 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ciência Florestal, Santa Maria, Vol. 16, Núm. 2, 2006, pp. 163-175 APORTE E DECOMPOSIÇÃO DE SERAPILHEIRA EM ÁREAS DE FLORESTA SECUNDÁRIA, PLANTIO DE SABIÁ (Mimosa caesalpiniaefolia Benth.)E ANDIROBA(Carapa guianensis Aubl.) NA FLONA MÁRIO XAVIER, RJ LITTER PRODUCTION AND DECOMPOSITION IN AREAS OF SECONDARY FOREST, PLANTATION OF SABIÁ (Mimosa caesalpiniaefolia Benth.) AND PLANTATION OF ANDIROBA (Carapa guianenses Aubl.) IN FLONA MÁRIO XAVIER, RJ Milton Marques Fernandes1 Marcos Gervasio Pereira2 Luis Mauro Sampaio Magalhães3 Adriano Rosa Cruz4 Rômulo Guimarães Giácomo4 1Engenheiro Florestal, Doutorando em Agronomia – Ciência do Solo, Universidade Federal Rural do Rio de Janeiro,
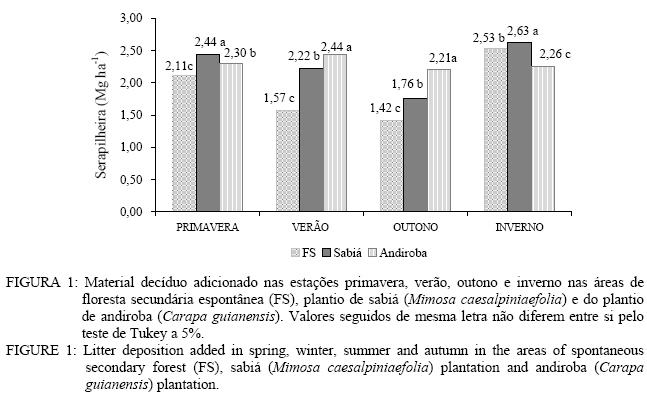
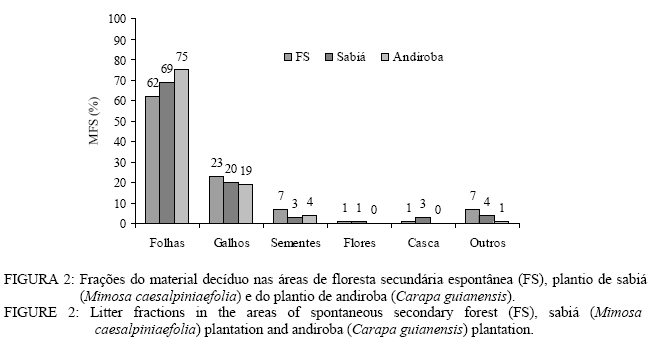
CEP 23890-000, Seropédica (RJ). milmarfer@bol.com.br Code Number: cf06016 RESUMO Este trabalho teve como objetivo avaliar o aporte e a decomposição da serapilheira em áreas de floresta secundária espontânea, plantio de sabiá (Mimosa caesalpiniaefolia Benth.) e plantio de andiroba (Carapa guianensis Aubl.). Para avaliação do aporte de serapilheira, foram instalados dez coletores cônicos sendo o aporte avaliado durante o período de outubro de 2003 a setembro de 2004. Posteirormente, foi feita a quantificação dos macronutrientes (N, P e K). A avaliação da taxa de decomposição foi realizada utilizando-se o método dos litter bags. A área de plantio de andiroba aportou a maior quantidade de serrapilheira 9,20 Mg ha-1,sendo seguida pelo plantio de sabiá com 9,06 Mg ha-1 e pela floresta secundária espontânea 7,63 Mg ha-1 por ano. A serapilheira das áreas de plantio de sabiá e do plantio de andiroba apresentou maiores valores de nitrogênio e fósforo que a área de floresta secundária. A velocidade de decomposição da serapilheira foi maior nas áreas de plantio de sabiá e andiroba, podendo tal comportamento ter sido influenciado pelo maior conteúdo de nitrogênio na serapilheira. O fósforo foi o nutriente que apresentou maior velocidade de liberação. Palavras-chave: serapilheira; deposição; decomposição; macronutrientes. ABSTRACT This study evaluated litter production and decomposition in areas of spontaneous secondary forest, sabiá (Mimosa caesalpiniaefolia Benth.) and andiroba (Carapa guianensis Aubl.) plantation. For litter contribution evaluation, 10 conical collectors were installed, and the contribution was evaluated from October, 2003 to September, 2004. N, P and K contents were quantified in litter. To evaluate decomposition, rate litter bags method was used. In the andiroba plantation area, it was observed the higher litter amount 9,20 Mg ha-1, followed by the sabia plantation with 9,06 Mg ha-1 and spontaneous secondary forest 7,63 Mg ha-1. Sabiá and andiroba litter presented larger nitrogen and phosphorus values in relation to the secondary forest area. Litter decomposition rate was faster in sabiá and andiroba areas. This behavior might be influenced by larger nitrogen content in the litter. Phosphorus was the nutrient that presented the higher liberation rate. Keywords: litter; litter fall; decomposition; nutrients. INTRODUÇÃO O estudo do fluxo dos nutrientes no ecossistema, isto é, a produção e decomposição da serapilheira com a conseqüente transferência desses para o ambiente, é essencial para a caracterização dos padrões de ciclagem, pois representa a principal via de retorno de nutrientes e matéria orgânica à superfície do solo (Pagano e Durigan, 2000). A produção de serapilheira e a devolução de nutrientes em ecossistemas florestais constituem a via mais importante no sistema solo-planta. Caracteriza-se, no primeiro estádio, pela absorção de nutrientes pelas raízes e por sua distribuição pelas diferentes partes da planta, sendo a taxa de absorção maior no período em que as árvores se encontram em estádio juvenil, o que corresponde ao período de maior produtividade dentro do processo de sucessão (Kimmins, 1993). Após esse período, os nutrientes são transferidos novamente para o solo, pela deposição de serapilheira, lixiviação pelas folhas, ramos e troncos e pela ação da chuva, além do trabalho da fauna herbívora e da dispersão de frutos e sementes (Poggiani e Schumacher, 2000). Vários fatores bióticos e abióticos influenciam