 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ciência Florestal, Vol. 18, No. 1, Jan-Mar, 2008, pp. 47-63 Eucalyptus sp. Seedling Response To Potassium Fertilization And Soil Water1 Matéria Seca E Relações Hídricas Em Mudas De Eucalyptus sp. Em Função Da Fertilização Potássica E Da Umidade Do Solo Paulo César Teixeira2, José Leonardo Moraes Gonçalves3, José Carlos Arthur Junior4 , Cleci Dezordi5 Code Number: cf08005 ABSTRACT A considerable portion of Brazil‘s commercial eucalypt plantations is located in areas subjected to periods of water deficit and grown in soils with low natural fertility, particularly poor in potassium. Potassium is influential in controlling water relations of plants. The objective of this study was to verify the influence of potassium fertilization and soil water potential (Ψw) on the dry matter production and on water relations of eucalypt seedlings grown under greenhouse conditions. The experimental units were arranged in 4x4x2 randomized blocks factorial design, as follow: four species of Eucalyptus (Eucalyptus grandis, Eucalyptus urophylla, Eucalyptus camaldulensis andhybrid Eucalyptus grandis x Eucalyptus urophylla), four dosages of K (0, 50, 100 and 200 mg dm-3) and two soil water potentials (-0.01MPa and -0.1 MPa). Plastic containers with 15 cm diameter and 18 cm height, with Styrofoam base, containing 3.0 dm3 of soil and two plants per container were used. Soil water potential was kept at –0.01MPa for 40 days after seeding. Afterward, the experimental units were divided into two groups: in one group the potential was kept at -0.01MPa, and in the other one, at -0.10 MPa. Soil water potential was controlled gravimetrically twice a day with water replacement until the desired potential was reestablished. A week before harvesting, the leaf water potential (Ψ), the photosynthetic rate (A), the stomatal conductance (gs) and the transpiration rate were evaluated. The last week before harvesting, the mass of the containers was recorded daily before watering to determine the consumption of water by the plants. After harvesting, total dry matter and leaf area were evaluated. The data were submitted to analysis of variance, to Tukey's tests and regression analyses. The application of K influenced A, gs and the transpiration rate. Plants deficient in K showed lower A and higher gs and transpiration rates. There were no statistical differences in A, gs and transpiration rates in plants with and without water deficit. The addition of K reduced the consumption of water per unit of leaf area and, in general, plants submitted to water deficit presented a lower consumption of water. Keywords:potassium nutrition; eucalypt; water deficit, water relations. RESUMO No Brasil, uma considerável porção dos plantios comerciais de eucalipto está localizada em regiões sujeitas a períodos de déficits hídrico (DH) e, normalmente, em solos de baixa fertilidade natural, particularmente pobres em K, o que pode comprometer o crescimento e as relações hídricas das plantas. Assim, este trabalho teve como objetivo verificar a influência da adubação potássica e do potencial de água do solo (Ψw) na produção de matéria seca e nas relações hídricas de mudas de eucalipto, em casa de vegetação. O experimento foi constituído de 32 tratamentos, compreendendo quatro espécies de eucalipto (Eucalyptus grandis, Eucalyptus urophylla, Eucalyptus camaldulensis e o híbridoEucalyptus grandis x Eucalyptus urophylla), quatro doses de K (0, 50, 100 e 200 mg dm-3 de K) e dois regimes de umidade do solo (-0,01MPa e -0,10 MPa), no delineamento em blocos casualizados, em esquema fatorial, com três repetições. Foram utilizados vasos de PVC, com fundo de isopor, contendo 3,0 dm3 de solo, com duas plantas por vaso. O Ψw foi mantido em -0,01MPa por 40 dias. Após, as unidades experimentais foram divididas em dois grupos, sendo o Ψw, em um deles, mantido a -0,01MPa e, no outro, a -0,10 MPa. O controle da umidade do solo foi feito gravimetricamente. Uma semana antes da colheita do experimento, foram avaliados o potencial da água da folha (Ψ), a taxa fotossintética (A), a condutância estomática (gs) e a taxa de transpiração. Durante a última semana de condução do experimento, a massa dos vasos foi anotada diariamente antes da colocação de água de modo a se determinar o consumo de água pelas plantas. Na colheita, as plantas foram coletadas e determinada a matéria seca. Também, foi determinada a área foliar. Os dados foram submetidos à análise de variância, a testes de Tukey e análises de regressão. A aplicação de K influenciou a A, a gs, a transpiração e a EUA. Plantas deficientes em K apresentaram menor A e maior gs e taxa de transpiração, com consequente menor EUA. Não houve diferenças estatísticas na A, gs e taxa de transpiração entre plantas com e sem DH. A adição de K diminuiu o consumo de água por unidade de área foliar e, em geral, plantas submetidas à DH apresentaram menor consumo de água. Palavras-chave: nutrição potássica; eucalipto; déficit hídrico; relações hídricas. INTRODUCTION The development of response curves of Eucalyptus sp growth for