 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ciência Florestal, Vol. 18, No. 2, 2008, pp. 233-245 FLORÍSTICA E CLASSIFICAÇÃO DA REGENERAÇÃO NATURAL EM FLORESTA OMBRÓFILA MISTA NA FLORESTA NACIONAL DE SÃO FRANCISCO DE PAULA, RS1 FLORISTIC AND CLASSIFICATION OF THE NATURAL REGENERATION IN THE MIXED RAINY FOREST AT NATIONAL FOREST OF SÃO FRANCISCO DE PAULA, RS Igor da Silva Narvaes2, Solon Jonas Longhi3, Doádi Antônio Brena4 1Parte da Dissertação intitulada Classificação e Caracterização da Regeneração Natural em Floresta Ombrófila Mista
na Floresta Nacional de São Francisco de Paula, RS, apresentada no Programa de Pós-graduação em Engenharia
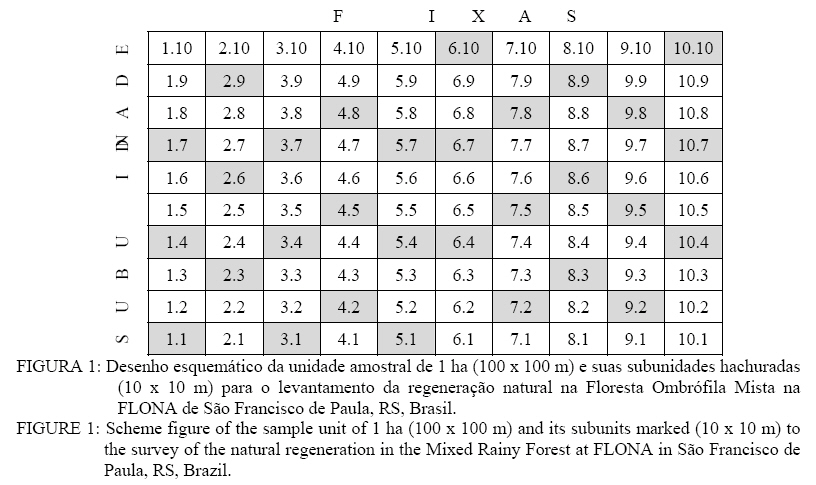
Florestal. Recebido para publicação em 2/02/2006 e aceito em 2/06/2008. Code Number: cf08021 RESUMO Na floresta Ombrófila Mista, a regeneração natural é pouco estudada. No Rio Grande do Sul, em conseqüência da ação antrópica, essas florestas se encontram sob diferentes condições de alteração, em alguns casos formando fragmentos. O estudo teve como objetivo analisar a florística da regeneração natural e verificar a ocorrência de grupos florísticos de uma área onde não foram constatadas alterações intensas. A área com 1606,69 ha localiza-se na Floresta Nacional de São Francisco de Paula (entre 29° 23’ e 29° 27’ S; 50° 23’ e 50° 25’ W), no município de São Francisco de Paula, RS. No estudo, foram selecionadas seis parcelas permanentes de crescimento do PELD (Projeto Ecológico de Longa Duração) cuja vegetação foi inventariada, utilizando-se dez faixas paralelas entre si e perpendiculares à direção da posição topográfica, as quais apresentaram 10 m de largura e 100 m de comprimento, subdivididas em dez unidades de 100 m² onde foram sorteadas três unidades amostrais por faixa para identificação e medição (altura e circunferência) de indivíduos com Cap maior ou igual a 3 cm e menor de 30 cm. Os cipós, trepadeiras e ervas também tiveram suas circunferências medidas e identificados até o nível de espécie, quando possível. Os dados de densidade por espécie formaram uma matriz (180x70) utilizada na análise multivariada. A presença de agrupamento de vegetação em estado de regeneração natural no interior da floresta foi testada pelo Método TWINSPAN (Two-way indicator species analysis). Constatou-se a existência de três grupos ecológicos (G-1, G-2 e G-3). O G-1 foi caracterizado por boa disponibilidade hídrica, por possuir características ambientais intermediárias e por situar-se na posição topográfica referente à encosta média; G-2 ocorreu na encosta inferior da floresta, apresentando forte influência da umidade e sujeita a alagamentos esporádicos; e G-3, na posição topográfica da encosta superior/platô, que, por conseguinte, não sofre influência da elevação da umidade. As espécies indicadoras dos grupos foram: Sebastiania brasiliensis Spreng. (G-1); Matayba elaeagnoides Radek., Myrceugenia myrcioides Cambess O. Berg, Myrceugenia oxysepala (Burret) D. Legrand et Kausel e Cinnamomum glaziovii (Mez) Kosterm. (G-2); e Zanthoxylum petiolare A. St.-Hil & Tul. (G-3). Palavras-chave: análise multivariada; agrupamento da vegetação; grupos ecológicos. ABSTRACT In Mixed Rainy Forest, natural regeneration is not studied very well. In Rio Grande do Sul, due to anthropic actions, it is under different alteration conditions, in some cases, forming fragments. This study aimed to analyze the floristic of natural regeneration and to verify the occurrence of floristic groups in the area where no pronounced alterations were found. The area with 1606,69 ha was located at São Francisco de Paula National Forest (among 29° 23 ' and 29° 27 ' S; 50° 23' and 50° 25 ' W), displayed in 1.606,69 ha, in São Francisco de Paula municipal county, RS. In the study, 6 permanent growing stands of PELD (Long Duration Ecological Project) were selected, the vegetation was inventoried, using 10 parallel strips to each other and perpendicular to the direction of the topographical position, which presented 10 m of width and 100 m of length, subdivided into 10 units of 100 m², totalizing 1 ha, where were raffled 3 sample units for strip to identify and measure (height and circumference) the individuals with Cap higher or equal to 3 cm and smaller than 30 cm. The lianas, creepers and herbs also had their circumferences measured and identified up to the species level, when possible. The density data for species formed a matrix (180x70) used in the multivariate analysis. The presence of vegetation grouping in state of natural regeneration inside the forest was tested by TWINSPAN Method (Two-way indicator species analysis). The existence of three ecological groups (G-1, G-2 and G-3) was proved. G-1 was characterized by good water availability and for having intermediate environmental characteristics, for locating in the topographical position regarding the medium hillside. G-2 occurred in the base hillside of the forest, presenting strong influence of the hydric saturation and subjected to sporadic overflow; and, in G-3, in the topographical position of the hillside higher/plateau that consequently doesn't suffer influence of the elevation of hydric saturation. The indicative species of the groups were: Sebastiania brasiliensis Spreng. (G-1); Matayba elaeagnoides Radek., Myrceugenia myrcioides Cambess O. Berg, Myrceugenia oxysepala (Burret) D. Legrand et Kausel and Cinnamomum glaziovii (Mez) Kosterm.