a produção de serapilheira, tais como: tipo de vegetação, altitude, latitude, precipitação, temperatura, regimes de luminosidade, relevo, deciduosidade, estágio sucessional, disponibilidade hídrica e características do solo. Dependendo das características de cada ecossistema, um determinado fator pode prevalecer sobre os demais (Figueiredo Filho et al., 2003). No solo, ocorre rápida decomposição inicial de material lábil e, posteriormente, num processo mais lento, de materiais mais resistentes. Essa lenta decomposição pode ser em conseqüência do mecanismo de adsorção, à estabilização de metabólitos e à queda da taxa de biomassa no solo. Enfim, a biodegradação é um processo complexo e multifacetado, envolvendo grande número e variedade de microorganismos do solo. A degradação de diferentes resíduos depende das condições locais e regionais como clima, tipo de solo, vegetação, fauna e microorganismos decompositores (Tauk, 1990). Nos solos altamente intemperizados, assim como nos degradados, a serapilheira constitui-se na maior fonte de matéria orgânica, sua quantidade e natureza desempenham importante papel na formação e manutenção da fertilidade desses e, conseqüentemente, de nutrientes para a flora e fauna do solo degradado. Dada a importância da serapilheira na auto-sustentabilidade dos ecossistemas florestais, muitos pesquisadores têm conduzido trabalhos sobre a produção e decomposição dessa e o retorno de nutrientes ao solo (Gisler, 1995). Os ecossistemas de florestas tropicais, via de regra, apresentam produção contínua de serapilheira no decorrer do ano, sendo que a quantidade produzida nas diferentes épocas depende do tipo de vegetação considerada (Leitão-Filho,1993). Áreas mais alteradas apresentam um número muito elevado de espécies pioneiras que têm um crescimento rápido, ciclo de vida mais curto, investindo intensamente na produção de biomassa em um curto espaço de tempo (Martins e Rodrigues, 1999). Por outro lado, as áreas menos modificadas possuem um pequeno número de espécies pioneiras, apresentando menor produção de biomassa (Leitão-Filho,1993). Dessa forma, diferenças na produção de serapilheira entre trechos próximos podem estar relacionadas aos diferentes graus de perturbação que são encontrados dentro de um mesmo tipo florestal (Werneck et al., 2001). No entanto, dependendo do tipo de manejo formações iniciais podem apresentar alta produção de serapilheira. Existem vários trabalhos sobre a produção e decomposição da serapilheira com áreas reflorestadas com espécies florestais, mas a maioria dos trabalhos avalia esses plantios nos seus primeiros anos sem que se possa observar como eles se comportam após a sua estabilização. Portanto, é importante avaliar o comportamento de espécies florestais decorrido um longo período de tempo após a sua implantação. Este trabalho teve como objetivo avaliar a produção e decomposição da serrapilheira em áreas de floresta secundária, plantio de Mimosa caesalpiniaefolia (sabiá) e plantio de Carapa guianensis (andiroba) ambos com posterior regeneração natural. MATERIAL E MÉTODOS Meio Físico O estudo foi realizado no município de Seropédica, RJ, localizado na latitude 22° 46’ de latitude sul e 43° 41’ de longitude oeste e situada a 33 m de altitude. O clima da região de estudo é classificado como Aw de Köppen. A temperatura média máxima é de 25,9°C em dezembro e a mínima de 20,6°C em julho e agosto. A precipitação média e de 1369 mm/ano com déficit hídrico de julho a setembro excedente hídrico de dezembro a março. A história de ocupação humana em Seropédica onde vários ciclos agrícolas se alternaram, aliado à prática da pecuária extensiva, quase nada deixou da cobertura florestal primitiva. Com exceção das áreas de encosta serrana, praticamente não existem áreas contínuas com floresta nativa, dentro dos limites do município. Atualmente, fragmentos de florestas secundárias ocorrem de maneira esparsa. Um dos maiores e mais significativos encontra-se na Floresta Nacional Mário Xavier, graças à proteção dada pela transformação da área em uma unidade de conservação de uso direto do tipo floresta nacional, em 1945. Para o estudo foram selecionadas três áreas floresta de sucessão secundária espontânea, plantio de sabiá (Mimosa caesalpiniaefolia) com posterior regeneração natural e plantio de