K fertilization and the determination of the nutritional efficiency of Eucalyptus species when they grow under a range of K fertility is of practical importance. Data will allow in order to obtain a rational K fertilizer recommendation for field and nursery culture, as well as selecting planting sites for the species. Plants are highly dynamic systems and are exposed to spatially and temporally varying environmental conditions. Plants have developed protection mechanisms against unfavourable fluctuations and efficient mechanisms to use the heterogeneous conditions of their environment, from which they obtain their resources (SCHURR et al., 2006). Plants are often subjected to periods of soil and atmospheric water deficit during their life cycle. In general, reduction in the stored water (plant water stress) can reduce the gas exchange by the leaf and reduce the available water for the tree growth. Consequently, it may modify the partitioning of carbon by the plant and favour the root growth (SHULZE, 1986). Plant water loss is tightly balanced with water uptake to maintain beneficial water status. The most important control on water transport is the change of stomatal aperture, which governs water diffusion from the leaf interior to the atmosphere, as well as the opposite flow of carbon dioxide into the photosynthesizing mesophyll. To balance transpiration and photosynthesis, guard cells may sense and integrate many environmental as well as physiological signals related to photosynthesis, the transpirational demand of the atmosphere, and the plant's current hydraulic status (BUCKLEY, 2005; ROELFSEMA and HEDRICH, 2005). Plant responses to water scarcity are complex, involving deleterious and/or adaptive changes, and under field conditions these responses can be synergistically or antagonistically modified by the superimposition of other stresses (CHAVES et al., 2002). Stomata respond to a variety of environmental and endogenous signals (DODD, 2003; ROELFSEMA and HEDRICH, 2005), and water deficiency is one of the most widely-studied signals. The water stress tolerance varies with the genotype. Some plants have adaptation mechanisms, becoming more efficient in absorption of soil water. One such adaptation is the development of an extensive and deep root system. Other adaptation are changes in leaf area and fast stomata closing under stress (LI et al., 2000; CHAVES et al., 2004, REIS et al., 2006). The maintenance of the growth and gaseous exchange requires the maintenance of a positive leaf turgor. Among plants species, examples of turgor maintenance is accomplished through the adjustment of the cellular elasticity (SOBRADO, 1986;WHITE et al., 1996) and reduction of the leaf’s osmotic potential (CLAYTON-GREENE, 1983; MYERS and NEALES, 1986). The leaf water relationship is a good indication of the capacity of the Eucalyptus species to maintain growth, engage in gas exchange and water use. When the supply of water in the soil is limited, turgor loss and wilting are typical symptoms of K deficiency (MARSCHNER, 1995). The low sensitivity of plants that are well-supplied with K is related to several factors: (a) the role of K+ in stomatal regulation, which is the main mechanism controlling the water regime of plants and (b) the importance of K+ in regulation of the osmotic potential in the vacuole, keeping high water content in tissues under drier conditions. Low biomass production and yield sensitivity to water stress can result from higher concentrations of K+ in the stroma and consequent higher photosynthesis rates (MARSCHNER, 1995). The objective of this work was to verify the influence of potassium fertilization and soil water potential (Ψw) on dry matter accumulation, nutrient accumulation, and water relations in Eucalyptus seedlings grown under greenhouse conditions. MATERIAL AND METHODS This experiment was carried out in the greenhouse of the Department of Forest Sciences, at ESALQ/USP, using 15 cm diameter and 18 cm height plastic containers with a styrofoam base that held 3.0 dm3 of soil. The experiment was comprised of 32 treatments that used four Eucalyptus species (Eucalyptus camaldulensis Dehn, Eucalyptus grandis Hillex Maiden, Eucalyptus urophylla ST Blakeandhybrid Eucalyptus grandis x Eucalyptus urophylla), four dosages of K (0, 50, 100 and 200 mg dm-3) and two soil water potentials: -0.01MPa (without water deficit) and -0.10 MPa (with water deficit). The experimental design was a 4 x 4 x 2 factorial design with three replications in randomized blocks, utilizing two plants per container. Samples of a dystrophic Red-Yellow Oxisol from the Experimental Station of Itatinga, ESALQ/USP, in Itatinga, SP, were used as the soil material. The soil samples were collected from the surface 0-20 cm, homogenized, dried and sieved through a 2-mm mesh sieve. Chemical and physical analyses were made according to EMBRAPA (1997) (Table 1). TABLE 1: Chemical and physical attributes of the 0-20 cm depth of a dystrophic Red-Yellow Oxisol. TABELA 1: Atributos químicos e físicos de um Latossolo Vermelho-Amarelo na profundidade de 0-20 cm.
