, que variaram no seu padrão de crescimento em relação ao fator solo. Pela análise de covariância baseando-se na não existência de diferenças significativas de inclinação, foram identificadas seis tedências de crescimento, sendo três para o clone 175, e três para os clones 2480 e 4013. Para o clone 175, a primeira tendência foi observada para clones crescendo em Nitossolos com horizonte B nítico e Argissolos com B textural, ambos não-hidromórficos (Grupos 1 e 2); a segunda para clones crescendo em cambissolos (Grupo 3); e a terceira para clones crescendo em Planossolos com horizonte B plânico, hidromórficos, Neossolos litólicos e Neossolos quartzarênicos (Grupos 4, 5 e 6) TABELA 1: Distribuição das unidades amostrais por clone e densidade inicial de plantio e valores máximos e mínimos observados para idade (t) em anos; diâmetro médio (d) em centímetros e altura dominante (h100 ) em metros. TABLE 1: Plots distribution per clone, initial density and observed maximum and minimum values for age (t) in years, average diameter (d) in centimeters and dominant height (h100) in meters.
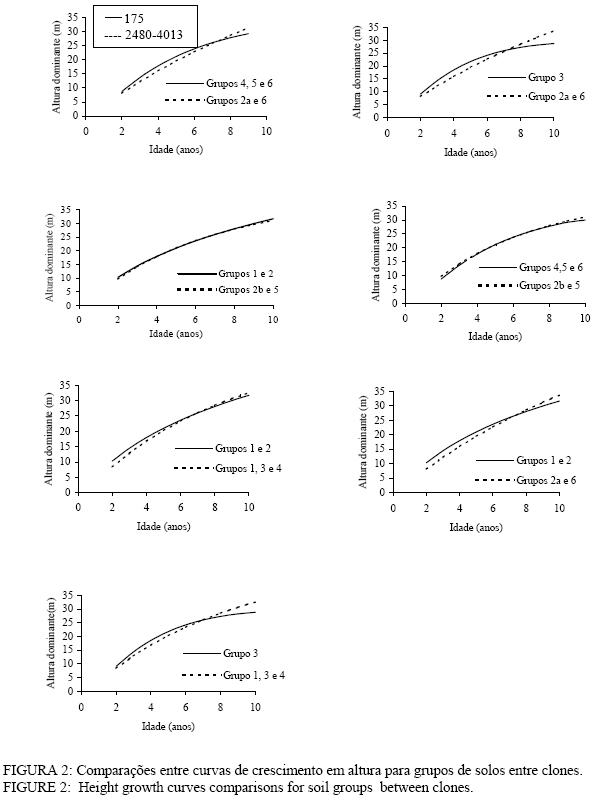
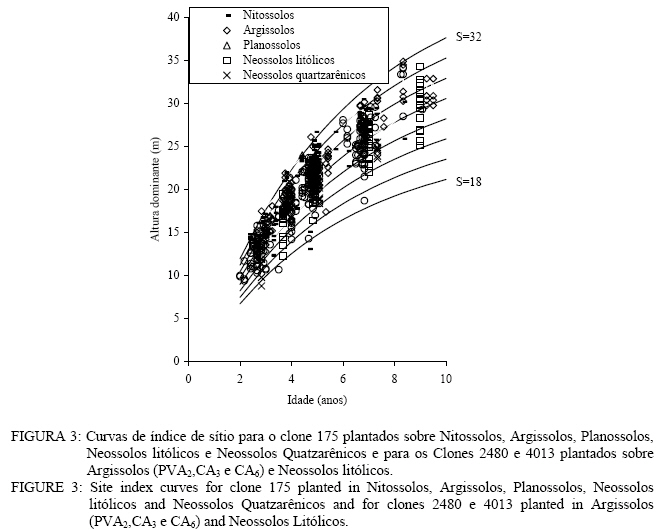
Para os clones 2480 e 4013, as tendências observadas foram: clones crescendo em Nitossolos com horizonte B nítico, não-hidromórficos, Cambissolos com horizonte B câmbico não-hidromórficos e Planossolos hidromórficos com horizonte B plânico (grupos 1, 3 e 4); Clones crescendo em Argissolos com horizonte B textural, e Neossolos quatzarênicos, ambos não hidromórficos (Grupos 2a e 6); e clones crescendo em Neossolos litólicos não-hidromórficos e alguns tipos de Argissolos com horizonte B textural (grupos 2b e 5). Maiores detalhes sobre a formação dos grupos e os resultados obtidos com a análise de covariância, podem ser observados em Tonini (2003) e Tonini et al. (2004). Neste estudo, somente serão apresentadas as análises gráficas que comprovaram os resultados obtidos na análise de covariância. A altura dominante foi estimada partindo do modelo de Bakmann que apresentou bom ajuste para todos os clones analisados (Tonini, 2003 e Tonini et al. 2004), sendo expressa por: 1nh100 =b0 + b11nt + b2in2t Em que: h100 = altura dominante (m); t = idade (anos); b0 = constante da equação; b1 e b2 = coeficientes da equação; ln = logaritmo nepariano. Para determinar a produtividade das áreas florestais, foram construídas curvas de índice de sítio monomórficas que expressam a qualidade do sítio florestal baseadas nas alturas das árvores dominantes em uma determinada idade índice. Como metodologia, empregou-se o método da curva mestre ou guia (Schneider, 1993). Idade índice e sistema de classificação adotado A idade índice, segundo Schneider (1993), é a idade-padrão, escolhida arbitrariamente, na qual se compara a altura dominante dos diferentes sítios. A escolha dessa idade está na dependência da rotação da espécie e deve ser preferencialmente fixada no final da rotação. Neste estudo, optou-se por uma idade de referência de 7 anos, por ser esta, a idade de rotação média para Eucalyptus no Brasil, manejados em densidade completa. Em razão do sistema absoluto de classificação de sítios ser mais ilustrativo e o mais utilizado em classificações brasileiras (Schneider, 1993), optou-se pelo seu emprego. Determinação do número de curvas necessárias para cobrir a variação de sítios As curvas de índice de sítio devem cobrir toda a variação entre o menor e o maior crescimento em altura dominante observado na região. Para a delimitação das curvas de índice de sítio, os pares de dados altura-idade, medidos nas unidades amostrais permanentes, foram plotados juntamente com as curvas de índice de sítio. O número ideal de curvas foi definido à medida que as curvas de índice de sítio, construídas com um intervalo de 2 m, abrangeram toda essa variação. Confiabilidade das curvas de índice de sítio A estabilidade dos índices de sítio, ao longo do desenvolvimento do povoamento florestal, é essencial para a acuracidade na aplicação dessas curvas em períodos sucessivos e essa estabilidade foi testada comparando-se as alturas reais com as curvas de índice de sítio, conforme utilizado por Machado (1980), Scolforo e Machado (1988) e Scolforo (1992). Segundo Clutter (1984), o desejável é que a altura média das árvores dominantes da unidade amostral permaneça em uma mesma classe de sítio durante toda a sua vida, propiciando, dessa forma, uma forte base para estudos de crescimento e produção. Para testar a confiabilidade e a estabilidade na construção das curvas, foram selecionadas aleatoriamente 125 unidades amostrais com, no mínimo, três medições. Comparações com curvas de índice de sítio para povoamentos de Eucalyptus saligna originados de mudas por sementesAs comparações entre as curvas de índice de sítio, desenvolvidas neste estudo, e as curvas para povoamentos de Eucalyptus saligna, originados de mudas por sementes, foram feitas com base nos trabalhos de Finger (1991 e 1997). O autor construiu dois sistemas de curvas de índice de sítio para Eucalyptus grandis e Eucalyptus saligna, contendo nove curvas para povoamentos plantados sobre solos profundos, com preparo de solo profundo, e plantados sobre solos rasos, com preparo de solo superficial, com uma idade índice de 7 anos, na mesma região deste estudo. Os valores estimados por Finger (1991 e 1997) foram comparados aos valores gerados por este estudo, de forma gráfica. RESULTADOS E DISCUSSÃO Construção das curvas de índice de sítio Uma vez que no estudo de Tonini (2003) e Tonini et al. (2004), foi verificada a existência de diferentes padrões de crescimento entre clones e dentro de um mesmo clone em função do grupo de solo, tornou-se necessário definir o número de sistemas de curvas de índice de sítio necessárias para representar os padrões de crescimento em altura dominante observados. Na Figura 1, observam-se diferenças na tendência de crescimento em altura entre os grupos de solo para cada clone. Essas diferenças foram estatisticamente comprovadas pela análise de covariância por Tonini (2003) para 1% de probabilidade de confiança. No entanto, as diferenças de inclinação detectadas pela análise de covariância entre os grupos 4, 5 e 6 e 1 e 2, no clone 175, e entre os grupos 1, 3 e 4 e 2a e 6, nos clones 2480 e 4013, podem ser consideradas pequenas, (± 1 m) ficando inclusive dentro dos erros cometidos ao se medir alturas à campo. Já para os grupos 3 e 2B e 5, existe uma leve superioridade inicial no