andiroba (Carapa guianensis) com posterior regeneração natural. Em cada uma das áreas de estudo foi delimitado um talhão de 0,1 hectare. O solo nas três áreas de estudo se enquadra na ordem dos Planossolos Háplicos situados na mesma condição de relevo plano a suave ondulado. Métodos Material decíduo O aporte de serapilheira foi avaliado, mensalmente, pelo uso de 10 coletores, dispostos acima da superfície do solo, com formato circular com 0,15 m2 de área (perímetro igual a 1,40 m) numa área delimitada de 0,1 hectare para cada tratamento. Os coletores foram distribuídos aleatoriamente dentro de cada área sendo fixados em árvores a aproximadamente 1 metro de altura do solo. A produção de serapilheira foi estimada segundo (Lopes et al., 2002) partindo da seguinte fórmula: PAS = (∑ PS x 10.000) / Ac Em que: PAS = Produção média anual de serapilheira (kg ha-1 ano-1); PS = Produção média mensal de serapilheira (kg ha-1 mês-1); Ac = Área do coletor (m2). Os coletores foram instalados no início da primavera no dia 21 de setembro de 2003. As coletas foram realizadas mensalmente até setembro de 2004. O material coletado foi seco em estufa a 65º C, sendo posteriormente separado em frações (folhas, galhos < a 2 cm diâmetro, sementes, flores, casca e outros correspondendo a fragmentos < 2 mm de diâmetro não identificáveis e pesado. Uma subamostra do material foi moída em moinho tipo Willey para a realização da análise química que foi realizada por meio de digestão sulfúrica (Tedesco et al., 1985), sendo determinados nitrogênio (N) pelo método de destilação de arraste a vapor (Bremer e Mulvaney, 1965), o fósforo (P) por colorimetria e potássio (K) por fotometria de chama. Todos os resultados foram transformados para conteúdo de nutrientes. Decomposição da serapilheira A avaliação da taxa de decomposição foi feita pela análise de perda de massa utilizando-se litter bags. Os litter bags consistiam em sacolas de polivinil com malha de 4 mm e dimensões de 25 x 25 cm e 1,5 cm de altura. Em cada litter bag foram adicionadas 10 gramas de folhas do material aportado nos coletores cônicos após esse ter sido seco em estufa a 65°C ate peso constante. Os “litter bags” foram distribuídos aleatoriamente na superfície do piso florestal, simulando a queda natural do material formador da serapilheira. Os litter bags foram instalados no inicio do verão (21/12/03) e coletados aos 30, 60, 90 e 120 dias. Após a coleta, os litter bags foram secos em estufa de circulação de ar forçada (65°C ± 5o) até atingir peso constante. Em seguida, foram levados ao laboratório onde o seu conteúdo foi examinado para retirada de partículas de solo e posteriormente sua massa foi pesada em balança analítica de precisão. A taxa de decomposição da serapilheira foi quantificada mediante avaliações de medidas da perda de massa, com a seguinte fórmula: Massa remanescente (%) = (massa final/massa inicial) x 100 Após o cálculo da massa remanescente ao longo do período, a constante de decomposição k foi calculada, segundo Thomas e Asakawa (1993), com o modelo exponencial: Xt = Xo . e-kt, Em que: Xt = peso do material seco remanescente após t dias; Xo = peso do material seco colocado nos sacos no tempo zero (t = 0). Já o tempo de meia vida (t1/2) foi calculado segundo Rezende et al. (1997) pela equação: t1/2 =ln(2)/K Em que: K = constante de decomposição calculada pela fórmula anterior (Xt = Xo . e-kt). RESULTADOS E DISCUSSÃO Produção de serapilheira Foi verificada diferença estatística para a deposição de serapilheira entre as três áreas estudadas. O maior valor de serapilheira adicionado foi observado na área de plantio de Carapa guianensis (9,20 Mg ha-1 ano-1), seguida pela área de plantio de Mimosa caesalpiniaefolia (9,06 t ha-1 ano-1) e pela floresta secundária com 7,63. Mg ha-1 ano-1. Balieiro et al. (2004a) encontraram aporte anual de serapilheira para plantios puros de guachapele (Pseudosamanea guachapele (Kunth) Harms, de eucalipto e para o consórcio das duas, respectivamente, 12,75; 11,84 e 12,44 Mg ha-1 num Planossolo, no município de Seropédica-RJ. Froufe (1999) na mesma localidade e tipo de solo, encontraram valores de 11,2 Mg ha-1 para um plantio de sabiá (Mimosa caesalpiniaefolia) seguido de regeneração natural. As áreas de plantio de Mimosa caesalpinaiefolia e plantio de Carapa