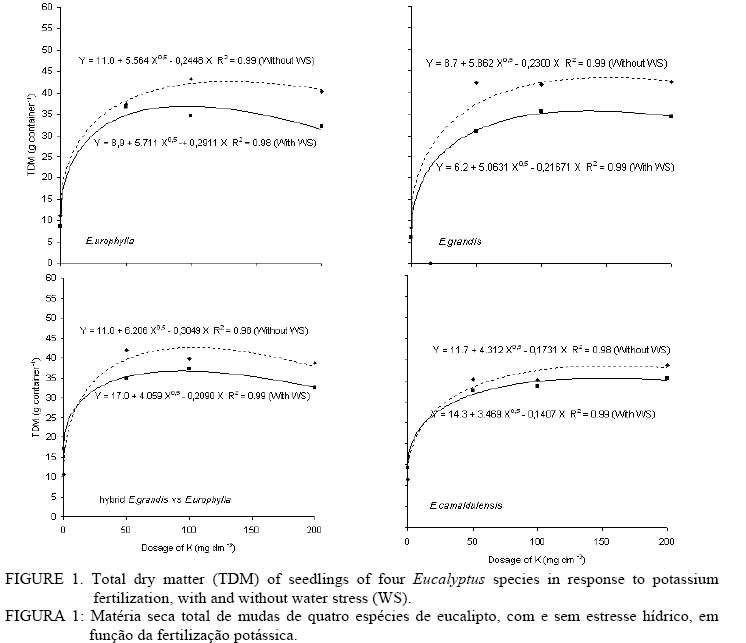
Besides the K applications, the soil received the following additional fertilizer: 100 mg dm-3 of N (50 mg immediately before seeding and 50 mg after seeding, parceled twice), 300 mg dm-3 of P, 45 mg dm-3 of S, 0.82 mg dm-3 of B, 4.0 mg dm-3 of Zn, 3.66 mg dm-3 of Mn, 1.55 mg dm3 of Fe, 1.39 mg dm-3 of Cu and 0.20 mg dm-3 of Mo. The nutrient sources were: KNO3, KCl, (NH4)2SO4, Ca(H2PO4)2.H2O, H3BO3, ZnSO4.7H2O, MnCl2.4H2O, FeCl3.6H2O, CuSO4.5H2O and NaMoO4.2H2O. Except for N and S, all the nutrients were applied before seeding. Approximately 20 seeds were sowed per container of treated soil. Ten days after the emergence, thinning was done, leaving two plants per container. Initially, all the plants were grown with Ψw kept near -0.01MPa, for 40 days. After that period, the containers were divided in two groups. In the first group, Ψw was kept close to -0.01MPa (without water stress). The second group was subjected to a one-week transition period of adaptation when Ψw was gradually reduced to -0.10MPa (with water stress). At the end of this phase, the second group was kept at approximately -0.10 MPa of Ψw. The regulation of Ψw was made gravimetrically. As the plants grew, estimates of the fresh biomass were made to correct the mass of the container in order to keep it at the desired w. One week before harvesting, the leaf water potential (Ψ), the photosynthetic rate (A), the stomatal conductance (gs) and the transpiration rate of the first completely expanded young leaves and exposed to the sun were measured. The Ψ was measured using the pressure bomb (SCHOLANDER et al., 1965) with precautions described by Turner (1988). The gs and A were evaluated using a photosynthesis analyzer (model LICOR LI-6400), on the adaxial surface of the leaves. The evaluations were carried out at 10 AM, 1 PM and 5 PM. The Ψ was also measured at 4 AM (predawn - pd). Eucalyptus camaldulensis was not included in this part of the study, nor the treatments of the other species with a dosage of 200 mg K dm-3.. In the last week before the harvest, the mass of the containers was recorded daily before watering to determine the consumption of water by plants. At the end of the experiment, the soil of each container was sifted using a 0.2 mm mesh sieve and placed on a table. The roots were visually separated. After washing, the roots were dried on paper towels. The vegetative material was collected, and dried for 72 h in an air forced circulation oven at 70ºC. The processing of samples followed the methods of Malavolta et al. (1997). Leaf area was determined by a leaf area meter “AP - ΨT – Devices” (Li-3100 area meter). The data were submitted to a variance analysis of a factorial design. The models of linear regression were adjusted to a square root base. The averages were compared by Tukey tests. In the adjusted models, the dosage of applied K was the independent variable and the dry matter, the leaf area and the water consumption per unit