 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
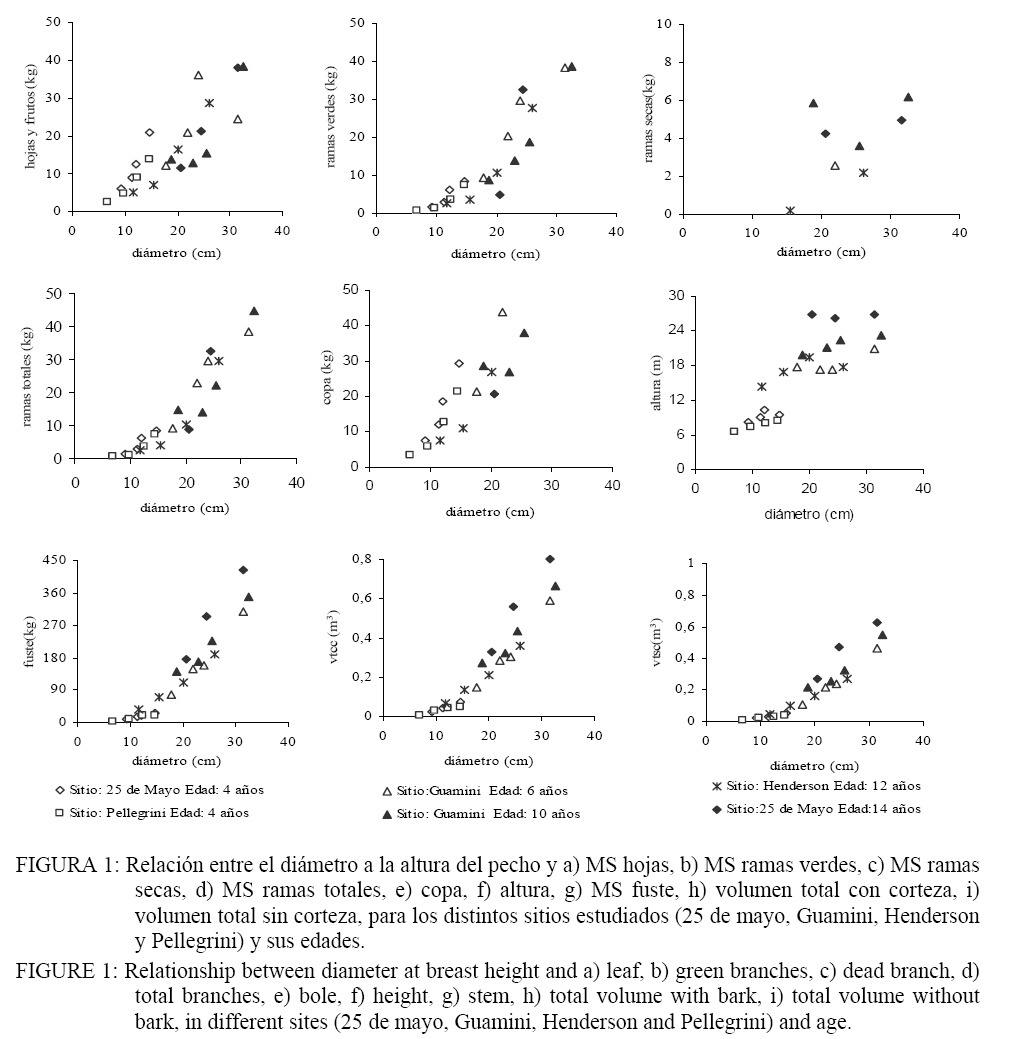
Ciência Florestal, Vol. 18, No. 3, 2008, pp. 291-305 BIOMASA EN PLANTACIONES DE Eucalyptus viminalis Labill. DE LA PROVINCIA DE BUENOS AIRES, ARGENTINA BIOMASS IN Eucalyptus viminalis Labill. PLANTATIONS IN BUENOS AIRES PROVINCE, ARGENTINA Paula Ferrere1, Ana Maria Lupi2, Rosa Boca3, Vicente Nakama4, Adelqui Alfieri5 1Ingeniera Forestal, Inst. Rec. Biól. AER INTA 9 de Julio, Mitre 857 (6500) 9 de Julio, Buenos Aires, Argentina. Recebido para publicação em 8/03/2007 e aceito em 11/08/2008. Code Number: cf08027 RESUMEN El presente trabajo se desarrolló en el oeste de la provincia de Buenos Aires, Argentina con el objetivo de ajustar funciones de biomasa de árboles individuales y determinar la biomasa en diferentes compartimientos de la planta y el sotobosque. Se identificaron rodales de Eucalyptus viminalis Labill. cuyas edades oscilaron entre 4 y 14 años. Se apearon 21 árboles con diámetros entre 9,2 y 32,5 cm. Se desarrollaron regresiones simples y múltiples y se estimó el volumen, la biomasa en ramas, hojas y fuste. Las ecuaciones de volumen de mejor comportamiento son basadas en modelos lineales en su forma normal, y el modelo más adecuado fue el que incorporó al d2 (R2 = 0,92). Para la estimación de la biomasa en hojas, ramas, copa y fuste se recomienda los modelos ln-ln que consideran el d y h o solo el d. El componente de la biomasa en hojas presentó una estimación más débil. La distribución de la biomasa en los individuos sigue el patrón de desarrollo que se encuentra en la bibliografía. La proporción de la biomasa en la copa disminuye con la edad, contrariamente a lo sucedido con el fuste. Palabras claves: modelos de biomasa; modelos de volumen; Eucalyptus; región semiárida.ABSTRACT The present work was developed in the West of Buenos Aires Province (Argentina) with the objective of adjusting functions of biomass of individual trees, in their different compartments and in the understorey. Stands of Eucalyptus viminalis Labill. were identified, with ages between 4 and 14 years-old. Twenty-one individuals were felled with diameters ranging from 9,2 to 32,5 cm. Simple and multiple regression models were developed and volume, branch leaf and stem biomass were estimated. The best volume equations were based on lineal models and the most adequate behavior was obtained with d2. To estimate leaf, branch and stem, ln-ln models have been suggested, with diameter and h or only diameter. The leaf biomass presented the weakest adjustment. The distribution of trees biomass agrees with the bibliography. The proportion of crown biomass decreases with age; on the other hand, the proportion of stem biomass increases with age. Keywords: biomassmodels; volume models; Eucalyptus; semiarid region.INTRODUCCIÓN En sentido amplio la estimación de la biomasa en plantaciones forestales es una información utilizada para comprender el