crescimento em altura (± 2 m), que não permanece à medida que a idade avança. A comparação gráfica entre as curvas de crescimento em altura para grupos de solo de mesma tendência de crescimento entre clones pode ser observada na Figura 2. Observa-se na Figura 2, que a diferença de inclinação detectada pela análise de covariância (Tonini, 2003), para a curva de crescimento da altura dominante entre clones e para o clone 175 em relação aos demais pode ser constatada graficamente, uma vez que as curvas para os clones 2480 e 4013 se apresentam mais inclinadas na maioria das comparações.No entanto, tem-se um mesmo padrão de crescimento em altura dominante para os grupos 4, 5 e 6; 1 e 2 e 2b e 5, indicando que os clones 2480 e 4013, crescendo sobre Neossolos Litólicos e alguns tipos de Argissolos, apresentam um padrão semelhante ao clone 175 crescendo sobre Nitossolos, Argissolos, Planossolos Hidromórficos, Neossolos Litólicos e Neossolos Quartzarênicos. Nesse caso, um único sistema de curvas de índice de sítio pode ser construído para representar o crescimento desses clones e grupos de solo. Pode-se concluir que as diferenças de inclinação detectadas pela análise de covariância (Tonini, 2003 e Tonini et al, 2004), indicaram de forma precisa a existência de diferentes padrões de crescimento entre os clones e dentro de um mesmo clone, e que as diferenças de crescimento em altura dominante se devem ao fato desses clones crescerem de maneira diferenciada na grande maioria dos solos analisados e não por apresentarem diferenças em um tipo de solo específico. Essas diferenças no padrão de crescimento em altura indicaram a necessidade de se construir três sistemas de curvas de índice de sítio, sendo dois sistemas para o clone 175, uma vez que o padrão de crescimento em altura dominante para árvores crescendo em Cambissolos foi bastante diferenciado em relação aos demais. O maior achatamento na curva de crescimento em altura dominante para o clone 175, plantado sobre cambissolos, concorda com os resultados obtidos por Rezende (1994). O autor afirma que esses solos geralmente estão em posição mais íngreme, são rasos, armazenando menos e perdendo mais água, constituindo sistemas muito problemáticos para a conservação e o manejo, por causa da pobreza de nutrientes, da declividade e da susceptibilidade à erosão. As curvas de índice de sítio definitivas foram construídas reajustando a função de Backmann para os grupos de solo 1, 2, 4, 5 e 6, no clone 175, e 2b e 5, nos clones 2480 e 4013; para o grupo de solo 3, no clone 175, e para os grupos de solo 1, 2a, 3, 4 e 6 nos clones 2480 e 4013 cujos valores estimados podem ser observados nas Tabelas 2, 3 e 4 e o número de curvas necessárias com os seus respectivos valores, nas Figuras 3, 4 e 5. TABELA 2: Altura dominante estimada para os índices de sítio do clone 175 plantados sobre Nitossolos, Argissolos, Planossolos, Neossolos litólicos e Neossolos quartzarênicos e dos clones 2480 e 4013 plantados sobre Argissolos (PVA2, CA3 e CA6). TABLE 2: Predicted dominant height for site index of clone 175 planted in Nitossolos, Argissolos, Planossolos, Neossolos litólicos and Neossolos quatzarênicos and of Clones 2480 and 4013 planted in Argissolos (PVA2, CA3 and CA6).