of leaf area were the dependent variables. RESULTS AND DISCUSSION The analysis of variance showed significant differences for the WS, dosage of K and for dosage of K x WS, dosage of K x species and WS x species interactions (Table 2). Total dry matter (TDM) increased upon addition of K for all Eucalyptus species, but decreased with water stress (WS) (Figure 1). In general, TDM did not vary statistically among the species (Table 2) and for all the species, and using equations presented in Figure 1, the estimated maximum TDM was higher for plants without WS. The estimated maximum dry matter production was reduced by WS by 13%, 35%, 14% and 7%, respectively in Eucalyptus urophylla, Eucalyptus grandis, hybrid Eucalyptus grandis x Eucalyptus urophyllaand Eucalyptuscamaldulensis (Figure 1). Nautiyal et al. (1994) also reported a reduction in TDM in eucalyptus plants under WS. These data are consistent with field experience, because Eucalyptuscamaldulensis is one of the most tolerant species to drought (REIS and HALL, 1987). The increase in TDM upon addition of K was higher because the increment in production comparing TDM obtained in a 0 mg dm-3 dosage and the maximum estimated by the regression was, on average, 399%, 552%, 285% and 290%, respectively for Eucalyptusurophylla, Eucalyptusgrandis, the hybrid Eucalyptusgrandis xEucalyptusurophylla and Eucalyptuscamaldulensis. TABLE 2: Analysis of variance results for total dry matter (TDM) and leaf area (LA). TABELA 2: Análise de variância relativa a matéria seca total e área foliar.
In that: * *, * and (ns) indicate significance to 1 and 5% and no significance to 5% of probability, respectively. Teixeira et al.(1995) verified that plant height and TDM of nine hybrids of eucalyptus were significantly affected by the water status and K. These authors reported that the variation in observed values among the hybrids reflected the genetic variability of the material. According to Barros et al. (1990) and Cicolim et al. (2002), the increase in growth due to fertilization varies among the different eucalypts species. The analysis of variance for leaf area (LA) was significant for the main factors and for the interactions of dosage of K x WS and the dosage of K x species (Table 2). In general, the leaf area was less in Eucalyptuscamaldulensis than in the other species (Table 3). In general, Eucalyptusurophylla and Eucalyptusgrandis showed a reduced leaf area with WS (Table 3). The growth due to photosynthesis was expected to be proportional to the leaf area (or weight) (CEULEMANS and SAUGIER, 1991; MILLER, 1995; LAMBERS et al., 1998), therefore, the gain in carbon may be sacrificed when the leaf area decreases in response to water and nutritional deficiencies. TABLE 3: Leaf area of seedlings of four eucalypts species in response to potassium fertilization, with and without water stress (WS). TABELA 3: Área foliar de mudas de quatro espécies de eucalipto, com e sem déficit hídrico, em função da fertilização potássica.