guianensis apresentam maior aporte anual de material decíduo em relação à floresta de sucessão secundária espontânea, provavelmente pela presença de uma maior quantidade de espécies pioneiras. Já para a floresta de sucessão secundária espontânea, a maior parte da biomassa vegetal está retida na própria biomassa das espécies florestais, garantindo uma menor produção de material decíduo. De acordo com Meguro et al. (1979), o estádio de maturação do ecossistema é um dos fatores determinantes nas taxas de deposição de serapilheira e um ecossistema jovem deposita mais matéria orgânica que um estabilizado. As áreas de floresta de sucessão secundária espontânea, plantio de Mimosa caesalpiniaefolia e Carapa guianensis apresentaram diferença estatísticas em todas as estações avaliadas quanto ao aporte de material decíduo (Figura 1). De forma geral, a floresta de sucessão secundária espontânea teve a menor produção de serapilheira, em contraste com plantio de Mimosa caesalpiniaefolia que apresentou a maior produção de serapilheira na primavera inverno e o de Carapa guianensis com a maior no verão e outono (Figura 1). Corrêa Neto et al. (2001), estudando o aporte de serapilheira numa floresta de sucessão secundária espontânea e em um plantio de eucalipto na Floresta Nacional Mário Xavier em Seropédica-RJ, constataram maior deposição de serapilheira na estação inverno com valores de 2,39 Mg ha-1 e 2,01 Mg ha-1 para as respectiva áreas. Toledo et al. (2002), estudando o aporte de serapilheira em florestas de sucessão secundária espontânea tardia e inicial, no município de Pinheiral-RJ, observaram maior produção de serapilheira no inverno, sendo os maiores valores (4,03 Mg ha-1) verificados na área de floresta tardia e os menores (2,91 Mg ha-1) na área de floresta inicial. Moreira e Silva (2004) explicam que o fato da maior produção de serapilheira dar-se na estação seca é resultante da redução da precipitação acarretando ao ecossistema florestal um estresse hídrico que utiliza a queda de folhas para diminuir seu consumo de água. A floresta de sucessão secundária espontânea apresenta sua maior deposição de serapilheira no mês de novembro e a menor no mês de março. O plantio de Mimosa caesalpiniaefolia apresenta a maior deposição de serapilheira no mês de setembro e a menor no mês de abril. O plantio de Carapa guianensis tem sua maior deposição no mês de junho e, em maio, a menor deposição. As três áreas estudadas parecem não ter um padrão de deposição de serapilheira similar, pois pode observar-se que o período de maiores e menores deposições foi diferenciado. Para a área de floresta secundária espontânea, não foi verificada correlação (r2 = 0,11ns) entre a deposição de serapilheira e a precipitação pluviométrica ao longo do período estudado, demonstrando que talvez outras variáveis estejam atuando no padrão de deposição de serapilheira. Para a área de plantio de Mimosa caesalpiniaefolia, também não foi constada correlação entre a taxa de precipitação e o aporte de serapilheira (r2 = 0,68 ns) sendo o valor de correlação não-significativo. A área de plantio de Mimosa caesalpiniaefolia apresentou dois picos de deposição de serapilheira, nos meses entre dezembro de 2003 a fevereiro de 2004 e no período de julho a setembro de 2004. Tal fato pode ser explicado pela presença da Mimosa caesalpiniaefolia nesda área, espécie decídua que perde material foliar durante prolongados períodos de baixa e alta precipitação. Diferindo das demais, a área de plantio de Carapa guianensis, com posterior regeneração natural, apresentou correlação significativa (r2 = 0,44*), demonstrando que a precipitação influência de forma significativa na deposição do material decíduo nessa área. A área de plantio de Carapa guianensis com posterior regeneração natural apresentou maior deposição mensal nos meses de janeiro e junho. O padrão de deposição de material decíduo dessa área é similar ao do plantio de Mimosa caesalpiniaefolia com deposição da serapilheira pela influência da maior precipitação em janeiro, e pela menor precipitação em junho e, conseqüentemente, maior aporte de serapilheira em razão do estresse hídrico que provoca a senescência e queda sobretudo das folhas. A fração folhas foi a predominante nas três áreas avaliadas com 62, 69 e 75% para as áreas de floresta de