 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ciência Florestal, Vol. 17, No. 2, 2007, pp. 175-185 MODELOS DE CRESCIMENTO RESULTANTES DA COMBINAÇÃO E VARIAÇÕES DOS MODELOS DE CHAPMAN-RICHARDS E SILVA-BAILEY APLICADOS EM Leucaena leucocephala (Lam.) de Wit GROWTH RESULTING FROM THE VARIATION AND COMBINATION OF MODELS OF CHAPMAN-RICHARDS AND SILVA-BAILEY APPLIED TO Leucaena leucocephala (Lam.) of Wit Cícero Carlos Ramos de Brito1 José Antonio Aleixo da Silva2 Rinaldo Luiz Caraciolo Ferreira3 Eufrázio de Souza Santos4 Ivan Ferraz5 1. Licenciado em Matemática, MSc., Professor do Panorama
Colégio e Curso, Av. Professor Cláudio Selva, 178, Dois
Irmãos, CEP 52171-260, Recife (PE). cicerocarlosbrito@yahoo.com.br Recebido para publicação em 13/04/2006 e aceito em 4/04/2007. Code Number: cf07021 RESUMOO objetivo deste trabalho foi desenvolver novos modelos de crescimento para recursos florestais aplicados à leucena [Leucaena leucocephala (Lam.) de Wit], tendo como base as hipóteses biológicas propostas por Chapman-Richards e Silva-Bailey. O experimento de leucena foi conduzido na Estação Experimental da Empresa Pernambucana de Pesquisa Agropecuária – IPA, Caruaru, PE. Foram utilizadas 544 árvores de leucena de um experimento com vinte remedições realizadas ao longo de 12 anos. Compararam-se novos modelos de crescimento resultantes da combinação e variações dos modelos de Chapman-Richards e Silva-Bailey, bem como outros comumente usados em recursos florestais. Para a seleção das equações, utilizaram-se o Índice de Ajuste (IA), o erro-padrão da estimativa e a distribuição gráfica dos resíduos. Os resultados indicaram que todos os modelos testados se ajustaram de maneira satisfatória aos dados, podendo ser utilizados para se estimar o crescimento em altura da leucena. Palavras-chave: Recurso florestal; leucena; biometria. ABSTRACT The main objective of this work was to develop new growth models for forest resources, applied to (leucena) [Leucaena leucocephala (Lam.) of Wit], based on the biological hypotheses proposed by Chapman-Richards and Silva-Bailey. The experiment of the leucena was carried out in the Experimental Station of the Agricultural and Liverstock Research Company of Pernambuco - IPA, in the municipal district of Caruaru - PE. Five hundred and forty four trees of Leucaena leucocephala (Lam.) of Wit were used, with 20 measures along the time. It was compared new growth models resulting from the combination and variation of the Chapman-Richards and Silva-Bailey models, as well other models used in forest resources. For the selection of the final equations, the fit of index, standard error of estimate and the graphic distribution of the residues were used. Results show that new models were as good as Chapman-Richards and Silva-Bailey and other tested. Therefore, any one of them can be applied to estimate the growth of leucena. Keywords: Forest resource; leucena; biometric. INTRODUÇÃO No planejamento de manejo de recursos florestais, uma variável de extrema importância é o crescimento. Sua modelagem é fundamental na prognose da produtividade, qualidade do local e dinâmica de populações. Na Engenharia Florestal, os modelos são imprescindíveis na prognose dos futuros recursos florestais, na escolha de metodologias silviculturais e apoio à tomada de decisões no manejo e na política florestal (Spathelf e Nutto, 2000). Tais modelos são sínteses de observações e/ou fenômenos biológicos, aplicados nas condições sob as quais foram coletados os dados. Com o avanço dos procedimentos computacionais, técnicas matemáticas mais sofisticadas são utilizadas no desenvolvimento de novos modelos aplicados em estudos de crescimento, uma vez que, geralmente, as hipóteses básicas utilizadas geram modelos que alterando os valores de alguns coeficientes, de positivo para negativo, a(s) equação(ões) resultante(s) pode(m) ser empregada(s) para estudos de acréscimos ou decréscimos na produção, no crescimento e, ou sobrevivência/mortalidade. Segundo Scolforo (1994), o conhecimento do crescimento e da produção presente e futura de árvores em povoamentos florestais é elemento fundamental no manejo