(G-2); and Zanthoxylum petiolare A. St.-Hil & Tul. (G-3). Keywords: multivariate analysis; vegetation grouping; ecological groups. INTRODUÇÃO O interesse no estudo da regeneração natural em nível mundial vem crescendo muito na última década, especialmente no que diz respeito à elaboração de planos de manejo, pois, de acordo com Amador e Viana (2000), a regeneração natural permite uma análise efetiva para diagnosticar o estado de conservação do fragmento e a resposta ao manejo e, além disso, segundo Daniel e Jankauskis (1989), o entendimento de seus processos regenerativos é importante para o sucesso de seu manejo o qual necessita de informações básicas em qualquer nível de investigação. Muitos estudos vêm sendo realizados sobre o tema, entretanto poucos são referentes à Floresta Ombrófila Mista (Longhi, 1980; Jarenkow, 1985; Calegari, 1999; Mauhs e Backes, 2002), sobretudo levando-se em conta que o referido tipo florestal vem sendo explorado de forma inadequada pela retirada de madeira de valor comercial de forma clandestina, ou pela substituição da floresta para o uso do solo como área de cultivo agrícola, exceto em pequenas áreas onde o relevo não o favorece. Inoue (1979) enumera e elucida alguns fatores responsáveis pela ocorrência das espécies em estado de regeneração natural em maior ou menor intensidade, agindo simultaneamente para o seu sucesso no estabelecimento, formação estrutural e fisionômica da floresta, sendo eles: água, temperatura, germinação, competição, luz e solo. Entretanto, deve-se levar em conta, além dos fatores ambientais, também a biologia das espécies consideradas (Morretes, 1992). De acordo com Dorneles e Negrelle (2000), a densidade e permanência de uma dada população, numa comunidade, dependem de sua capacidade de auto-regeneração e de fatores extrínsecos que lhes são impostos, tais como clima, interações bióticas ou disponibilidade lumínica. Com base nessas informações, verifica-se a importância do estudo da regeneração natural nesse Bioma, estando englobado em um conjunto de estudos e pesquisas previstas no Projeto Ecológico de Longa Duração – PELD/CNPQ – “Conservação e Manejo Sustentável de Ecossistemas Florestais – Bioma Araucária e suas Transições”, em andamento na Unidade Experimental de Pesquisa: Fazenda Tupi, em Nova Prata, RS e na Floresta Nacional de São Francisco de Paula onde o presente trabalho foi realizado. Esse tipo de estudo vêm sendo realizados nas últimas décadas, pois além de caracterizar a composição florística e estrutura fitossocilógica das florestas naturais remanescentes, busca entender sobretudo a sua dinâmica e, com base nos resultados, propor modelos que facilitem a regeneração e sucessão florestal, dando subsídio para a recuperação e manejo dos ecossistemas, como nos estudos de Almeida (1998)e Park (2001),entre outros. A vegetação é influenciada por diversos fatores ambientais formadores dos “gradientes ambientais” na floresta, influenciando direta e/ou indiretamente, avaliados com diversos enfoques e seguindo metodologias das mais variadas, como nos estudos de Diaci (2002); Hubbell et al. (1999); Malcolm et al. (2001), porém não somente levando-se em conta a formação da regeneração natural no seu aspecto florístico, fitossociológico, estrutural como também na formação de grupos florísticos, estes decorrentes dos fatores ambientais (Narvaes, 2004). Assim, o presente trabalho teve como objetivo verificar a ocorrência de grupos florísticos (classificação) na regeneração natural, servindo como base para a obtenção de subsídios para a definição de métodos de manejo florestal da regeneração por meio de indicações práticas, cientificamente comprovadas, tanto para a conservação como na recuperação desse tipo florestal. MATERIAL E MÉTODOS Caracterização da área de estudo O estudo foi realizado na Floresta Nacional (FLONA) de São Francisco de Paula, localizada no Rincão dos Kröeff, no município de São Francisco de Paula, entre as coordenadas 29°23’ e 29°27’S e 50°23’ e 50°25’W, no estado do Rio Grande do Sul. A área total de 1.606,69 hectares está situada na zona de transição entre Floresta Ombrófila Densa e Floresta Ombrófila Mista, dos quais 901,9 ha são de floresta nativa, cobrindo aproximadamente 56% da área total da FLONA, na qual existem ainda reflorestamentos de Araucaria angustifolia, Eucayptus spp. e Pinus spp., possuindo relevo predominantemente ondulado, de acordo com Dobrovolski et al. (2006) com 7,1% da área total com declividade superior a 30°. O solo é do tipo Cambissolo Húmico Alumínico típico (STRECK et al., 2002). Os valores médios anuais de temperatura e precipitação são, respectivamente, inferiores a 18,5oC e 2468 mm (Moreno, 1961), e com clima classificado de acordo com a classificação de Köppen do tipo “Cfb”, mesotérmico médio, dominado por cotas altimétricas entre 1.000 e 1.100m no norte do estado em questão. Amostragem e coleta dos dados Tomando-se como base as dez unidades amostrais demarcadas anteriormente na floresta pelo Projeto PELD (Projeto Ecológico de Longa Duração) – Conservação e Manejo Sustentável de Ecossistemas Florestais, foram selecionadas seis unidades amostrais, adotando-se, como critério de seleção, as características de declividade, a posição topográfica, as características físicas do solo e a exposição do terreno. Dentro de cada unidade amostral, compostas de dez faixas paralelas (10x100m), foram sorteadas três subunidades de 10 x 10 m (100 m²), totalizando trinta subunidades por hectare, conforme detalhada na Figura 1: Nas subunidades, foram mensurados todos os indivíduos com altura superior a 1,30 m com circunferência à altura do peito (Cap) mínima de 3,0 cm, até a circunferência-limite de 30,0 cm, dos quais foram anotadas as seguintes informações: nome comum; altura total, circunferência a altura do peito; forma de vida: árvore, palmeira, arbusto, herbácea, trepadeira e cipó, conforme descrição do Inventário Florestal Contínuo do Rio Grande do Sul (RIO GRANDE DO SUL, 2002). A identificação, em nível de família, gênero e espécie, foi obtida por meio de consulta ao Herbário do Departamento de Ciências Florestais (HDCF), da Universidade Federal de Santa Maria, em caso de não-identificação, em um primeiro estágio, enviadas ao herbário da Universidade Federal de Minas Gerais. Método de análise A determinação de grupos florísticos na Floresta Ombrófila Mista da FLONA de São Francisco de Paula foi realizada pelo método de classificação TWINSPAN (“Two Way Indicator Species Analysis”) o qual verifica a ocorrência de padrões na distribuição de espécies, associados às condições ambientais do local, constatadas no campo (Hill, 1979). De acordo com Hill (1979), é um método divisivo, hierárquico e politético, desenvolvido e discutido em um contexto fitossociológico em que a matriz de dados é construída pela abundância das espécies nas parcelas, sendo aplicável para uma ampla gama de matrizes de dados, expressando seus atributos individuais. A matriz utilizada referente a esse método de classificação foi analisada pelo programa PC-ORD (Mc Cune e Mefford, 1995;1997). Segundo o mesmo autor, cada divisão gera um autovalor que indica o percentual de variação dos dados da comunidade no eixo de ordenação. Normalmente, um autovalor em torno de 0,30 é considerado significativo para explicar as variações ocorrentes na natureza, logo quando a divisão atingir um autovalor menor que 0,30, esta poderá ser descartada (Kent e Coker, 1992). Na análise multivariada, para a regeneração natural, utilizou-se uma matriz de 180 x 70, levando-se em consideração o total de 180 subunidades amostrais (linhas), setenta espécies (colunas). As espécies com poucos indivíduos foram descartadas da análise, já que apresentam pouca ou nenhuma influência sobre os resultados dos agrupamentos. Assim, as espécies com menos de cinco indivíduos foram desprezadas, embora inventariadas nas parcelas. Trabalhos de Nascimento (2000) e Araujo (2002) consideraram um critério menos rígido, desconsiderando espécies com menos de três indivíduos. Para a execução da análise, os pontos de corte estipulados para as pseudoespécies foram 0, 2, 5, 10 e 20. Assim, conforme Araujo (2002), uma espécie se particiona em uma “série de espécies”, tendo diferentes densidades de cobertura. Por exemplo, a pseudoespécie 1 teria a cobertura de um a dois indivíduos em determinadas parcelas; a pseudoespécie 2 de dois a cinco indivíduos e, assim, sucessivamente, até a pseudoespécie 5 que corresponderia vinte indivíduos na parcela ou mais. Assim, dependendo da densidade de cobertura de cada espécie, ela se encaixasse em determinada pseudoespécie. A escolha do TWINSPAN se baseou em sua maior flexibilidade em relação aos outros métodos, na análise de indicação de espécies, e também por envolver muitos aspectos novos. O mais espressivo desses “aspectos novos” é que o programa primeiro constrói uma classificação para as parcelas, e então a utiliza para obter uma classificação das espécies de acordo com as suas preferências ecológicas, utilizando as parcelas como base (Hill, 1979). As duas classificações (linhas e colunas) são então utilizadas juntas para obter uma tabela de ordenação de duplo caminho “ordered two-way table” que expressa as relações sinecológicas das espécies, construindo o que é denominado por Felfili et al.