funcionamiento y los procesos del ecosistema. En términos particulares es una información de base para el cálculo de la producción de un sitio y para el estudio del almacenaje de los nutrientes en el rodal. Sin embargo, en los últimos años la cuantificación de la biomasa ha tomado relevancia con el fin de evaluar la potencialidad que tienen los bosques como destino del CO2 atmosférico y cuantificar así su capacidad de almacenaje. La biomasa en plantaciones se estima mediante la selección y extracción de un determinado número de individuos para luego establecer su materia seca. Sin embargo, la metodología de cuantificación de la biomasa en forma directa es muy engorrosa y habitualmente muy costosa que solo puede ser enfrentada mediante proyectos de investigación básica y específicos. Para salvar esta limitante se han desarrollado relaciones alométricas que permiten estimar la biomasa de los distintos componentes del árbol mediante ecuaciones donde las variables observadas son de fácil registro y el número de árboles extraídos es limitado (PORTÉ et al., 2002). Aplicando estas relaciones a datos de inventario pueden obtenerse estimaciones indirectas de la biomasa del rodal para un sitio específico con gran precisión (BELLOTE e SILVA, 2004). Montagu et al. (2005) trabajando con Eucalyptus pilularis Sm, desarrollaron una relación alométrica regional para estimar la biomasa de diferentes componentes del árbol en distintos ambientes. Según los autores, utilizando el diámetro a la altura del pecho como única variable predictora se logró la relación más estable, si se lo compara con la adición de la altura. Wang (2006) también indica que el uso de diámetro como variable independiente explica una gran parte de la biomasa leñosa; sin embargo, la inclusión de la variable altura mejora las estimaciones de algunos componentes como las hojas. Navar et al. (2001) desarrollaron modelos de regresión lineal simple y múltiple, y destacan que los primeros ajustaron mejor la biomasa total a partir de sus componentes. Laclau et al. (2000) establecieron modelos predictivos donde el diámetro a la altura del pecho y la edad fueron las variables independientes utilizadas para estimar la biomasa y el contenido de diferentes nutrientes. En Argentina, los estudios de biomasa son relativamente recientes concentrándose la mayor parte en la última década. Los primeros fueron desarrollados en plantaciones de Eucalyptus grandis Hill sobre diferentes tipos de suelos de la provincia de Entre Ríos (GOYA et al., 1997). Estos, además de estimar la biomasa desarrollan modelos alométricos y cuantifican los nutrientes almacenados en diferentes compartimientos de la biomasa aérea. Un trabajo similar se desarrolló en plantaciones de Pinus taeda L., Pinus elliottii Engelm. y Araucaria angustifolia (Bert.) Kuntze. en la provincia de Misiones (GOYA et al., 2003; FERNANDEZ TSCHIEDER et al., 2004), donde los parámetros utilizados en las ecuaciones fueron el