florestal sustentável, sendo necessário possuir informações sobre a existência de relações quantitativas e modelos matemáticos que sejam consistentes e, numericamente, compatíveis para a predição do desenvolvimento do povoamento em qualquer idade. Dada a dinâmica com que os modelos matemáticos vêm sendo aplicados nas pesquisas biológicas, novos estudos em ciências florestais são altamente justificáveis. O objetivo deste trabalho foi gerar novos modelos de crescimento pela combinação ou adaptações dos modelos de Chapman-Richards e Silva-Bailey, compará-los entre si e com os modelos comumente usados na pesquisa florestal. REVISÃO DE LITERATURA Um modelo é uma formulação que simula um fenômeno do mundo real, de forma a ser possível fazer previsões, e pode ser utilizado nos mais diversos campos. Em suma, não se pretende que um modelo, independentemente da área em estudo, seja uma cópia exata do mundo real, mas, sim, uma simplificação que revele os processos-chave do fenômeno em causa, de forma a ser possível perceber e prever novas situações dentro do universo em estudo. Sendo assim, um bom modelo é um espelho eficiente da realidade (Brito, 2005). Um modelo pode ser definido como uma formulação matemática baseada em hipóteses que tentam representar fenômenos físicos ou sistemas biológicos, com a finalidade de gerar uma equação que possa representar tal(is) fenômeno(s) a um determinado nível de confiabilidade α, por exemplo: Y=β0 +β1 X1i +...+βn Xni + εi A diferença principal entre um modelo de crescimento empírico e um biológico é que o primeiro recorre a uma expressão matemática de uma relação entre variáveis que podem ser esperadas logicamente, enquanto o segundo é expressão de uma hipótese lógica relativa à biologia do processo de crescimento (Pienaar, 1965). Um modelo matemático somente representa um conjunto de suposições físicas ou biológicas. Essas suposições são sempre simplificações da realidade (France e Thornley, 1984). Nas ciências, os modelos matemáticos são muito importantes, e usam uma ferramenta especial: as equações. Um modelo de crescimento ou mortalidade pode abranger um sistema de equações, sendo capaz de prognosticar a produção sob várias condições (Spathelf e Nutto, 2000). Segundo Clutter (1963), uma equação de crescimento, mesmo que apresente bons resultados em termos de ajuste a um determinado conjunto de dados, deve apresentar duas importantes características: convergência e invariância. Por “convergência” se entende que, quando o tempo final tf tender para o tempo inicial ti, ou vice-versa, o valor obtido para o crescimento final Wf deve ser igual ao obtido no tempo inicial Wi. Invariância por sua vez é a propriedade que uma equação possui de predizer um valor futuro no tempo tf sem depender de tempos intermediários, isto é, indiretamente de um ti para tf. Segundo Prodan (1968), nas ciências florestais, biológicas, sociológicas, econômicas, etc, um grande número de tentativas tem sido feito para exprimir o crescimento mediante fórmulas matemáticas. As tentativas em florestas estão, provavelmente, entre as primeiras no campo biológico, destacando-se os estudos de Spath em 1797, Hossfeld em 1822 e Smalian em 1857. O desenvolvimento de novos modelos de crescimento e sobrevivência/mortalidade tem tido avanços consideráveis nas ciências florestais, como o que ocorreu na generalização de Chapman-Richards (Turnbull, 1963) para o modelo de Bertalanffy (1957) e trabalhos de Prodan (1968) dando um tratamento compreensivo de várias funções de crescimento e leis de crescimento que já têm sido estudados com referência ao crescimento florestal. Entre os modelos clássicos utilizados no estudo de crescimento podem-se citar: Modelo de Mitscherlich Em 1930, Mitscherlich utilizou um princípio usado na economia (a produção se aproxima de um limite e o incremento na produção tende a zero), traduzindo-o em termos de crescimento biológico como “Lei dos efeitos dos fatores de crescimento” (Silva, 1986). Essa lei diz que a taxa de crescimento dW/dt é proporcional à diferença entre o tamanho máximo (último nível de crescimento ou termo assintótico) e o próprio parâmetro de crescimento.