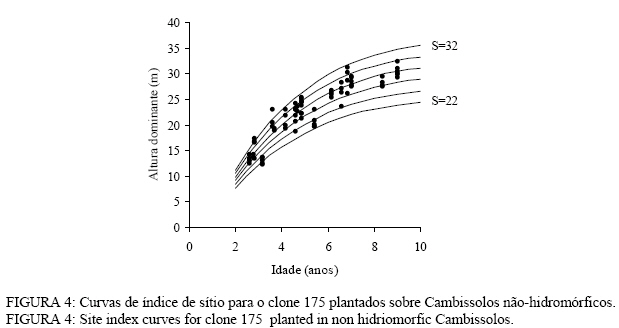
TABELA 3: Altura dominante estimada para os índices de sítio do clone 175 plantados sobre Cambissolos não-hidromórficos, na região da Encosta do Sudeste. TABLE 3: Predicted dominant height for site index of clone 175, planted in non hidromorfic Cambissolos.
TABELA 4: Altura dominante estimada para os índices de sítio dos clones 2480-4013 plantados sobre Nitossolos, Argissolos (PVA1 e PVA3) Planossolos, Cambissolos e Neossolos quartzarênicos nas regiões da Serra do Sudeste e Litoral. TABLE 4: Predicted dominant height for site index of clones 2480 and 4013 planted in Nitossolos, Argissolos (PVA1, and PVA3), Planossolos, Cambisolos and Neossolos Quartzarenicos.
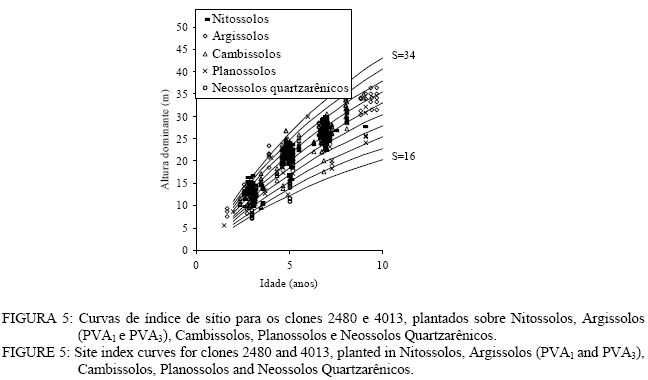
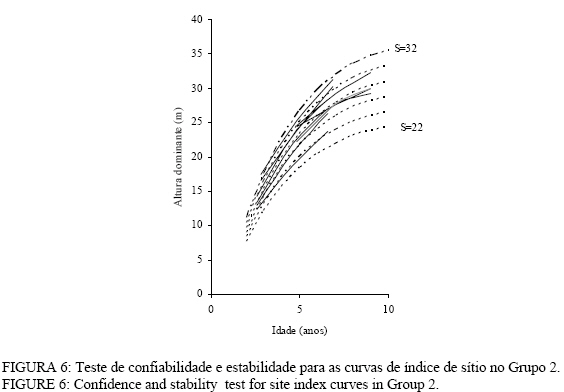
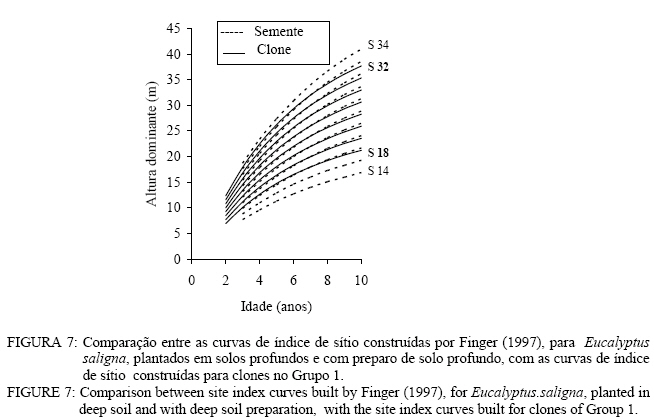
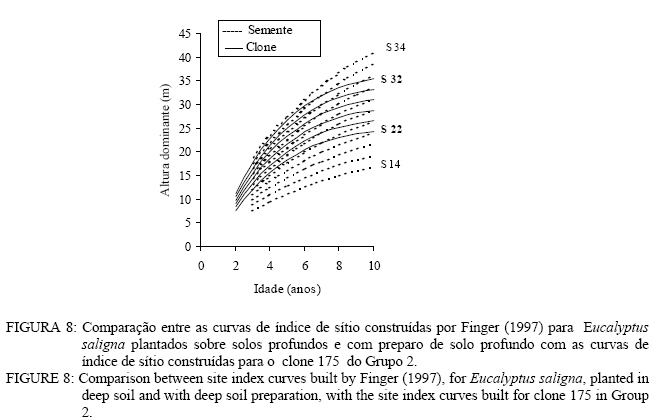
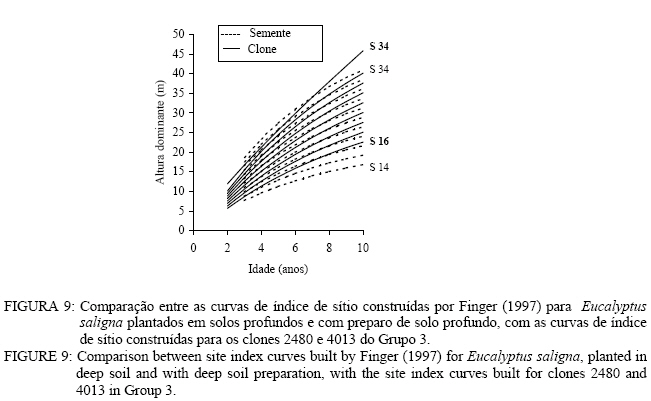
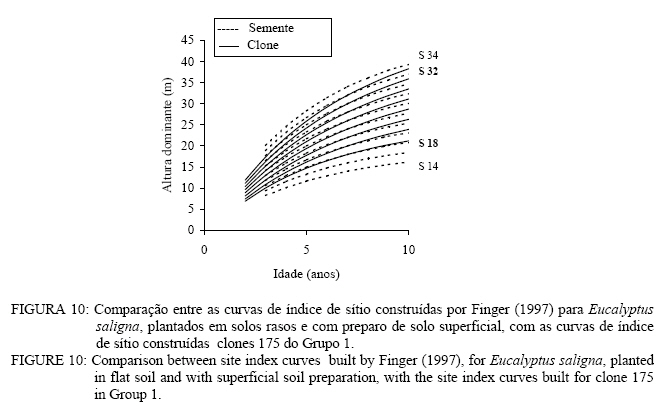
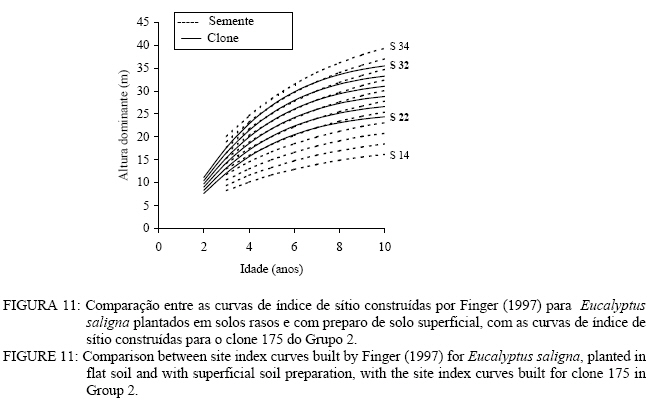
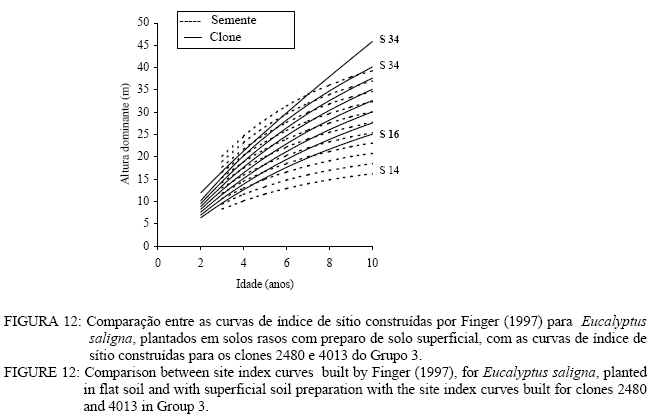
Observa-se, na Figura 3, que foram necessárias oito curvas de índice de sítio, com intervalo de 2 metros, para abranger toda a variação em altura dominante observada. Para esses clones, os piores índices de sítio estão associados aos Nitossolos litólicos e Nitossolos quartzarênicos, sendo observadas, no melhor índice de sítio unidades amostrais crescendo sobre Nitossolos e Argissolos. Observa-se, na Figura 4, que, para o clone 175 crescendo sobre Cambissolos, o número de unidades amostrais permanentes disponíveis para a realização deste estudo foi bem menor, sendo necessária a construção de um sistema com seis curvas de índice de sítio, para cobrir a variação em altura dominante observada. Observa-se, na Figura 5, que, para os clones 2480 e 4013, foi necessária a construção de um sistema de curvas de índice de sítio com dez curvas distribuídas em um intervalo de 2 metros. Os piores índices de sítio foram observados em unidades amostrais localizadas sobre Nitossolos quartzarênicos, Planossolos e Cambissolos, e os melhores nos Nitossolos e Argissolos . Confiabilidade e estabilidade das curvas de índice de sítio O teste