 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ciência Florestal, Vol. 16, No. 4, 2006, pp. 399-405 VARIABILIDAD GENÉTICA DEL CRECIMIENTO EN PROGENIES SELECTAS DE Pinus radiata GENETIC VARIABILITY IN GROWTH OF SELECTED PROGENIES OF Pinus radiata Freddy Mora1 Francisco Zamudio2 1. Engenheiro Florestal, MSc., Doutorando em Genética e Melhoramento pela Universidade Estadual de Maringá, Av.
Colombo, 5790, Bloco 05, Sala 01, CEP 87020-900, Maringá (PR). fmora@universiabrasil.net Recebido para publicação em 22/06/2005 e aceito em 8/09/2006. Code Number: cf06036 RESUMEN Heredabilidad en sentido estricto (h2), correlación genética intra-clase (Γ2IC) y componentes de varianza para la altura, diámetro a la altura del pecho (DAP) y volumen del fuste, a los siete años de edad después de la plantación, fueron analizados en dos ensayos de progenie de Pinus radiata establecidos en el sur de Chile, en orden a examinar la variabilidad genética del crecimiento en una generación avanzada de la especie. Estos ensayos incluyeron árboles provenientes de propagación vegetativa y generativa. Las heredabilidades fueron estimadas para los árboles originados de semillas, y mostraron los siguientes valores: altura, 0.08-0.13; diámetro a la altura del pecho, 0-0.13; volumen del fuste, 0-0.09. Los valores de la correlación genética intra-clase, estimados para el grupo de árboles originados de propagación vegetativa, fueron moderados a bajos (Γ2IC: altura, 0.1-0.11; diámetro a la altura del pecho, 0.01-0.05; volumen del fuste, 0.03-0.06). No obstante la presencia de heredabilidades y correlaciones moderadas para algunas características y, la variación relativamente limitada entre grupos de árboles reflejan la fuerte selección realizada a las progenies. En general, los árboles provenientes de estacas presentaron una tendencia a la menor variación intra-genotípica, y evidenciaron magnitudes relativas de las varianzas debida al grupo (σ2G) mayores a las varianzas familiares (σ2F), en cuatro de seis caracteres evaluados, indicando una tendencia a la mayor homogeneidad en el crecimiento, característica común de la propagación vegetativa. Palabras-clave: Pinus radiata; correlación genética intra-clase; heredabilidad; estacas. ABSTRACT Narrow sense heritability (h2), intra-class genetic correlations (Γ2IC) and variance components for height, diameter at breast height and stem volume of seven-year-old trees were investigated. Two progeny trials of Pinus radiata were planted in southern Chile and growth data obtained to estimate genetic variability. These trials included trees from vegetative and generative propagation. Generative propagated trees were used to estimate the heritability, which indicated the following values: height, 0.08-0.13; stem diameter, 0-0.13; stem volume, 0-0.09. Estimates of intra-class genetic correlations among groups of trees from vegetative propagation were moderate to low (Γ2IC at 7 years of age: height, 0.1-0.11; stem diameter, 0.01-0.05; stem volume, 0.03-0.06). The relatively limited variation between trees groups was expected, reflecting the strong selection applied. In general, trees from cuttings displayed a tendency to have a smaller intra-genotypic variation, and a greater among groups variances (σ2G) than family variances (σ2F), in four out of six traits evaluated. This indicated a tendency for a greater homogeneity in growth, as is common in vegetatively propagated plants. Keywords:Pinus radiata;intra-class genetic correlation; heritability; cuttings. INTRODUCCION En los últimos años las exportaciones forestales chilenas han tenido un crecimiento contínuo, llegando a superar la barrera de los FOB US$ 2 mil millones. La principal especie productiva corresponde a pino radiata (Pinus radiata D. Don), seguida por diversas especies del género Eucalyptus (Barrios, 2004). La especie Pinus radiata es una de las protagonistas del desarrollo del sector forestal chileno, pasando de un consumo industrial de 3,7 millones de metros cúbicos en 1970, a 22,1 millones de metros cúbicos en el año 2003. El crecimiento observado se debe principalmente al esfuerzo nacional de forestación realizada hasta la década de los noventa, con una participación del pino radiata sobre el 