Em que: W = parâmetro de crescimento; U = termo assintótico (significa o último nível de crescimento); c' = c1 = c2 = ... = cn; t= tempo. Modelo de Bertalanffy Bertalanffy (1934, 1957) derivou seu modelo de estudos das então chamadas relações alométricas dos organismos. A relação alométrica será compreendida por existir entre elementos dimensionais de um organismo, denominado P e Q, e foi expresso por:P = α · Qθ . Em que: P correspondia ao comprimento do fêmur; Q era a largura do crânio de certas espécies animais ou a área fotossintética e área do cambio do tronco de espécies vegetais; α e θ são parâmetros da relação. Isso sugere a suposição de que em indivíduos normais, o padrão de crescimento específico de P produzirá uma relação proporcional constante para o padrão de crescimento específico de Q, que é:
Estudos extensivos de muitos organismos aquáticos e terrestres levaram Bertalanffy à conclusão de que para uma classe principal de organismos a relação alométrica entre a área da superfície (também total, ou área absortiva total) e volume total pode ser adequadamente expresso como:
Em
que: S = área da superfície; V = volume do organismo; Após prosseguimento dos estudos, o autor chegou à conclusão de que o crescimento segue um processo de síntese (anabolismo) e degradação (catabolismo). A fórmula original foi expressa da seguinte forma:
A vantagem da equação está na sua fundamentação teórica. A razão anabólica é proporcional à área da superfície do organismo, enquanto que a razão catabólica é proporcional ao volume da biomassa (SILVA, 1986). Modelo de Chapman-Richards Richards (1959), estudando o crescimento de plantas e Chapman (1961), o crescimento de peixes, consideraram que a constante alométrica de 2/3 do modelo de Bertalanffy era muito restritiva, pois esse expoente pode assumir diferentes valores dependendo da natureza da população. Nas Ciências Florestais, Richards foi o primeiro a aplicar a equação de crescimento desenvolvida por Bertalanffy (France e Thornley, 1984). A generalização de Chapman-Richards do modelo de crescimento de Bertalanffy é uma expressão matemática de uma hipótese concernente a causas essenciais do fenômeno de crescimento, de tal modo que os parâmetros no modelo têm pelo menos uma total interpretação biológica ou fisiológica (Turnbull, 1963; Pienaar e Turnbull, 1973). Assim sendo, fazendo essa modificação obtém-se:
Em que: U = valor assintótico (tamanho teórico máximo que o organismo pode atingir). Esse modelo foi nomeado como modelo de crescimento de Chapman-Richards por Turnbull (1963) e tem sido usado freqüentemente em pesquisas de florestas (Pienaar, 1965; Machado, 1978; Campos, 1980; Clutter et al, 1983; Barros et al, 1984; Mctague, 1985; Silva, 1986; Scolforo, 1994; Fekeduleng et al, 1999; Santos Júnior, 2005). Dependendo do valor do parâmetro m, o modelo de Chapman-Richards representa várias leis de crescimento como, por exemplo: o modelo de Bertalanffy (m = 2/3), a função de crescimento Monomolecular (m = 0), a função de crescimento Gompertz (m = 1) e a função de crescimento Logístico (m = 2). Tais funções são casos especiais da função de crescimento de Chapman-Richards (Machado, 1978). Clutter e Jones (1980) desenvolveram um procedimento em que o valor assintótico U que é constante no modelo, pode ser eliminado e substituído por um valor que corresponde um determinado valor de W no tempo i. Para isto usaram o método das diferenças algébricas. Assim, tem-se: Em um tempo inicial i, o modelo é:
Em que um tempo final f, o modelo é:
Como U é constante nos tempos i e f, pode-se conhecer seu valor, por exemplo, no tempo i:
Substituindo (2) em (1), tem-se:
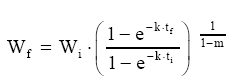
Dessa forma, o valor assintótico que, às vezes, por ser teórico, pode apresentar resultados fora da realidade, é eliminado do modelo para dar lugar a um valor real Wi. Modelo de Silva-Bailey Silva (1986), estudando o crescimento de Pinus elliottii Engelm var. elliottii em função de diferentes fontes de nutrientes, considerou a idéia de dinâmica de populações proposta por Leslie (1945) que diz que, para qualquer distribuição arbitrária no tempo i, a estrutura de uma população pode ser expressa na forma de n+1 equações lineares, em que de n para n+1 é o ultimo grupo de idade na tabela de distribuição etária. O modelo expressa o tamanho da população (Wi+1) no tempo i+1 como função do tamanho da população no tempo i(Wi), da taxa de sobrevivência (S), do número médio de recrutamentos (F) produzido na população no tempo i. Sua expressão é: Wi+1=Wi(SF)n. Em populações florestais, o problema de recrutamento (nascimento ou migração) no período de investigação é desprezível, bem como migração que são fenômenos comuns em populações animais (móveis). Ocorre mortalidade, mas esta pode ser modelada sem maiores problemas. Dessa forma, Silva (1986) considerou o plantio de Pinus como uma população fechada e o parâmetro F pode ser retirado do modelo. O parâmetro S é desconhecido e depende da taxa de mortalidade que pode ser regular ou irregular (Burkhart, 1974, citado por Silva, 1986). Mortalidade regular é resultante de fatores tais como densidade de plantio, qualidade do sítio, tratamentos silviculturais, etc, enquanto que mortalidade irregular é conseqüência de eventos catastróficos como fogo, doenças, pragas, ventos, etc. MATERIAL E MÉTODOS Os dados de leucena foram provenientes de um experimento implantado em 1989, localizado na Estação Experimental da Empresa Pernambucana de Pesquisa Agropecuária (IPA), no município de Caruaru, no estado de Pernambuco, que possui as seguintes coordenadas geográficas: Latitude 08°14’18’’S; Longitude 38°00’00’’ WGr. e altitude de 537 m. O clima, pela classificação climática de Thornthwaite, é do tipo Dd’a’ (semi-árido megatérmico). O solo caracterizado pela associação de Neossolo (EMBRAPA, 1999). Os tratamentos foram constituídos de duas estirpes de rizóbio, mais tratamentos comparativos nitrogenados em três níveis de adubação fosfatada e na ausência ou presença de composto de resíduo urbano, perfazendo inicialmente, um total de 24 tratamentos com 24 repetições. Para este estudo, baseando-se nas pesquisas de Meunier (1991), Souza (2003) e Santos Júnior (2005) em que se constatou que o efeito do composto orgânico influenciou o crescimento das mudas e das plantas até o quarto ano, decidiu-se pela formação de dois grupos: os que não possuem composto orgânico e os que possuem. Foram utilizadas para este trabalho 544 árvores sobreviventes das 576 plantadas no início do experimento. A variável altura total (h) foi medida em todos os indivíduos durante 12 anos, sendo tomadas 19 medidas. O tempo inicial considerado foi de 120 dias que correspondem à idade em que as mudas foram plantadas no campo. Modelos utilizados Neste trabalho, todos os modelos empregados foram derivados usando a metodologia das diferenças algébricas proposta por Clutter e Jones (1980), com base nos modelos:
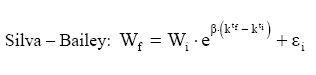
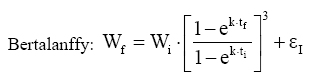
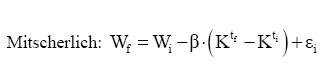
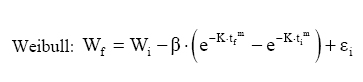
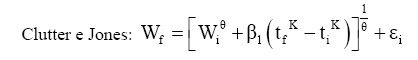