sucessão secundária espontânea, plantio de Mimosa caesalpiniaefolia e plantio de Carapa guianensis respectivamente (Figura 2). Balieiro et al. (2004a) encontraram os seguintes percentuais da fração folhas 85,1; 57,7; e 56,4 % respectivamente para plantios puros de guachapele (Pseudosamanea guachapele (Kunth) Harms, eucalipto e consórcio das duas espécies. Araújo (2002), estudando a deposição da serapilheira em diferentes modelos de revegetação na reserva biológica de Poço das Antas em Silva Jardim, encontrou entre 69,1 a 76,2% de fração foliar. Observa-se que as áreas de plantio de Carapa guianensis e Mimosa caesalpiniaefolia apresentam percentuais da fração folhas, superiores ao da floresta de sucessão secundária espontânea, o que pode ser pela maior presença de espécies pioneiras que investem mais na produção de biomassa via folhas (Figura 2). Segundo Martins e Rodrigues (1999), a elevada deposição de folhas em florestas que apresentam distúrbios se deve ao rápido crescimento e renovação foliares das espécies pioneiras, canalizando os fotoassimilados em um primeiro momento para produção vegetativa. As áreas de plantio de Mimosa caesalpiniaefolia e Carapa guianensis apresentaram um percentual da fração galhos inferiores à floresta de sucessão secundária espontânea (Figura 2). A área de floresta de sucessão secundária espontânea depositou anualmente 149,81 kg ha-1 ano de nitrogênio, 3,00 kg ha-1 ano de fósforo e 16,30 kg ha-1 ano de potássio. O plantio de Mimosa caesalpiniaefolia 176,86 kg ha-1 ano de nitrogênio, 3,90 kg ha-1 ano de fósforo e 15,20 kg ha-1 ano de potássio. Já a área de regeneração contribuiu com 216,17 kg ha-1 ano de nitrogênio, 3,60 kg ha-1 ano de fósforo e 23,40 kg ha-1 ano de potássio. O aporte anual de nitrogênio pela serapilheira das áreas de plantio de Mimosa caesalpiniaefolia e plantio de Carapa guianensis foi superior à floresta de sucessão secundária espontânea, o que se pode dever, em parte, à capacidade das plantas leguminosas, tais como o pau-jacaré (Piptadenia gonoacantha (Mart.) Macbr.), encontradas na área de plantio de Mimosa caesalpiniaefolia e plantio de Carapa guianensis de formarem associações mutualísticas com bactérias fixadoras de N2. Essas espécies fixadoras de nitrogênio atmosférico também podem ter suas raízes colonizadas por fungos do solo, formando associações micorrízicas que contribuem para melhorar o aproveitamento de fósforo (Siqueira e Franco, 1988), o que explica o maior aporte anual de fósforo nas áreas de plantio de Carapa guianensis e Mimosa caesalpiniaefolia. Balieiro et al. (2004b) observaram maior aporte de N em área de plantio da leguminosa arbórea guachapele em relação ao plantio de eucalipto, evidenciando o benefício dessa no sistema. Quanto aos teores de nutrientes sazonais analisados na serapilheira, verifica-se que as áreas de plantio de sabiá e de Carapa guianensis apresentaram uma melhor qualidade nutricional se comparados à floresta de sucessão secundária espontânea (Tabela 1). De forma geral, as áreas de plantio de Mimosa caesalpiniaefolia e plantio de Carapa guianensis apresentaram teores de nitrogênio e fósforo superiores à floresta de sucessão secundária espontânea em praticamente todas as estações do ano (Tabela 1). O maior teor de fósforo foi encontrado no plantio de sabiá (Mimosa caesalpiniaefolia) no inverno e o menor na floresta sucessão secundária espontânea no verão e outono (Tabela 1). A área de plantio de andiroba (Carapa guianensis) na primavera foi a área que apresentou o maior teor de potássio e a floresta sucessão secundária espontânea e o plantio de sabiá no outono apresentaram o menor teor de potássio (Tabela 1). Tabela 1: Conteúdo sazonal de nitrogênio (N), fósforo (P) e potássio (K) aportados pela serapilheira das áreas floresta secundária espontânea (FS), plantio de sabiá (Mimosa caesalpiniaefolia) e plantio de andiroba(Carapa guianensis) na primavera (Pri), Verão (Ver), Outono (Out) e Inverno (Inv) em quilograma por hectare (kg ha-1). Table 1: Litter seasonal content of nitrogen (N), phosphorus (P) and potassium (K) of the areas spontaneous secondary forest (FS), sabiá (Mimosa caesalpiniaefolia) plantation and andiroba (Carapa guianensis) plantation in spring (Pri), summer (Ver), autumn (Out) and winter (Inv) in kilogram per hectare (kg ha-1).