(2001) de tabela dicotômica. Para a definição dos agrupamentos, além da análise multivariada, levaram-se em consideração as características observadas no campo e possíveis explicações ecológicas para a formação de tais grupos. A saída de dados do TWINSPAN, ou seja, o resultado final se constitui na descrição das divisões realizadas, com respectivos autovalores, conjunto de unidades amostrais e de espécies similares (agrupamentos), espécies indicadoras e preferenciais, amostras malclassificadas, além de uma matriz organizada com a classificação hierárquica das amostras e espécies conjuntamente (Araujo, 2002). RESULTADOS E DISCUSSÃO Classificação e caracterização geral dos grupos florísticos Com base na análise de cluster pelo método de Twinspan, houve a formação de três grupos florísticos, de acordo com a preferência de determinado grupo de espécies em relação às características ambientais existentes na área de estudo. Para a formação de grupos, a preferência ecológica para a ocorrência de espécies em determinado local é determinada pelas espécies indicadoras (HILL et al., 1975). Para a definição dos grupos florísticos, além das espécies indicadoras, as espécies preferenciais (abreviadas) servem também para melhor caracterizar cada grupo, mostrando o número de parcelas onde ocorreu a pseudoespécie. Conforme a Figura 2, o número após a espécie se refere a pseudoespécies (nível de corte), enquanto que os números entre parênteses se referem às parcelas onde estão contidas as pseudoespécies, em ambos lados da divisão. A classificação das unidades amostrais em grupos se deu da seguinte forma: a primeira divisão, com autovalor de 0,2514, embora baixo (< 0,30) indicou haver diferenciação entre as parcelas, segundo a posição topográfica que ocupa no relevo predominante da paisagem, influindo na formação dos outros fatores ambientais. A formação desse grande grupo ecológico demonstrou que existe similaridade florística na vegetação formadora das unidades amostrais da encosta média (Grupo ecológico 1: G-1); e diferença daquelas que ocorrem em posições topográficas extremas (encosta superior/platô e encosta inferior) o que indicou a existência de espécies com diferentes graus de tolerância à umidade do solo e outros fatores que acarretam tais variações de umidade (Figura 2). Na maioria dos casos, as espécies não foram exclusivas, porém ocorreram em maior abundância em um determinado grupo de acordo com sua adaptabilidade as condições ecológicas, com isto tendo preferência a um grupo característico. No estudo de Burgman e Thompson (1982),também foram classificados três grupos de espécies, de acordo com os locais de ocorrência, divididos em “Paisagens secas, Comunidades associadas a ambientes úmidos e comunidades provindas de regiões areníticas”. Entretanto, a maioria dessas espécies, denominadas de espécies diagnósticas, não se mostraram exclusivas, porém ocorreram em maior abundância em determinado grupo, de acordo com a suas preferências ecológicas. Sebastiania brasiliensis Spreng. foi a espécie indicadora do G-1 (Encosta média), ocorrendo em 101 parcelas. Essa espécie é descrita por Smith et al. (1988) como espécie freqüente nos sub-bosques dos pinhais, adaptada a solos úmidos e situadas em pequenas depressões, sendo seletiva higrófila e adaptada à luz difusa. Nesse mesmo grupo, as pseudoespécies Sebst br 1 a 5 (Sebastiania brasiliensis em quatro pontos de corte), Myrc cuc 1 e 2 (Myrceugenia cucullata D. Legrandem dois pontos de corte), Allophylus edulis (A. St-Hil., Cambess. & A. Juss) Radlk. (All edu 1), Allophylus guaraniticus (A. St.- Hil.) Radlk.(All gua1), Campomanesia xanthocarpa O. Berg (Camp xan 1), Maytenus evonymoides Reissek. (Mayt evo 1) Campomanesia rhombea O. Berg (Camp rom2), Ilex paraguariensis A. St.-Hil. (Ilex par 2), ocorrem em mais parcelas pertencentes ao grupo 1 do que nas parcelas do outro lado da divisão. Por outro lado, espécies como Rudgea parquioides (Cham.) Müll. Arg., Lamanonia ternata Vell., Ocotea indecora (Schott) Mez-Rohwer e Casearia decandra Jacq. foram as espécies indicadoras do agrupamento das posições topográficas extremas. Nestas, houve um grande número de espécies preferenciais discutidas posteriormente, constituindo-se no outro grande grupo formado na primeira divisão. Esse mesmo grupo foi novamente dividido, gerando os grupos 2 e 3. A segunda divisão, com autovalor superior ao da primeira (0,2974), demonstrou uma forte separação das unidades amostrais de posições topográficas extremas em dois grupos florísticos, uma na encosta inferior (G-2) e outra na zona de transição encosta superior/platô (G-3) (Figura 2). Embora com autovalor próximo ao proposto na literatura, a divisão formadora destes grupos condizem com as condições ambientais encontradas na área de pesquisa, sendo assim consideradas válidas. Ao se analisar os dados complementares, presentes em Narvaes (2004), nas matrizes dos fatores ecológicos e os relatórios de campo, foi possível estabelecer tais diferenças, sobretudo em relação à posição topográfica em que se encontram as unidades amostrais, sendo que o primeiro grupo ecológico (G-1) se encontra em uma posição topográfica intermediária, o grupo ecológico 2 (G-2) situado na encosta inferior e o terceiro grupo na posição topográfica de transição (G-3) entre a encosta superior e o platô. As inter-relações entre a distribuição de espécies lenhosas e as variáveis solo-topografia já foram observadas por Oliveira-Filhoet al.