de confiabilidade e estabilidade das curvas de índice de sítio, realizado partindo da plotagem das alturas reais sobre as curvas de índice de sítio, permitiu avaliar graficamente o comportamento das árvores quanto a uma eventual mudança de índice de sítio ao longo do tempo, bem como avaliar a confiabilidade de se efetuar classificações em idades mais jovens, conforme pode ser observado na Figura 6. Pela análise da Figura 6, verifica-se boa estabilidade das curvas de índice de sítio, no grupo 2, no qual a altura dominante observada, na grande maioria das unidades amostrais, permaneceu estável (sem tendência a mudar de curva). Isso indica que tendem a permanecer em um mesmo índice de sítio, o que é desejável para os estudos de crescimento e produção. Resultado semelhante foi obtido para os demais grupos. De forma geral, observou-se maior instabilidade até a idade de 4 anos, indicando que a utilização dessas curvas para povoamentos muito jovens pode trazer alguma imprecisão na classificação de sítios. Essa instabilidade inicial é provavelmente ocasionada pela adaptação da planta ao local de crescimento nas idades mais jovens e tem sido mencionada em vários estudos que procuraram classificar o sítio mediante o crescimento em altura dominante como Scolforo e Machado (1988), Marcolin (1990) e Selle (1993). Para verificar a independência das curvas de índice de sítio em relação à idade, as alturas estimadas pelas curvas de índice de sítio foram plotadas sobre as idades de 3, 4, 5, 7 e 9 anos, em que não foi observada nenhum tipo de tendência. A relação índice de sítio (IS) e idade (t) representada pela função: IS = b0+b1t apresentou valor de correlação simples de 0,09, não-significativo a 1% de probabilidade. Comparação com curvas de índice de sítio para Eucalyptus saligna originados de mudas por sementesA comparação entre as curvas construídas para os clones 175 e 4013-2480 com as curvas construídas por Finger (1997) para Eucalyptus saligna, para solos profundos com preparo de solo profundo e para solos rasos com preparo de solo raso podem ser observadas nas Figuras 7, 8, 9, 10, 11 e 12. As Figuras 7, 8, 9, 10, 11 e 12 mostram que o número