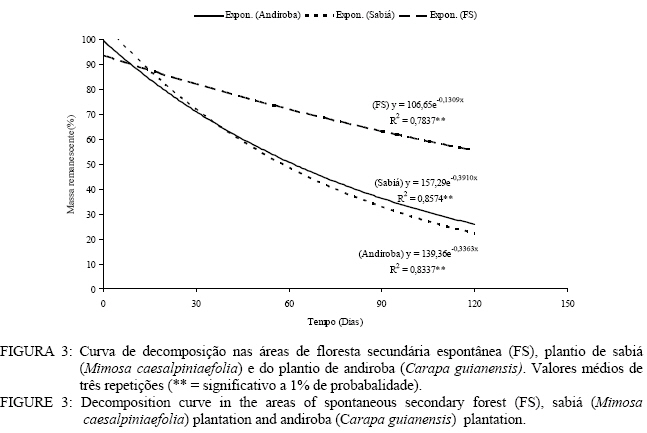
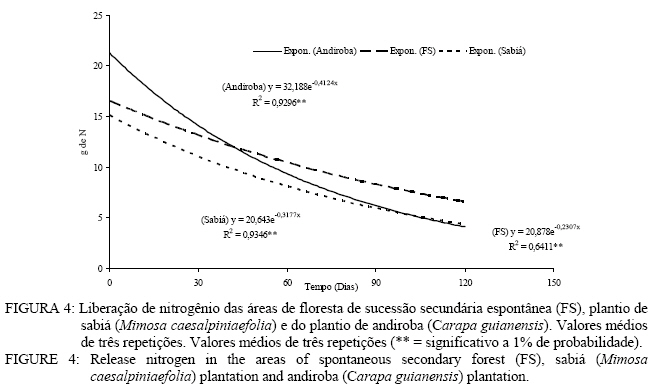
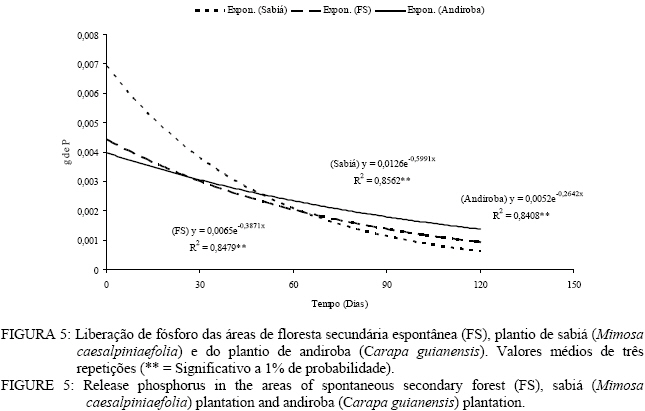
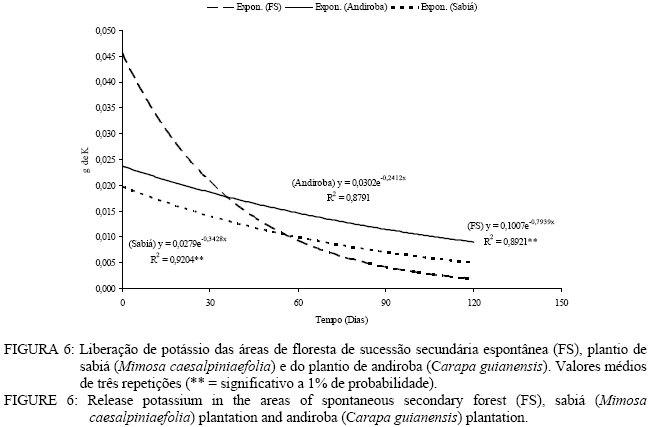
Em que: Valores seguidos da mesma letra na mesma coluna não diferem estatisticamente a 5% de significância pelo teste Tukey. Decomposição da serapilheira Observa-se que decorridos 30 dias após a instalação dos litter bags, aqueles instalados nas áreas de plantio de sabiá (Mimosa caesalpiniaefolia) e do plantio de andiroba (Carapa guianensis) foram os que apresentaram uma menor perda de massa (Figura 3). Entretanto, a partir de 30 dias, esse comportamento se inverte e as menores perdas de massa passam a ser verificadas na área de floresta de sucessão secundária espontânea (Figura 3). As áreas de plantio de Mimosa caesalpiniaefolia e Carapa guianensis tiveram maior perda de massa que a floresta de sucessão secundária espontânea o que se deve sobretudo a melhor qualidade do material aportado das leguminosas presentes, sendo a de principal ocorrência o pau-jacaré (Piptadenia gonoacantha (Mart.) Macbr.) nas áreas de plantio de Mimosa caesalpiniaefolia e de Carapa guianensis. Oliveira et al. (2003) verificaram maior taxa de decomposição por litter bags em consórcio de gramíneas com uma leguminosa herbácea quando comparado à gramínea pura. A introdução da leguminosa herbácea no consórcio com a gramínea acelerou a taxa de decomposição. A equação que descreve a velocidade de decomposição da floresta de sucessão secundária espontânea é o modelo matemático y = 106,65 e-0,1309x com índice de regressão de 0,78 apresentando significância a 1%. A constante de decomposição (K) e o tempo de meia vida (t1/2) foram, respectivamente, de 0,00194 g g-1 dia e 357 dias. Kolm e Poggiani (2003) estudando um plantio de Eucalyptus grandis W. Hill. ex Maiden com 0, 67 e 83% de desbastes, encontraram tempo de meia vida, respectivamente, de 452; 313 e 361 dias e constante de decomposição (K) de 0,56; 0,81 e 0,70 g g-1 dia. Na Figura 3, pode-se observar a equação que representa o modelo matemático e o índice de regressão significativo a 1% da velocidade de decomposição da área do plantio de sabiá. O valor da constante de decomposição (K) foi de 0,004974 g g-1 dia e o tempo de meia vidade 139 dias. Arato et al. (2003) encontraram tempo de meia-vida de 215 dias num sistema agroflorestal no município de Viçosa-MG. Balieiro et al. (2004b) obtiveram constante decomposição (K) de 0,00165 g g-1 dia e tempo de meia-vida de 421 dias para filódios de Acacia mangium Willd.em um plantio de sete anos com a espécie no município de Seropédica, valor superior ao encontrado neste estudo. O modelo exponencial que representa a velocidade de decomposição da serapilheira do plantio de andiroba pelo método dos litter bags encontra-se na figura 3 com coeficiente de correlação de 0,83, significativa a 1%. O tempo de meia-vida calculado foi de 179 dias, e a constante de decomposição (K) obtida foi de 0,003852 g g-1 dia. Balieiro et al. (2003) encontraram uma constante de decomposição (K) de 0,0047 g g-1 dia e o tempo de meia-vida (t1/2 ) 147 dias dos folíolos de Pseudosamanea guachapele Harms – Leguminosae. A floresta secundária espontânea apresentou maior tempo de meia-vida