80% de la nueva superficie anual de plantaciones (Pérez y Paredes, 2005). El grado de variabilidad genética que tiene Pinus radiata en el país le ha permitido ser objeto de selección y mejoramiento genético sostenido, para diversos propósitos comerciales. La variación genética es compleja, pero si se conocen su magnitud y tipo, ésta puede ser usada para obtener importantes ganancias en base a la selección de individuos genéticamente superiores (Zobel y Talbert, 1984). Existen diversos parámetros que permiten determinar el control genético, y por lo tanto la variabilidad genética asociada a alguna característica de interés. Entre las estimaciones más relevantes se encuentra la heredabilidad. El concepto de heredabilidad es uno de los más importantes y más usado en genética cuantitativa. El valor de la heredabilidad expresa la proporción de la variación en la población que es atribuible a diferencias genéticas entre individuos. La heredabilidad depende del coeficiente de parentesco y de la correlación intra-clase. Así, el cálculo de la correlación intra-clase permite observar la variabilidad genética entre diferentes grupos de árboles, vegetativos o generativos, cuando se desconoce el parentesco entre individuos (Falconer, 1976). Tales parámetros están en función de la estimación de los componentes de varianza, y una manera clásica de estimarlas es a través de los cuadrados medios (Giannini y Raddi, 1992; Montgomery, 1992; Vencovsky y Barriga, 1992). Rodríguez y López (2002), estimaron componentes de varianzas genéticas y parámetros afines en progenies de polinización abierta de Pinus sp., a través de las esperanzas de los cuadrados medios generadas por los análisis de varianza y covarianza. Para el caso de datos balanceados, la estimación de componentes de varianza por el método de mínimos cuadrados (análisis de varianza) es equivalente, i.e. conducen a resultados idénticos, a la estimación por máxima verosimilitud restringida - REML (Farias-Neto y Resende, 2001). El objetivo de este estudio fue analizar la variabilidad genética del crecimiento en familias selectas de Pinus radiata de siete años de edad, originadas de un huerto semillero de polinización controlada, a partir de dos ensayos de progenie ubicados en el sur de Chile, a través de las estimaciones de heredabilidad, correlación intra-clase y sus respectivos componentes de varianza. MATERIALES Y METODOS Dos ensayos de progenie de Pinus radiata fueron establecidos en el año 1990, en un diseño de bloques completos al azar, con cinco bloques, cinco progenies originadas de polinización controlada y cinco grupos de árboles provenientes de estacas de las mismas familias selectas evaluadas. Estas estacas fueron tomadas a partir de setos de un año de edad, los cuales provenían de un huerto semillero de polinización controlada. Los ensayos fueron instalados en las regiones del Bío-Bío y Araucanía, VIII y IX Regiones administrativas de Chile, respectivamente. El ensayo 1 (VIII Región) se localiza en el predio El Labrador, área de la ciudad de Angol, y el ensayo 2 (IX Región) en el predio San Antonio, área de Nacimiento. Los caracteres de crecimiento analizados fueron altura, diámetro a la altura del pecho (1,3 m) y volumen medidos a los 7 años de edad en cada ensayo. Para el cálculo del volumen se utilizó la siguiente fórmula: Volumen = (π / 4 * DAP2 *1,3) + (π /12 * DAP2 (H −1,3)) /10.000 (1) Donde DAP y H representan el diámetro a la altura del pecho (en centímetros) y la altura total del árbol (en metros), respectivamente. Heredabilidad, en sentido estricto, y correlación genética intra-clase fueron los parámetros genéticos estimados en los árboles originados de estacas y semillas, respectivamente, para cada característica de crecimiento y ensayo genético. Para los árboles originados de semillas, se determinaron los componentes de variación asociados a cada efecto en el siguiente modelo lineal: Yijk = μ + Ri + Fj + (RF)ij +εijk (2) Donde Yijk representa el valor observado del k-ésimo árbol de la j-ésima progenie en el i-ésimo bloque o replica, µ es el promedio total, Ri representa el efecto de la i-ésima replica, Fj representa el efecto aleatorio de la j-ésima progenie [NID (0,σ2 f )], (RF)ij representa el efecto aleatorio entre parcelas [NID (0,σ2 rf)], εijk representa el efecto residual aleatorio [NID (0,σ2 ε)]. La heredabilidad fue estimada en el grupo de árboles provenientes de propagación generativa, utilizándose la expresión:
Donde: h2 representa el valor de la heredabilidad estimada para progenies de hermanos completos, propio de un cruzamiento controlado (Falconer, 1976); σ2 f representa la varianza familiar;σ2 rf representa la varianza entre parcelas;σ2 ε) representa la varianza residual. La fracción del numerador corresponde a la varianza aditiva. La sumatoria del denominador corresponde a la varianza fenotípica, y k, es el coeficiente de relación familiar definida por Burdon et al. (1992a), con valor de ½ para hermanos completos. Para los árboles provenientes de estacas, el modelo lineal usado fue: Yijk = μ + Ri + Gj + (RG)ij +εijk (4) Donde Yijk representa el valor observado del k-ésimo árbol originado de estaca en el j-ésimo grupo dentro del i-ésimo bloque o replica, µ es el promedio total, Ri representa el efecto de la i-ésima replica, Gj representa el efecto aleatorio del j-ésimo grupo (5 grupos) [NID (0,σ2 g)], (RG)ij representa el efecto aleatorio entre parcelas [NID (0,σ2 rg)], εijk representa el efecto residual aleatorio [NID (0,σ2 ε)]. Para el grupo de árboles provenientes de propagación vegetativa, la heredabilidad no fue estimada debido a que se desconocía el origen exacto de las estacas dentro de cada progenie (parentesco desconocido). Sin embargo, el cálculo de la correlación intra-clase permite observar el grado de variabilidad entre los diferentes grupos de estacas, independiente del parentesco (Falconer, 1976). La correlación intra-clase fue estimada para los caracteres métricos altura, diámetro y volumen en cada ensayo (a la edad de 7 años). La estimación de esta correlación fue realizada de acuerdo al procedimiento usado por Falconer (1976), que define tal correlación a partir de la expresión general: 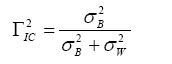 (5)
(5)
Donde σ2 B es la componente entre grupos genéticos y σ2 Wes la componente dentro de los grupos. Γ2 IC representa la correlación intra-clase. Falconer (1976) sostiene que mientras mayor sea la similitud dentro de los grupos, mayor será la proporción de diferencia que exista entre los grupos. El grado de parecido entre individuos puede, por lo tanto, expresarse como la componente entre grupos en proporción a la varianza total. A partir del modelo en (4), el componente entre grupos, para el presente estudio, se deriva a partir de la expresión de covarianza: COV(Yijk;Yijk´) = σ2 G ; Donde σ2 G representa la varianza debida al grupo. La varianza fenotípica o total (σ2 p) se deriva desde la expresión: VAR(Yijk) = σ2 p= E[(Yijk-E(Yijk)]2 = E[Yijk–(μ+Ri)]2 = σ2 g+σ2 rg+σ2 ε; aquí σ2 ε es la varianza residual y σ2 rg representa la varianza de la interacción grupo-replica. Luego, la correlación intra-clase de este estudio puede ser estimada para cada carácter métrico, en cada ensayo, a partir de la siguiente expresión (White y Hodge, 1989; Burdon y Shelbourne, 1974): 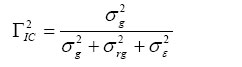 (6)
(6)
La relación entre la varianza genética y varianza fenotípica provee una estimación cuantitativa de la importancia relativa de los factores genéticos versus los ambientales (Hansche, 1983). Los componentes de varianzas, parámetros de heredabilidad y correlación intra-clase, fueron estimados a través del procedimiento de modelos lineares mixtos PROC MIXED de SAS® (SAS Institute, 1996). RESULTADOS Y DISCUSIÓN La varianza residual o varianza dentro de las familias (σ 2ε ) fue la mayor fuente de variación en ambos ensayos (Tabla 1). La magnitud relativa de esta fuente de variación (expresada en porcentaje de la variación total) fue de 93%, para el diámetro y volumen en ambos ensayos, y de 87 y 90 % para la altura en los ensayos de Angol y Nacimiento, respectivamente. Las heredabilidades de la altura y diámetro en el ensayo de Angol fueron moderadas (h2=0,13 en ambas características) (Tabla 1). Burdon et al. (1992a), estimó la heredabilidad en sentido restringido en un ensayo de Pino radiata de 8 años de edad, obteniendo valores moderados en diámetro y altura (h2 = 0,1 y entre 0,1-0,2, respectivamente). Shelbourne y Low, en Pederick (1990), reportaron valores moderados de la heredabilidad para el crecimiento en diámetro (h2=0,12), en un ensayo de progenie de pino radiata de 7 años de edad. TABLA 1: Componentes de varianza y heredabilidad (h2) para los caracteres de crecimiento de las familias de pino radiata originadas de semillas de polinización controlada, a los 7 años de edad, en las localidades de Angol y Nacimiento, sur de Chile. TABLE 1: Variance component and heritability (h2) per growth trait, of seven-year-old seedling among radiata pine families, in Angol and Nacimiento localities, southern Chile.