de curvas de índice de sítio necessárias para abranger a variação do crescimento em altura para povoamentos oriundos de mudas por sementes é maior que os clonais, o que reflete a maior heterogeneidade desses povoamentos. A análise das Figuras 7, 8 e 9, que compararam o sistema de curvas de índice de sítio construído por Finger (1997) para povoamentos de Eucalyptus saligna plantados em solos profundos com preparo de solo profundo, com os três sistemas construídos neste estudo, mostra que o primeiro sistema (Figura 7) apresenta mesma tendência de crescimento nas idades iniciais, porém, com tendência a ser mais achatado à medida que a idade avança. O segundo sistema (Figura 8) apresentou tendência bastante diferenciada em relação ao desenvolvido por Finger (1997), ocorrendo cruzamentos entre estes. O sistema desenvolvido para Eucalyptus saligna originados de mudas de sementes, apresenta-se nitidamente mais inclinado, atingindo, em um mesmo índice de sítio, maiores alturas sobretudo a partir dos sete anos. Esse resultado confirma a hipótese de que, sob certas circunstâncias, a produção pode ser maior em um povoamento geneticamente diverso do que em um uniforme, pois um mesmo genótipo, ao demandar pelos mesmos fatores de produção ao mesmo tempo, utiliza o sítio de maneira menos eficiente do que uma mistura de genótipos. Na comparação com o terceiro sistema (Figura 9), ocorreu o inverso. O sistema construído para povoamentos de Eucalyptus saligna originados de mudas de sementes, plantados sobre solos profundos, mostra-se bem mais achatado, ocorrendo novamente cruzamentos entre as curvas de índice de sítio. Nas Figuras 10, 11 e 12, podem-se observar as comparações entre os três sistemas desenvolvidos neste estudo com os construídos por Finger (1997) para povoamentos de Eucalyptus saligna originados de mudas de sementes plantados sobre solos rasos com preparo superficial. Na Figura 10, observa-se que o primeiro sistema se comporta novamente com tendência semelhante nas idades iniciais. No entanto, ao contrário da comparação anterior, mostrou-se levemente mais inclinado nas maiores idades. O segundo sistema (Figura 11) apresentou semelhante