(1998), na Floresta Seca Decídua em Santa Vitória, MG, entretanto se deve levar em consideração que ao menos em alguns casos os fatores relacionados aos gradientes de luz têm importância principal na sua diferenciação. O grupo ecológico 2 (G-2) diferenciado por representar locais de maior umidade (encosta inferior) teve como espécies indicadoras Matayba elaeagnoides Radlk. (Matay el 1 a 4, com 27, 25, 15 e nove subparcelas respectivamente), Myrceugenia myrcioides (Cambess.) O. Berg (Myrc myr 1 e 2, com 22 e 17 sub-parcelas, respectivamente), Myrceugenia oxysepala (Burret) D. Legrand et Kausel (Myrc oxy 1 a 4, com 26,19,14 e 6 sub-parcelas, respectivamente) e Cinnamomum glaziovii (Mez) Kosterm. (Cin glaz 1 a 3, com 19, 15 e 7 sub-parcelas, respectivamente). Além das espécies indicadoras, as preferenciais foram Blepharocalyx salicifolius(Kunth) O. Berg (Blef sal 1), Cupania vernalis Cambess. (Cupan ve 1 e 2), Matayba elaeagnoides (Matay el 1 a 4), Sebastiania commersoniana (Baill.) L. B. Sm. & Downs(Sebst co 1), Seguieria aculeata Jacq. (Segui ac 1 e 2), Stillingia oppositifolia Baill. ex Müll. Arg. (Still op 1 a 5), Myrceugenia myrcioides (Myrc myr 1 e 2), Myrceugenia oxysepala (Myrc oxy 1 a 4), Cinnamomum glaziovii (Cin glaz 1 a 3), Cryptocarya aschersoniana Mez(Crypt as 2 e 3), Myrceugenia miersiana (Gardner) D. Legrand et Kausel (Myrc mie 2), Prunus myrtifolia (L.) Urb. (Prun myr 2), Rollinia rugulosa Schltdl. (Roll rug 2), Myrcia oligantha O. Berg (Myrc oli 3) e Ocotea indecora (Schott) Mez (Ocot ind 3). A espécie Matayba elaeagnoides é descrita por Reitz (1980), Reitz et al. (1983) e Lorenzi (1998) como mesófila e seletiva higrófila, ocorrendo no local provavelmente pelas boas condições de umidade causadas pelo dossel fechado/entreaberto, pela baixa declividade e também por situar-se na encosta inferior do terreno. O interessante é que duas espécies de Myrtaceae ocorreram nesse grupo como espécies indicadoras, representadas por Myrceugenia myrcioides e Myrceugenia oxysepala como espécies seletivas higrófilas e ciófitas (Legrand e Klein, 1970), desenvolvendo-se no interior das matas, sendo bastante freqüentes somente nos solos bastante úmidos e muito raras nos solos de drenagem rápida das encostas mais íngremes. Essa afirmação, dos autores anteriormente citados, é confirmada ao se analisar a matriz dos fatores ambientais (Narvaes, 2004), em que há boa condição de umidade decorrente da baixa declividade, da baixa pedregosidade, e estarem sob dossel fechado/entreaberto, situadas na encosta inferior do terreno. Sobre essa perspectiva, apesar de não se conhecer descrições ecológicas à respeito de Cinnamomum glaziovii, e pelo fato de que o gênero possui pouco interesse florestal (Marchiori, 1997), ela possui o mesmo padrão ecológico das outras espécies indicadoras do grupo. A espécie indicadora do grupo ecológico 3 (G-3) foi Zanthoxylum petiolare A. St.-Hil. & Tul. (Zant pet 1 e 2), presente em 28 e 14 subparcelas respectivamente, e caracterizada por ser uma espécie ciófita e seletiva xerófita (Cowan e Smith, 1973), ocorrendo preferencialmente nas encostas enxutas ou íngremes das matas primárias, confirmada pela matriz dos fatores ambientais desse grupo ecológico (Narvaes, 2004) no qual a alta declividade do terreno e a posição topográfica de encosta superior/platô, tornam o sítio drenado com baixa umidade. Dado a esse fato, tal grupo é provavelmente o que possui a situação de estágio de menor alteração, possibilitando a ocorrência de espécies adaptadas a ambientes sombreados. Já as espécies preferenciais desse grupo são Dalbergia frutescens (Vell.) Britton (Dalb fru 1), Eugenia involucrata DC. (Eug invol 1), Ilex dumosa Reissek (Ilex dum 1), Mollinedia elegans Tul. (Molli el 1,2 e 3), Myrceugenia cucullata (Myrc cuc 1 e 2), Myrcianthes gigantea (D. Legrand) D. Legrand (Myrc gig 1), Myrrhinium atropurpureum Schott (Myrr atr 1), Nectandra megapotamica (Spreng.) Mez (Nect meg 1), Ocotea pulchella (Ness) Mez (Oct pulc 1 e 2), Podocarpus lambertii Klotzsch ex Endl. (Podoc la 1), Sebastiania brasiliensis (Sebst bra 1), Siphoneugena reitzii D. Legrand (Siph rei 1 e 2), Zanthoxylum petiolare (Zant pet 1 e 2), Myrciaria delicatula (DC.) O. Berg (Myrc del 1), Ocotea diospyrifolia (Meisn.) Mez (Oct dio 1), Duranta vestita Cham. (Duran ve 1), Cryptocarya moschata Nees e Mart. ex Ness (Crypt mo 1), Piptocarpha notata (Less.) Baker (Piptc no 1 e 2), Gordonia fruticosa (Schrad.) H. Keng (Gord fru 1 e 2), Cipós (Cipós 2), Myrsine umbellata Mart. (Myrs umb 3) e Rudgea parquioides (Rudg par 4 e 5). Diferenciação entre grupos florísticos Com base nos grupos florísticos formados, podem ser feitas as seguintes observações, como forma de obter diferenciações entre eles. O grupo ecológico 1 (G-1) foi caracterizado pela presença conjunta das espécies Sebastiania brasiliensis e Casearia decandra, esta última “generalista”, pois ocorre em todos os grupos. Essas espécies, além de Calyptranthes concinnaDC. e Stillingia oppositifolia,são as mais freqüentes, porém não-exclusivas do grupo ecológico. Entretanto, verifica-se que são as mais adaptadas às saturações hídricas, pois ocorreram em maior freqüência do que nos outros dois grupos (G-2 e G-3). O primeiro grupo tem como grande diferencial o elevado número de espécies exclusivas, tais como: Dasyphyllum tomentosum (Spreng.) Cabrera, Hennecartia omphalandra Poisson, Luehea divaricataMart. & Zucc., Machaerium paraguariense Hassl., Ocotea puberula (Rich.) Nees, Pilocarpus pennatifolius Lem., Rollinia salicifolia Schltdl., Sapium glandulosum (L.) Morong, Sloanea monosperma Vell., Solanum iraniense LB Sm. & Downs, Solanum pseudoquina A.St.