que as áreas de plantio de Mimosa caesalpiniaefolia e Carapa guianensis. Verifica-se que as áreas reflorestadas ou com regeneração com leguminosas arbóreas apresentam maior velocidade de decomposição com maiores constantes de velocidade de decomposição (K) e menores tempo de meia-vida, o que pode ser benéfico para no estabelecimento de programas de recuperação de áreas degradadas Aidar e Joly (2003) encontraram tempo de meia-vida de 469 dias (15,6 meses) e desaparecimento total da serapilheira em 584 dias (19,5 meses) de araribá (Centrolobium tomentosum Guill. ex Benth. – Fabaceae) em um fragmento de mata ciliar do Rio Jacaré-Pepira, São Paulo, SP. Balieiro et al. (2004a) encontraram menor tempo médio de residência (TMR) da serapilheira no solo num plantio consorciado de eucalipto com Pseudosamanea gachapele que num plantio puro de Eucalyptus grandis onde a introdução de material rico em nitrogênio pela leguminosa via deposição da serapilheira no plantio consorciado foi determinante na redução do TMR. Verificam-se, na Figura 4, as equações exponenciais e o valor da correlação entre a liberação de nitrogênio da serapilheira e o tempo em dias. As áreas em estudo demonstraram correlação significativa a 1% com valor r2 = 0,60. Quanto ao tempo de meia-vida (t1/2) e valor da constante de decomposição (K), foi observado os seguintes valores nas áreas de floresta de sucessão secundária espontânea (t1/2 = 10 dias e K = 0,067115 g.g-1. dia), plantio de Mimosa caesalpiniaefolia (t1/2 = 15 dias e K = 0,045923 g g-1 dia) e plantio de Carapa guianensis (t1/2 = 22 dias e K = 0,030299 g g-1 dia) respectivamente. O maior valor de tempo de meia vida e menor valor da constante de decomposição foi verificado para a área de plantio de Carapa guianensis e o menor tempo de meia-vida e maior valor da constante de decomposição na floresta de sucessão secundária espontânea. Após 30 dias de instalação dos litter bags, verifica-se que as três áreas apresentaram uma liberação de nitrogênio similar (Figura 4). Dos 30 aos 90 dias na área de plantio de Mimosa caesalpiniaefolia observa-se um aumento da liberação de nitrogênio em relação às demais áreas que se igualam nesse mesmo período quanto à liberação de nitrogênio. Nos últimos 30 dias, as áreas de plantio de Mimosa caesalpiniaefolia e de Carapa guianensis igualam-se na liberação de nitrogênio, entretanto a área de floresta de sucessão secundária espontânea apresentou um aumento no conteúdo de nitrogênio (Figura 4). Andrade (1997) registrou um pequeno aumento no teor de nitrogênio, em torno de 11% aos 160 dias após a instalação dos litter bags no campo, avaliando a decomposição de folhedo de Mimosa caesalpiniaefolia. Para o autor, esde aumento pode ser atribuído a vários fatores, como a grande deposição de N pelo litter fall, fixação biológica de nitrogênio, contaminações por solo, fezes e restos animais que podem ter enriquecido o folhedo em nitrogênio (Figura 4). As equações exponenciais que descrevem a liberação do fósforo assim como o valor de correlação são apresentados na Figura 5. A área de floresta de sucessão secundária espontânea apresentou tempo de meia-vida (t1/2 = 19 dias) valor de correlação (r = 0,85) com significância ao nível de 1% e valor