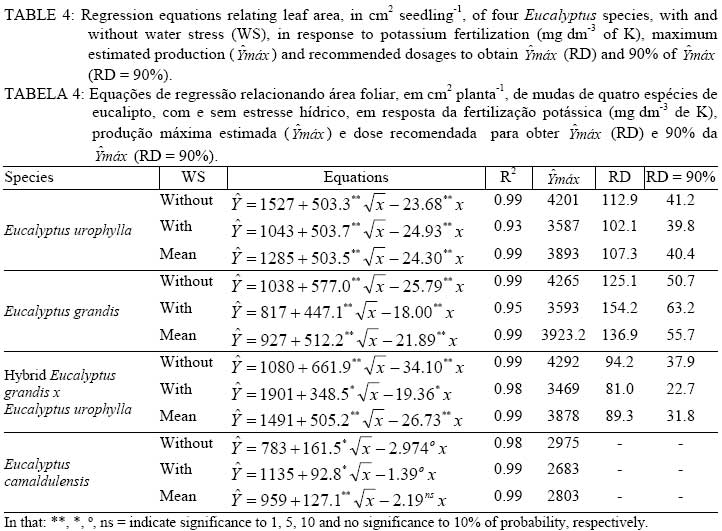
In that: 1 / -0.01 MPa;2 / -0.10 MPa; 3/ Averages followed by the same capital letter, in the column, did not differ at p<0.05 by the Tukey test. The estimated maximum leaf area of all the species was always larger in plants without WS, compared to plants with WS (Table 4). There was a reduction in the maximum estimated leaf area of 14.6%, 15.7%, 19.2% and 9.8%, respectively in Eucalyptus urophylla, Eucalyptusgrandis, hybrid Eucalyptus grandis x Eucalyptus urophyllaand Eucalyptuscamaldulensis with WS when compared with that obtained in plants without WS. In these species, the gains in the leaf area with the application of K, if compared with the leaf area obtained with a dosage of 0 mg dm-3 and the maximum estimated dosage, were 243%, 434%, 259% and 296%, respectively. Potassium fertilization was a factor that strongly influenced the photosynthetic rate (A) of all Eucalyptus species (Table 5). In spite of the lack of statistical differences for A, when different K dosages were used 50 and 100 mg K dm-3 tended to be higher than control in almost all the situations. In Eucalyptus grandis without WS, there was a reduction in A at 1 PM with 50 mg K dm-3, and there was no difference in A in relation to WS. Only Eucalyptus grandis plants with WS showed significant reduction in A when 50 mg K dm-3 were added. Reduction of growth observed in plants with WS was due mainly to reduction in leaf area and not due to reduction in A. On the other hand, the obtained advantage in dry matter production with K fertilization may be due to increased leaf area and resulting in enhanced total photosynthetic assimilation. TABLE 5: Photosynthetic rate of seedlings of three eucalypts species, with and without water stress (WS), at different times of the day, in response to potassium fertilization. TABELA 5: Taxa fotossintética de mudas de três espécies de eucalipto, com e sem estresse hídrico, em diferentes horários do dia, em resposta da fertilização potássica.
In that: For every time and species, averages followed by the same small letter, in the column, did not differ amongst themselves for the Tukey test at p<0.05; for each species, averages followed by the same capital letter, in the line, did not differ amongst themselves. The smallest variations in the applied dosages of K and WS were observed at 5 PM (Table 5). There was no significant difference in any of the studied factors. In general, there were no differences in the species at any of the times evaluated (Table 5). Larcher (1995) and Lambers et al. (1998) reported that the reduction of growth when w is low was due to the reduction of the gs and A, smaller leaf elongation and protein synthesis. When the supply of nutrients is not enough the leaf growth rate as well as the leaf area may be limited by low rates of net photosynthesis or insufficient leaf expansion or both (MARSCHNER, 1995). In contrast to the response of A, plants deficient in K showed high gs, especially at 10 AM and 1 PM (Table 6), and a high transpiration rate (Table 7). In general, the maintenance of the opened stomata in plants that did not receive application of K (thus allowing higher exchange of CO2 with the atmosphere) was not enough to increase A. The flow of CO2 through the stomata is less affected by the partial stomata closure than by the flow of water outside of the leaf (NOBEL, 1991). Therefore, A and gs may not always be highly correlated, although they usually seem to be (MEINZER et al., 1984; TESKEY et al., 1994). The largest gs values for the 0 mg K dm-3 treatment were observed at 10 AM and 1 PM. With the application of K, the gs decreased (Table 6) reducing the excessive loss of water by transpiration (Table 7). For the 0 mg K dm-3 treatment, Eucalyptus urophylla and the hybridshowed larger gs at 10 AM than when under WS, indicating that when under acute deficiency of K these species lose the capacity to control the loss of water by the stomata. In average, plants of Eucalyptus grandis and hybrid Eucalyptus grandis x Eucalyptus urophylla without WS tended to have higher gs (Table 6) than plants with WS, especially at 10AM and 1 PM. Thus, as was observed for A, the smallest gs values and transpiration rates were obtained at 5 PM. TABLE 6. Stomatal conductance of seedlings of three eucalypts species, with and without water stress (WS), at different times of the day, in response to potassium fertilization. TABELA 6: Condutância estomática de mudas de três espécies de eucalipto, com e sem estresse hídrico, em diferentes horários do dia, em resposta da fertilização potássica.