-Hil., Solanum mauritianum Scop., Styrax leprosus Hook. & Arn., Chomelia obtusa Cham. & Schltdl., Piper gaudichaudianum Kunth, Myrceugenia foveolata (O.Berg) Sobral, Cestrum intermedium Sendtn., Justicia brasiliana Roth, Endlicheria paniculata (Spreng. J. F.) Macbr., Myrcia arborescens O. Berg, Bauhinia microstachya (Raddi) J.F. Macbr., Macfadyena ungüis-cati (L.) A. H. Gentry, Pavonia malvacea (Vell.) Krapov. & Cristóbal, Acalypha gracilis Spreng., Anchietea parvifoliaHallier F. e Cyphomandra corymbiflora Sendtn. Muitas dessas espécies, por não apresentarem formas de vida arbustiva e de árvores (trepadeira, cipó e erva), se encontram nesse grupo classificadas como raras, pois a dimensão mínima para a inclusão de indivíduos é possuir pelo menos 3,0 cm de circunferência e pelo menos 1,30 m de altura para a sua inclusão; por outro lado, cabe destacar que Dasyphyllum tomentosum, Luehea divaricata, Ocotea puberula, Rollinia salicifolia, Sapium glandulosum, Myrceugenia foveolata, Solanum iraniense, Solanum pseudoquina, Solanum mauritianum, Styrax leprosus e Cyphomandra corymbiflora são espécies de hábito arbóreo-arbustivo e que apresentam padrão aleatório de distribuição espacial, exceto Luehea divaricata que, provavelmente, dê indícios de agregação nesse grupo por causa das suas características ambientais, possuindo baixa freqüência e densidade relativamente alta. O grupo ecológico 2 (G-2) teve como aspecto interessante a presença de Cabralea canjerana (Vell.) Mart. e Alsophila sp. como exclusivas, distinguindo-se do grupo ecológico 1, especialmente pela elevada freqüência de Cinnamomum glaziovii. A diferença dos sítios G-2 e G-3, com relação à presença/ausência das espécies pode ser constatada pela presença de Allophylus guaraniticus, Cedrela fissilis Vell., Celtis iguanaea, Acca sellowiana (O. Berg) Burret, Ilex dumosa, Ilex microdonta Reissek, Myrsine coriácea (Sw.) R. Br., Nectandra megapotamica, Scutia buxifolia Reissek, Solanum sanctaecatharinae Dunal, Symplocos uniflora (Pohl) Benth., Trichilia elegans A. Juss., Xylosma tweediana (Clos) Eichler, Zanthoxylum kleinii (R. S. Cowan) P. G. Waterman, Zanthoxylum petiolare, Zanthoxylum rhoifolium Lam, Psychotria leiocarpa Cham. & Schltdl., Citharexylum solanaceum Cham., Myrciaria delicatula, Miconia hyemalis A. St.-Hil & Naudin ex. Naudin, Oreopanax fulvus Marchal, Lippia ramboi Moldenke, Ocotea diospyrifolia, Myrcia lajeana D. Legrand, Duranta vestita, Symphyopappus itatiayensis (Hieron.) R. M. King & H. Rob., Clethra uleana Sleumer, Solanum pabstii L. B. Sn. & Downs, Xylosma ciliatifolia (Clos) Eichler¸ Casearia obliqua Spreng. e Heteropterys sp., no grupo 3 (G-3) e a ausência de Cabralea canjerana, Rhamnus sphaerosperma Sw., Alsophila sp.e Acacia sp., nesse mesmo grupo. Ao se comparar o G-3 e o G-1, observou-se um grande número de espécies presentes em um grupo e ausentes em outro; com a presença de Ilex microdonta, Symplocos uniflora, Xylosma tweediana, Zanthoxylum kleinii, Psychotria leiocarpa, Cinnamomum amoenum (Nees) Kosterm., Myrcia lajeana, Myrsine loefgrenii (Mez) Imkhan., Clethra uleana, Xylosma ciliatifolia, Gordonia fruticosa, Heteropterys sp. e Solanum inodorum Vell., no grupo 3e a ausência no grupo G-1. As espécies exclusivas no grupo 3 foram Ilex microdonta, Symplocos uniflora, Xylosma tweediana, Xylosma ciliatifolia, Zanthoxylum kleinii, Psychotria leiocarpa, Myrcia lajeana, Clethra uleana e Heteropterys sp. Caracterizando a ocorrência de cada grupo, com relação às características gerais o G-1 (encosta média), ocorreu em cotas de transição, em relação àquelas extremas, o que logicamente permite a seletividade de espécies tolerantes a tais características. O fator relevante a ser considerado nesse grupo diz respeito ao nível do lençol freático que, apesar de situar-se na encosta média, o solo desse grupo possui boa condição de umidade, decorrente sobretudo da baixa declividade. Com base na composição florística de cada grupo ecológico, foi possível constatar que a distribuição e a ocorrência de determinadas espécies são diretamente relacionada aos fatores ambientais predominantes em cada local, sendo influenciadas diretamente pela posição topográfica do relevo, influindo também nas outras características ambientais, já que a abertura do dossel (entrada de luz) é muito semelhante em todos os grupos, sendo essa tendência também constatada por Araujo (2002) em um fragmento de Floresta Estacional Decidual Ripária. Conforme Runhaar e Haes (1994 apud ARAUJO, 2002), a composição das espécies está em equilíbrio com as condições do ambiente até que estas mudem e, conseqüentemente, as espécies já não serão indicativas daquele ambiente, tendendo a haver uma substituição. Sobre a perspectiva