Em que: Wf = altura ou comprimento final; Wi = altura ou comprimento inicial; β, m, θ e K = parâmetros dos modelos; ti e tf = Tempo inicial e final das medições das alturas ou comprimento; εi= erro aleatório. Dedução dos modelos matemáticos propostos Modelo proposto 1 O primeiro modelo desenvolvido foi resultante da combinação (produto) dos modelos Chapman-Richards e Silva-Bailey, pelo fato de que são dois modelos que têm apresentado bons resultados nas estimativas de espécies florestais. Foram consideradas duas situações: a soma e o produto dos modelos. Como o proveniente do produto apresentou melhores resultados, este foi o considerado neste trabalho. Sejam Considere-se: W=(W'·W'') . Logo substituindo tem-se que: que após seu desenvolvimento resulta
em Modelo proposto 2 Considere-se que a variação do crescimento em função do tempo é proporcional a uma condição inicial, multiplicado pela diferença entre o anabolismo e o catabolismo do indivíduo ao longo do tempo.
Assim, Modelo proposto 3 Considerando-se que a variação do crescimento em função do tempo é proporcional à diferença entre o anabolismo e o catabolismo ao longo do tempo; tem-se:
Análise das equações de regressão Utilizou-se para os ajustes dos modelos o procedimento NONLINEAR do Programa Computacional SYSTAT 05 for Windows (Demo) sendo utilizado o método Simplex para calcular as estimativas dos parâmetros. Os critérios usados nas comparações das equações foram o Índice de Ajuste (I.A.) (Schalaegel, 1981), conforme segue:
Em
que: IA% =
Índice de Ajuste em percentagem; Yi = Valor observado (real); Em equações ajustadas pelo método dos mínimos quadrados, o índice de ajuste corresponde ao coeficiente de determinação R2. Para o cálculo da análise da distribuição gráfica residual, os resíduos foram obtidos pela expressão:
Em
que: Ei=
resíduo da i-ésima observação;Yi= altura dominante observada; O erro-padrão da estimativa percentual (Syx %) foi obtido pela fórmula (SCHNEIDER, 1998):
Em
que: Syx%=
erro-padrão da estimativa em percentagem; QMR = Quadrado Médio do Resíduo do
modelo ajustado; RESULTADOS E DISCUSSÃO Nas Tabelas 1 e 2, são apresentadas as estimativas dos parâmetros dos modelos testados e seus Índice de Ajustes. Observa-se que todos os modelos resultaram em equações com altos valores dos Índices de Ajuste, sendo que a equação de Bertalanffy apresentou o menor I.A., porque trabalha com valores do parâmetro m = 2/3, isto é, um valor fixo. TABELA 1: Estimativas dos parâmetros e índices de ajuste dos modelos testados para modelagem de crescimento de leucena, no tratamento 1, Estação Experimental da Empresa Pernambucana de Pesquisa Agropecuária, Caruaru, PE. TABLE 1: Estimates of the parameters and index of adjustments of the models tested for modelling of leucena growth, in treatment 1, Experimental Station of the Agricultural Research Company in Caruaru, PE.
Em que: IA = índice de ajuste (%). Os modelos que apresentaram os mais altos valores de Índices de Ajuste foram os de Weibull e Mitscherlich, com valores, respectivamente, de 99,184 e 99,178 para o tratamento 1. No tratamento 2, o modelo de Weibull apresenta um índice de 99,112, seguido do modelo proposto 2 com 99,108. melhor distribuição residual. Pelo fato do tratamento 2 levar em consideração o efeito do composto orgânico, ocorreu maior variabilidade, resultando em valores de Índices de Ajuste inferiores ao tratamento 1. TABELA 2: Estimativas dos parâmetros e índices de ajuste dos modelos testados para modelagem de crescimento de leucena, no tratamento 2, Estação Experimental da Empresa Pernambucana de Pesquisa Agropecuária, Caruaru, PE. TABLE 2: Estimates of the parameters and index of adjustments of the models tested for modelling of leucena growth, in treatment 2, Experimental Station of the Agricultural Research Company in Caruaru, PE.