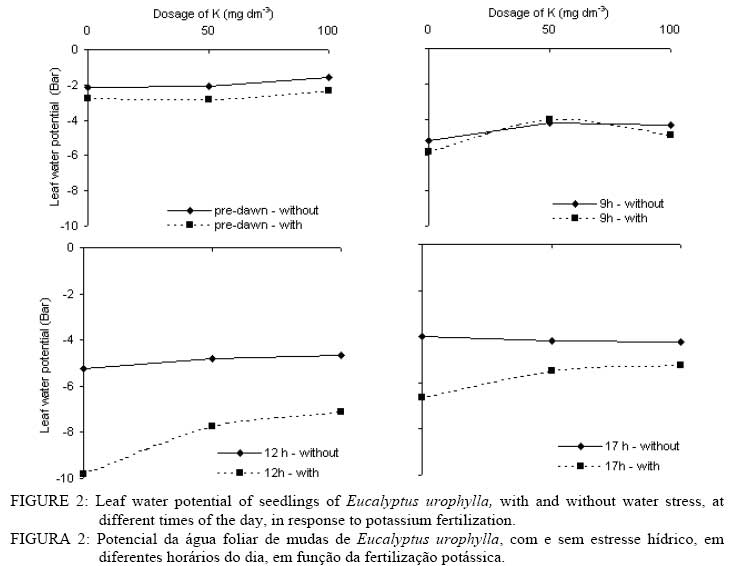
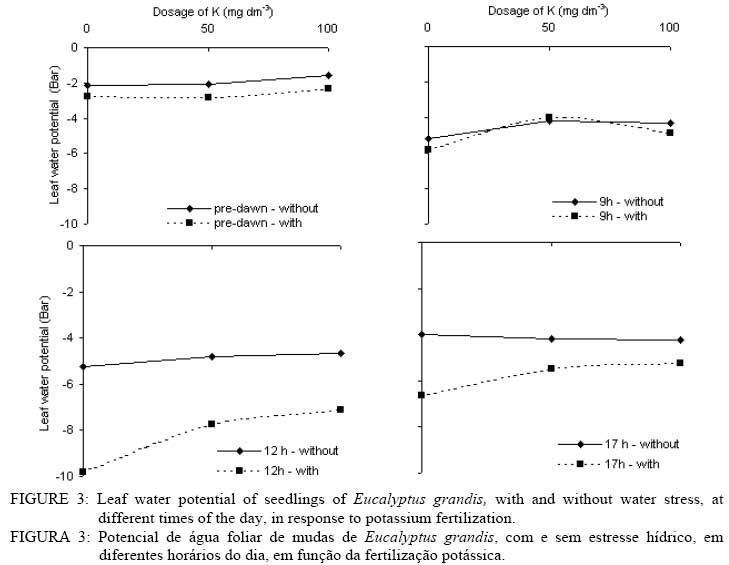
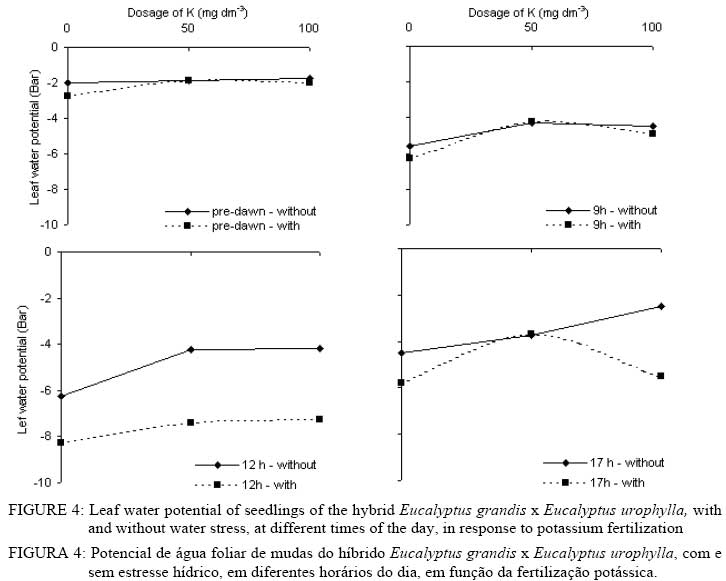
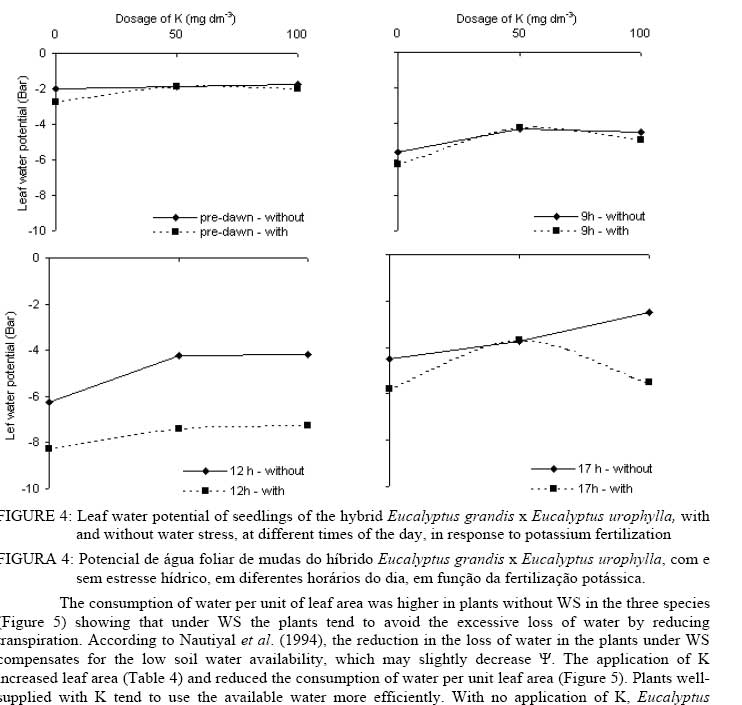
In that: For every time and species, averages followed by the same small letter, in the column, did not differ amongst themselves for the Tukey test at p<0.05; for each species, averages followed by the same capital letter, in the line, did not differ amongst themselves. In spite of no statistical difference in most treatments, the highest gs values were found in plants without WS. This shows that in the studied species there was a tendency to avoid an excessive loss of water when under WS. The idea that the plants adjust leaf area better than the gs in response to the climate has become popular in the literature (eg., SPETCH, 1972; EAGLESON, 1982; HATTON and HU, 1995). As observed for A, the gs did not show any difference among the species in at of the sampling times. Some species are capable of experiencing accentuated water deficiency without totally closing their stomata (high stomatal conductance) and maintaining CO2 fixation (TEIXEIRA et al., 1995). Also, early production of thick, narrow, petiolate, vertically oriented, isobilateral, amphistomatal adult leaves by seedlings appears to be related to the stressful conditions within its local habitat (JAMES and BELL, 2001) and it is possible that the differences in the tolerance to water deficit among eucalyptus species may be due to a larger or a smaller number of stomata in the abbatial and adaxial leaf surface, but this is not clear. The regulation of gas exchange at the leaf level is a key factor for plant survival under a fluctuating environment (BUCKLEY, 2005). The gs values for the three species at 10 AM and 1 PM were always higher without application of K. At these times the differences