dos fatores ambientais, é evidente que estes são determinantes na formação dos grupos florísticos, decorrentes, sobretudo da posição topográfica e conseqüente umidade de cada grupo. Um aspecto muito importante, que deve ser mencionado, é que algumas unidades amostrais, apesar de estarem próximas, ficaram classificadas como características de outro grupo, por ocasião da análise de classificação de grupos pelo método TWINSPAN como, por exemplo, as unidades de 2 a 6, que apesar de estarem próximas das unidades 1 e 7, apresentam características ambientais peculiares à encosta inferior do relevo. Para a formação dos grupos, por ocasião da primeira divisão, no grupo 1 do total de 121 subparcelas, 11 subparcelas foram consideradas borderlines, e nenhuma foi mal classificada. Na segunda divisão, o grupo 2 (28 subparcelas) apenas duas foram consideradas mal classificas e uma borderlines, enquanto que, para o grupo 3, do total de 31 subparcelas, nenhuma foi considerada mal classificada ou borderlines. Considerando que as parcelas consideradas mal classificadas podem comprometer a análise fitossociológica, estas foram excluídas por ocasião de sua análise. Florística dos Grupos Ecológicos encontrados na Regeneração Natural da FLONA A amostragem utilizada para representar as espécies dos diferentes grupos florísticos, bem como a fisionomia da vegetação em cada grupo podem ser vistos nos trabalhos de Narvaes (2004) e Narvaes et al. (2005). O grupo ecológico 3 (G-3) se destacou por apresentar maior diversidade florística (Tabela 1), pelo fato de provavelmente reunir as condições favoráveis para o estabelecimento de uma maior gama de espécies, estando situado na encosta superior/platô, em um local com elevada declividade, boas condições de luminosidade e, por conseguinte, de boa drenagem, além de possuir exposição do relevo razoavelmente favorável à entrada de luz (noroeste), conforme Narvaes (2004). Esse conjunto de fatores favoreceu o estabelecimento de um maior número de espécies na área, influenciando especialmente as espécies que necessitam de locais bem drenados e de boa condição de luminosidade. Os grupos florísticos 1, 2 e 3 apresentaram, respectivamente, 106, 62 e 87 espécies; oitenta, 53 e 59 gêneros; e 41, 27 e 32 famílias, além de indivíduos mortos e que por ocasião da medição, se encontravam em pé, indivíduos não-identificados e cipós não-identificados. A diversidade foi superior no grupo ecológico da encosta superior/platô (G-3) do que nos outros dois grupos (encosta inferior e encosta média), evidenciando relativa diferença (Tabela 1), provavelmente em conseqüência do maior número total de indivíduos no G-2 e concomitantemente com o pequeno número de espécies por unidade amostral do G-1, fazendo com que a diversidade diminuísse. Pelo índice de Shannon o G-2, apresentou leve superioridade a respeito da riqueza florística em relação ao G-1. Estes valores foram ligeiramente inferiores aos encontrados por Mauhs e Backes (2002) no mesmo tipo florestal, porém esse resultado foi obtido em um fragmento exposto a perturbações antrópicas e, mesmo assim, a diversidade foi bastante significativa (H’= 2,90) para a regeneração natural. Nesse local, cujo gado acarretava importante influência antrópica, foi observada baixa riqueza florística (59 espécies de hábito arbóreo), assim como ocorreu no G-3. TABELA 1: Número de famílias (F), gêneros (G), espécies (Sp), índice de diversidade de Shannon (H’) dos três grupos florísticos da Regeneração Natural da Floresta Ombrófila Mista na FLONA de São Francisco de Paula, RS, Brasil. TABLE 1: Family number (F), genera (G), species (Sp), Shannon diversity index (H’) of three floristic groups in the Mixed Rainy Forest at FLONA of São Francisco de Paula, RS, Brazil.
Já Jarenkow (1985) verificou, na estação ecológica de Aracuri, no mesmo tipo florestal, índice de Shannon igual a 2,93, porém, no estrato considerado por ele como arbustivo (Dap ³ 5,0cm). Estudos da regeneração natural, os quais levam em conta a comparação da diversidade florística, devem ser postos à prova com o de outros tipos fitogeográficos, sobretudo quando analisados com os tipos que fazem transição com a mata de araucária, como é o caso do estudo realizado por Dorneles e Negrelle (2000) o qual se encontra na zona de ecótono com a Floresta Atlântica (Floresta Ombrófila Densa), verificaram índice de diversidade de Shannon médio de 3,00 para os estágios de crescimento adulto e juvenil. Entretanto, em um estudo realizado por Caldato et al. (1996) na Floresta Ombrófila Mista, em uma floresta primária, foi encontrado índice de Shannon de 1,68, valor bem inferior ao encontrado para todos os grupos florísticos deste estudo (Tabela 1). Isso pode ser explicado pelo seu estágio de sucessão natural, já que o número de espécies secundárias tardias e clímax são mais restritas do que espécies secundárias iniciais e pioneiras, estas tolerantes a diferentes graus de perturbação da floresta. No G-1, predominam as famílias Myrtaceae (vinte espécies), Solanaceae (dez), Lauraceae (nove), Euphorbiaceae (cinco) e Asteraceae (cinco), caracterizando o grupo de maior representatividade florística, ao contrário do G-2, que apresentou Myrtaceae (16), Lauraceae (seis) e Solanaceae (quatro). No grupo ecológico 3, ocorreram 19 espécies de Myrtaceae, oito de Lauraceae, cinco de Salicaceae e Solanaceae. Quando se faz afirmações a respeito da riqueza florística, deve-se tomar muito cuidado, já que nem sempre as famílias que possuem o maior número de espécies são as mais bem representadas. No G-1, apesar da Myrtaceae possuir um número mais representativo de espécies, a família Euphorbiaceae apresentou praticamente o dobro da densidade, com 3.005,26 indivíduos/ha na regeneração natural, sendo que a maior parte representada pelas espécies Stillingia oppositifolia (1.667,54 ind/ha) e Sebastiania brasiliensis (1.289,47 ind/ha), enquanto que Myrtaceae possuía 1.516,64 indivíduos/ha. No G-2, apesar de Myrtaceae (1.886,21 ind/ha) possuir o número mais representativo de espécies, a família que apresentou maior densidade foi Salicaceae (2.589,66 ind/ha), representados apenas por duas espécies (Casearia decandra e Xylosma pseudosalzmannii), seguida de Euphorbiaceae com densidade de 2.172,41 ind/ha, representada por três espécies, sendo que Stillingia oppositifolia perfez quase que a totalidade da densidade, com 2.000 ind/ha. Um aspecto importante do grupo ecológico 2 é que a espécie Casearia decandra, por ocorrer em grande abundância e freqüência no grupo, se constituiu em uma espécie generalista, adaptando-se a muitos habitats, sendo descrita por Marchiori (1997) como árvore pequena que habita, sobretudo, o estrato médio da Floresta Ombrófila Mista, confirmando os resultados encontrados. No G-3, ocorreu a maior densidade de espécies da família Rubiaceae (1.443,24 ind/ha), seguido de perto por Salicaceae (1.432,44 ind/ha), enquanto que Myrtaceae apresentou 1.075,68 indivíduos/ha. Em um fragmento de Floresta Ombrófila Mista, Rondon Neto et al. (2002) observaram que, apesar da família Myrtaceae possuir o maior número de espécies, ou seja, a maior riqueza, os seus indivíduos se apresentaram com baixa densidade, sendo que a maior densidade de indivíduos foi das famílias Sapindaceae, Salicaceae e Lauraceae. Calegari (1999) observou Myrtaceae, em primeiro lugar, com o maior número de indivíduos, porém Euphorbiaceae foi a família que obteve a maior densidade. Caldato (1998)também verificou que a família Myrtaceae apresentou o maior número de espécies. Mauhs e Bakes (2002), estudando a regeneração natural, observaram Myrtaceae, em primeiro lugar, e com um número elevado de indivíduos (35,44%). Com base no que foi relatado na descrição dos grupos, o que pode ser dito a respeito da família Myrtaceae é que ocorrem comumente na regeneração natural da Floresta Ombrófila Mista bem como na Floresta Atlântica, conforme Dorneles e Negrelle (1999; 2000), apresentando tolerância a ambientes sombreados e úmidos nos quais são bastante representativas quanto à densidade de indivíduos. A síndrome de dispersão zoocórica é o mecanismo de dispersão das espécies que compõem a família Myrtaceae o que serve como atrativo de dispersores aos diferentes grupos florísticos, aumentando a possibilidade de regeneração natural desses ambientes (Araujo, 2002). Como fato interessante, cabe ressaltar que, em todos os grupos florísticos, os gêneros predominantes foram Myrceugenia (guamirim) e Eugenia (Pitanga, cerejeira-do-mato e batinga). CONCLUSÕES Com base nos resultados, conclui-se que: a) Existem três grupos florísticos avaliados nas unidades amostrais, associadas às mudanças na topografia. Desse modo, a regeneração natural dessas áreas não pode ser tratada como um todo, mas se levando em conta tais variações; b) A diferença entre a diversidade florística dos grupos formados demonstrou a evidente separação dos grupos florísticos analisados, cada um com suas peculiaridades florísticas, decorrentes das diferenças na posição topográfica; c) Sebastiania brasiliensis e Stillingia oppositifolia estão adaptadas à posição topográfica intermediária (encosta média) onde existem boas condições de umidade, acarretada em especial pela baixa declividade; Casearia decandra, Matayba elaeagnoides, Myrceugenia oxysepala e Myrcia oligantha são espécies tolerantes às condições ambientais extremas de umidade e até a alagamentos eventuais (encosta inferior), juntamente com Stillingia oppositifolia, esta com características de maior ‘plasticidade’ com relação à disponibilidade hídrica do solo; e Casearia decandra, Rudgea parquioides e Myrsine umbellata foram tolerantes a locais situados na encosta superior/platô e de alta declividade que, por conseguinte, não sofrem a influência da elevação do teor de umidade. Essas oito espécies estiveram bem representadas na regeneração natural da floresta e são fundamentais na conservação dos grupos florísticos e, conseqüentemente, da floresta estudada. REFERÊNCIAS BIBLIOGRÁFICAS
Copyright 2008 - Ciência Florestal The following images related to this document are available:Photo images[cf08021f1.jpg] [cf08021f2.jpg] | ||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}