Não foram obsevadas diferenças entre os modelos propostos e os utilizados freqüentemente na literatura florestal como critérios comparativos, o que comprova o bom ajuste dos modelos propostos. O intuito de desenvolver novos modelos, teve como princípio a aplicabilidade da modelagem matemática em áreas biológicas, uma vez que as diferentes situações encontradas podem gerar novas hipóteses biológicas e conseqüentemente novos modelos. E isto é um dos objetivos da biometria que, fundamentalmente, envolve uma interação entre as ciências matemática, estatística e a biológica. Análise da distribuição gráfica residual dos modelos testados A análise gráfica de distribuição dos resíduos indicou que os modelos apresentados se mostraram adequados para a estimação do crescimento em altura para a leucena, sendo perceptível o pior ajuste para o modelo de Bertalanffy que considera o parâmetro m fixo, igual a 2/3 (Figuras 1, e 2a, 2b ). CONCLUSÕES A análise do Índice de Ajuste e da distribuição residual indicou que os modelos propostos apresentaram ajustes semelhantes aos comumente utilizados em recursos florestais para as estimativas de crescimento da leucena. O ajuste dos modelos utilizados para leucena no segundo grupo das plantas que receberam o composto orgânico, com exceção do terceiro modelo proposto, foram inferiores ao grupo 1 das plantas que não receberam o composto orgânico. Os novos modelos de crescimento apresentaram ajustes semelhantes aos que serviram de base para as derivativas. Com a inclusão de novos modelos de crescimento derivados de modelos existentes, constatou-se que os seus resultados são semelhantes aos que serviram de base para as derivativas, entretanto, mostra que o estudo de modelagem é dinâmico e nunca se pode prever se o que já existe é suficiente, sobretudo, quando se considera outros campos nas ciência biológica, que podem gerar novos modelos baseados em hipóteses biológicas específicas. REFERÊNCIAS BIBLIOGRÁFICAS
Copyright 2007 - Ciência Florestal The following images related to this document are available:Photo images[cf07021f1b.jpg] [cf07021t2.jpg] [cf07021f1a.jpg] [cf07021t1.jpg] [cf07021f2a.jpg] [cf07021f2b.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

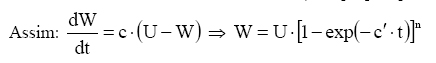
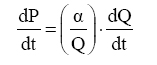
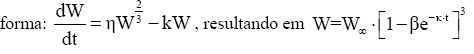
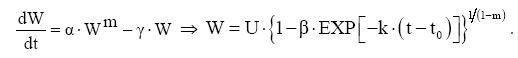
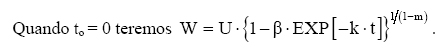 .
. 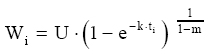
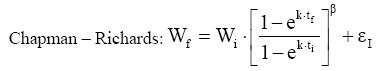
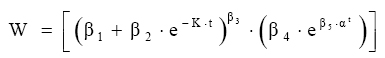
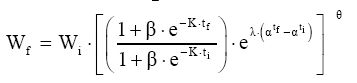
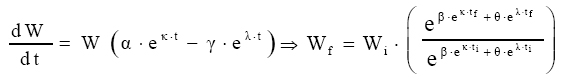
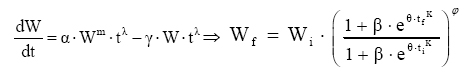
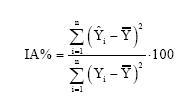
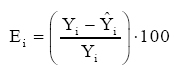
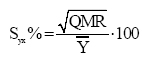
{kind=link}
{kind=link}
{kind=link}