in the gs were not verified for dosages of 50 and 100 mg dm-3 in any of the species. At 5 PM no statistical difference was evident for all treatments. The values for the transpiration rate followed the same behavior observed for gs (Tables 6 and 7). In general, for all species, the highest transpiration rates were observed in the treatments without K fertilization and the lowest in treatments with K. There was also, in most cases, no statistical difference in transpiration rates between plants with and without WS; however, it is probable that if the water deficit were higher, differences might be found in plants with and without WS. Eucalyptus grandis, at 10 AM and the hybrid, at 10 AM and 1 PM, showed low values of transpiration rate when under WS in treatments where there was addition of K.The higher plant K was a fundamental factor in the control of water loss from seedlings of the three species. Only at the end of the afternoon (5 PM) there was no difference in loss of water among the treatments. In this case, it is probable that plants deficient in K were slower to close e stomata. The sensitivity to stomata closure was greater in plants supplied with K. There was no statistical difference in the transpiration rate among the species at any of the measurement times; however, higher mean values were observed in the hybrid. Leaf water potential (ψ) was different among treatments (Figures 2, 3 and 4). Lower values were measured at predawn (Ψpd) in plants subject to WS. TABLE 7: Transpiration rate of seedlings of three eucalyptus species, with and without water stress (WS), at different times of the day, in response to potassium fertilization. TABELA 7: Taxa de transpiração de mudas de três espécies de eucalipto, com e sem estresse hídrico, em diferentes horários do dia, em resposta da fertilização potássica.