tendência inicial de crescimento em altura dominante, indicando estar mais próximo do padrão de crescimento apresentado pelo povoamento de Eucalyptus saligna originado de mudas de sementes crescendo em solos rasos do que pelos povoamentos crescendo em solos profundos. No entanto, nas maiores idades mostrou maior achatamento. O terceiro sistema (Figura 12), da mesma forma que na comparação anterior, mostrou-se bem mais inclinado, com um padrão de crescimento em altura dominante bastante diferenciado em relação ao Eucalyptus saligna originado de mudas de sementes, crescendo em solos rasos. Essas comparações mostraram que a construção de curvas de índice de sítio, específicas para esses clones, foi necessária pois, apresentaram padrão de crescimento diferenciado em relação aos povoamentos de Eucalyptus saligna originados de mudas por sementes. CONCLUSÕES Tomando por base a realização deste trabalho, pode-se concluir que: a) A existência de diferentes padrões de crescimento em altura dominante entre clones e grupos de solo tornou necessária a construção de três sistemas de curvas de índice de sítio, sendo dois sistemas para o clone 175, uma vez que o padrão de crescimento em altura dominante para árvores crescendo em Cambissolos foi bastante diferenciado em relação aos demais. b) A comparação entre as curvas de índice de sítio, construídas para povoamentos de Eucalyptus saligna originados de mudas por sementes e clones, indicou que as curvas de índice de sítio construídas por Finger (1997), para povoamentos originados de mudas de sementes, dependendo do clone e grupo de solo, não poderiam ser utilizadas, pois haveria grande diferença no padrão do crescimento em altura dominante entre esses povoamentos. REFERÊNCIAS BIBLIOGRÁFICAS
Copyright 2006 - Ciência Florestal
The following images related to this document are available:Photo images[cf06004f3.jpg] [cf06004f5.jpg] [cf06004t1.jpg] [cf06004f2.jpg] [cf06004f8.jpg] [cf06004f11.jpg] [cf06004f10.jpg] [cf06004t3.jpg] [cf06004f6.jpg] [cf06004f1.jpg] [cf06004f12.jpg] [cf06004f9.jpg] [cf06004t4.jpg] [cf06004f7.jpg] [cf06004f4.jpg] [cf06004t2.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}