En que: σ2F = varianza familiar; σ2rf = varianza entre parcelas; σ2ε = varianza residual; σ2p = varianza fenotípica; σ2a = varianza aditiva. La heredabilidad del volumen en Angol fue relativamente inferior a de la altura y diámetro (h2=0,092), debido a que la varianza familiar tuvo menor incidencia en la variación total (alrededor del 5 %), arrojando una varianza genética aditiva menor. Burdon et al. (1992a), estimó un valor similar para la heredabilidad en volumen (h2=0,1), en progenies provenientes de polinización abierta de Pino radiata de 8 años de edad. Cotterill y Dean (en Borralho et al., 1992a), reportaron tendencias distintas en heredabilidad para el crecimiento en volumen de pino radiata durante todo el periodo de crecimiento. Los componentes de varianza familiar (σ2f) no fueron significativos para el diámetro y volumen en Nacimiento, estimando heredabilidades con valores cero (h2=0, Tabla 1). Algunos estudios han reportado un incremento en la varianza aditiva de la altura en Pinus spp., con una suave declinación a la edad de 7 años (Foster, 1986). Varianzas genéticas aditivas iguales a cero fueron obtenidas por Karlsson y Russell (1990), ya que los componentes de varianza familiar fueron negativos o despreciables en diversas características de crecimiento. La heredabilidad de la altura en Nacimiento fue baja (h2=0,076). Pederick (1990), estimó la heredabilidad en sentido restringido para pino radiata y obtuvo valores bajos en altura y diámetro (alrededor de 0,09), en un ensayo con 18 familias de 12 años de edad. Las estimaciones de los componentes de varianza y la correlación genética intra-clase de los árboles originados de estacas enraizadas (Tabla 2) para los caracteres altura, diámetro y volumen a los 7 años de edad, indicaron que la varianza dentro del grupo o varianza residual (σ2e) en el ensayo de Angol alcanzó magnitudes de hasta 96 y 99% de la variación total, en volumen y diámetro, respectivamente. En el ensayo de Nacimiento, estas magnitudes disminuyeron a 90% para volumen, y 86% para diámetro. En ambos ensayos, la varianza residual de la altura fue menor a las otras características de crecimiento (86 y 79%, para los ensayos de Angol y Nacimiento, respectivamente). En experimentos de propagación vegetativa, o comparación clonal, es posible reducir la variabilidad genética, lo que permitirá minimizar la variación residual (Zobel y Talbert, 1984). En el presente estudio, la variación dentro de los grupos de estacas se debe a efectos tanto genéticos como ambientales, ya que las estacas fueron tomadas de setos diferentes dentro de cada familia. En ambos ensayos todas las varianzas debidas al grupo fueron positivas (σ2g>0). Las magnitudes mayores fueron en altura, las que alcanzaron valores de hasta 10 y 11 % de la variación total, en Angol y Nacimiento, respectivamente. El cálculo de la heredabilidad para el crecimiento de los propágulos vegetativos no fue estimado, debido a la ausencia de información del origen genético de las estacas dentro de cada familia. Esta información es imprescindible para poder estimar el grado de parentesco de los propágulos (Falconer, 1976). No obstante lo anterior, la correlación genética intra-clase permitió estimar la calidad del control genético presente en los grupos de estacas. TABLA 2. Componentes de varianza y correlación genética intra-clase (Γ2IC) de los árboles originados de estacas de pino radiata, por característica de crecimiento, a los 7 años de edad. TABLE 2. Variance components and intraclass genetic correlation (Γ2IC) among radiata pine cuttings, per traits, after 7 years in the field.