In that: For every time and species, averages followed by the same small letter, in the column, did not differ amongst themselves for the test of Tukey; for each species, averages followed by the same capital letter, in the line, did not differ amongst themselves. Stoneman et al. (1994) verified, in Eucalyptus marginata, that leaf growth rate decreased linearly with decreases in Ψpd. Eucalyptus grandis was the species that presented the smallest differences in Ψpd, in plants with and without WS for dosages of 50 and 100 mg dm-3 of K (Figure 4). Of the three species studied, Eucalyptus urophylla showed the highest reduction of Ψ at 1 PM when under WS. Some authors suggest that Ψpd can be used to infer on Ψw while others do not agree (eg., Begg and Turner, 1970; White et al., 2000). During the day, the lowest values of Ψ were observed in treatments with no application of K. The largest differences in the Ψ between plants with and without WS were observed at 1 PM in all the species. In general, in almost all the treatments, the application of K increased slightly the Ψ in all the species. Plants without WS presented higher, or at least the same, values at every measurement time. The smallest values of Ψwhich were being found in plants without WS can be explained by the higher gain in leaf area found in all the species of plants with WS, because higher turgidity favors leaf expansion. The consumption of water per unit of leaf area was higher in plants without WS in the three species (Figure 5) showing that under WS the plants tend to avoid the excessive loss of water by reducing transpiration. According to Nautiyal et al. (1994), the reduction in the loss of water in the plants under WS compensates for the low soil water availability, which may slightly decrease Ψ. The application of K increased leaf area (Table 4) and reduced the consumption of water per unit leaf area (Figure 5). Plants well-supplied with K tend to use the available water more efficiently. With no application of K, Eucalyptus grandis was the species that showed the lowest consumption of water when under WS, but, in the plants without WS, Eucalyptus urophylla showed the lowest values. In general, the three species had similar values of consumption of water per unit of leaf area (Figure 5). The highest variations were found between plants with and without WS. Different species can transpire at the same rate per unit of leaf area (EAGLESON, 1982; HATTON et al., 1998), however studies have shown great inter-specific variation in the gs (eg., ATTIWILL and CLAYTON-GREENE, 1984). Differences in stomatal conductance cannot explain the different transpiration rates due to variation in conductance in the leaf boundary layer (MEINZER, 1993) and decoupling degree (McNAUGHTON and JARVIS, 1983; JARVIS and McNAUGHTON, 1986) in the canopy. CONCLUSIONS Total dry matter increased upon addition of K for all Eucalyptus species, and decreased with WS. Reduction of growth observed in plants with WS was due mainly to reduction in leaf area and not due to reduction in A. On the other hand, the obtained advantage in dry matter production with K fertilization was due to increased leaf area and resulting in enhanced total photosynthetic assimilation. In average, plants of Eucalyptus grandis and hybrid Eucalyptus grandis x Eucalyptus urophylla without WS tended to have higher stomatal conductance than plants with WS. In general, for all species, the highest transpiration rates were observed in the treatments without K fertilization and the lowest in treatments with K The consumption of water per unit of leaf area was higher in plants without WS in the three species compared to plants with WS. ACKNOWLEDGEMENTS The authors thank Ivan Mateus Moura, Luciane Stoppa, Thais Dias Gonçalves, Jarede, Atus, “Seu Chico”, Marcos Wichert and Jean Paul Laclau for their valuable help in data collection and for the friendship. Acknowledgment is also due to Nicholas Brian Comerford for revision of the paper. BIBLIOGRAPHICAL REFERENCES
Copyright 2008 - Ciência Florestal The following images related to this document are available:Photo images[cf08005t7.jpg] [cf08005t3.jpg] [cf08005t6.jpg] [cf08005f4.jpg] [cf08005f1.jpg] [cf08005f2.jpg] [cf08005t1.jpg] [cf08005t2.jpg] [cf08005t4.jpg] [cf08005f5.jpg] [cf08005t5.jpg] [cf08005f3.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}