da constante de decomposição de 0,036196. Os valores de tempo de meia-vida e da constante de decomposição (K) das áreas de plantio de Mimosa caesalpiniaefolia e plantio de Carapa guianensis foram de 48 e 16 dias e de 0,014478 e 0,043435 g g-1 dia. A área de plantio de Mimosa caesalpiniaefolia apresentou maior liberação de fósforo decorrido 120 dias em relação às demais áreas (Figura 5). As áreas de floresta secundária e plantio de Mimosa caesalpiniaefolia praticamente não apresentaram liberação de fósforo entre os 30 a 60 dias, mas dos 60dias ate o 120 dias a velocidade de liberação de nutrientes aumenta acentuadamente (Figura 5).Gama-Rodrigues et al. (2003) observaram menor liberação de fósforo em uma capoeira no sudeste da Bahia do que na floresta natural onde não apenas a qualidade do substrato regula o processo de liberação e decomposição dos nutrientes da serapilheira, mas também a qualidade do microambiente. Foi observada correlação significativa entre o tempo de exposição do material e o teor de potássio nas áreas de floresta de sucessão secundária espontânea, plantio de sabiá (Mimosa caesalpiniaefolia) e plantio de andiroba (Carapa guianensis) com valor de correlação (r2) superiores a 0,80 sendo todossignificativos ao nível de 1% de significância (Figura 6). Obtiveram-se os seguintes valores para tempo de meia-vida e constante de decomposição (K) nas áreas de floresta sucessão secundária espontânea (t1/2 = 17 dias e K = 0,040631 g.g-1. dia), plantio de Mimosa caesalpiniaefolia (t1/2 = 14 dias e K = 0,049792 g.g-1. dia) e plantio de Carapa guianensis (t1/2 = 14 dias e K = 0,049293 g.g-1. dia). Quanto ao tempo de meia-vida e a constante de decomposição (K), verificou-se que as áreas de plantio de sabiá e Carapa guianensis apresentaram valores similares. A área de floresta de sucessão secundária espontânea apresentou tempo de meia vida superior ao das áreas de plantio de Mimosa caesalpiniaefolia e de Carapa guianensis. A área da floresta secundária apresentou uma liberação de quase todo potássio aos 120 dias. Já as áreas de plantio de Mimosa caesalpiniaefolia e Carapa guianensis apresentaram uma velocidade de liberação de potássio mais lenta em relação à floresta secundária (Figura 6). Conclusões A floresta secundária espontânea e o plantio de sabiá (Mimosa caesalpiniaefolia) apresentaram a maior produção de serapilheira no inverno, porém o plantio de andiroba (Carapa guianensis) teve a maior produção de serapilheira no verão. A fração predominante no material formador da serapilheira foi as folhas com 60 % do total. As áreas de plantio de sabiá (Mimosa caesalpiniaefolia) e andiroba (Carapa guianensis) aportaram maior quantidade de nitrogênio e fósforo ao solo via serapilheira em comparação com a floresta secundária. As áreas de plantio de sabiá (Mimosa caesalpiniaefolia) e andiroba (Carapa guianensis) apresentaram maior perda de massa que a floresta de sucessão secundária espontânea. Referências Bibliográficas
Copyright 2006 - Ciência Florestal The following images related to this document are available:Photo images[cf06016t1.jpg] [cf06016f4.jpg] [cf06016f2.jpg] [cf06016f5.jpg] [cf06016f6.jpg] [cf06016f3.jpg] [cf06016f1.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}