En que: σ2G = varianza debida al grupo; σ2rg = varianza entre parcelas; σ2e = varianza residual; σ2TOTAL = varianza total. Los controles genéticos del diámetro y del volumen, en ambos ensayos, fueron bajos (Γ2IC=0,0132 – 0,054 y 0,0324 – 0,063 respectivamente). La correlación genética intra-clase fue superior a estos valores para el crecimiento en altura (Γ2IC =0,1001 y 0,1149, para Angol y Nacimiento, respectivamente), debido a que la varianza del grupo tuvo mayor incidencia en la variación total. Burdon et al. (1992b), trabajando con estacas de pino radiata, reportaron un alto control genético para altura a los 8 años de edad (H2 = 0,26-0,41). La varianza residual fue la mayor fuente de variación en ambos tipos de propágulos. Las magnitudes relativas más bajas fueron encontradas en el carácter altura, registrándose una menor variación en los árboles originados de estacas que crecieron en Nacimiento (79%). Con la utilización de estacas enraizadas en plantaciones forestales, se espera lograr una mayor uniformidad al eliminar la variación genética dentro de los clones (Roulund, 1974). Las magnitudes relativas de las varianzas debida al grupo (σ2G) fueron mayores a las varianzas familiares (σ2F) para altura en el ensayo de Angol, y altura, diámetro y volumen en el ensayo de Nacimiento. Un caso destacable fue la varianza debida al grupo en altura, que mostró magnitudes de 10 y 11% de la variación total (Angol y Nacimiento, respectivamente). En las plantas provenientes de semillas, estas magnitudes variaron apenas entre 7 y 4%. Los árboles originados de estacas mostraron una correlación intra-clase siempre mayor que cero, pero también con valores bajos (correlación del diámetro) a moderados (correlación del carácter altura, Tabla 2). Por otro lado, los árboles originados de semillas exhibieron heredabilidades con valor cero en Nacimiento (heredabilidad del diámetro y volumen), debido a la varianza familiar baja o nula, y valores moderados para altura y diámetro en Angol (h2=0,13). La variabilidad genética expresada a través de los parámetros estimados en los árboles originados de estacas y semillas de pino radiata, puede cambiar en las distintas etapas de su desarrollo, dependiendo del efecto de maduración de los propágulos y de la depresión endogámica producido por autocruzamiento, entre otros (Burdon et al. 1992a; Burdon et al. 1992b; Borralho et al. 1992b). CONCLUSIONES La variabilidad genética dependió de las variables de crecimiento y de las condiciones ambientales en el cual se desarrollaron las progenies. Heredabilidades de la altura con valor moderado en Angol, y correlaciones intra-clase moderadas en ambos ensayos indican que aún persiste la obtención de cierta variabilidad genética. Sin embargo, a pesar de la existencia de heredabilidades y correlaciones intra-clase moderadas para algunas características, la variación relativamente limitada entre grupos de árboles era esperada, debido al rigor de la selección aplicada a las progenies. AGRADECIMIENTOS A la empresa Forestal MININCO S.A., por el apoyo y material concedido para la realización de este trabajo. REFERENCIAS BIBLIOGRAFICAS
Copyright 2006 - Ciência Florestal The following images related to this document are available:Photo images[cf06036t2.jpg] [cf06036t1.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

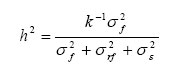 (3)
(3)