diámetro, para estimar el peso seco en hojas y ramas, y el diámetro cuadrático y la altura para el peso seco del fuste. Por otro lado, en la Patagônia, Laclau (2003) empleó el diámetro y el volumen como variables independientes para un estudio de biomasa en diferentes compartimientos de Pinus ponderosa Douglas ex Lawson & C. Lawsony Austrocedrus chilensis (D. Don) Florin & Bontelje. La región oeste de la Provincia de Buenos Aires, conocida como Pampa Arenosa, presenta una alta predominancia de la actividad agrícola-ganadera. A pesar de su gran potencialidad, el sector forestal cuenta con escaso desarrollo que puede ser atribuido a cuestiones culturales y financieras, inclusive en sectores con presencia de suelos que no compiten con la actividad agrícola. La región de la pampa arenosa es ecológicamente apta para el cultivo de Eucalyptus viminalis (NAKAMA et al., 2000; MOSCHINI et al., 2000), sin embargo, solo se observan pequeños montes de reparo menores a 10 has y cortinas. En la actualidad, la posibilidad de contar con proyectos forestales destinados a brindar servicios ambientales ha puesto en relieve la ausencia de información básica forestal que permita ser empleada en la formulación de estos emprendimientos. En este trabajo se reportan los resultados de un estudio desarrollado en el oeste de la provincia de Buenos Aires (Argentina) sobre plantaciones de Eucalyptus viminalis. Los objetivos fueron: (i) ajustar funciones de biomasa de árboles individuales, (ii) determinar la biomasa aérea en diferentes compartimientos de la planta y en el sotobosque. MATERIAL Y MÉTODOS Área de relevamiento de datos Se trabajó con rodales de Eucalyptus viminalis del centro-oeste hasta el sur oeste de la provincia de Buenos Aires, Argentina. Esta subregión denominada geomorfológicamente “Pampa Arenosa” se compone principalmente por lomadas arenosas intercaladas con bajos que se orientan de sudoeste a noreste. Los suelos se desarrollaron sobre sedimentos loésicos transportados por el viento, de textura arenosa, las que fueron depositadas en sucesivas capas de gran espesor. Sobre los suelos más desarrollados del norte de la región, la vegetación natural es la pastura que fue reemplazada por cultivos anuales. Hacia el sudoeste, algunos médanos todavía se encuentran con escasa cobertura natural y en muchos casos se ha sembrado pasto llorón Eragrostis curvula (Schrad) para su estabilización. La temperatura media anual oscila entre 15 °C y 16 °C; sólo los partidos Daireaux y Guaminí, hacia el sur, observan temperaturas medias anuales inferiores a los 15 °C. Las precipitaciones medias anuales varían de los 1000 mm en el NE a 800 mm en el extremo SO del área de trabajo. En el mismo sentido, el número medio de días con heladas varía de 20 a 45 (MOSCHINI et al., 2005). Según la clasificación de Köppen-Grieger, el clima corresponde al grupo Cfa, es decir templado y húmedo, sin estación seca, con la temperatura media del mes más cálido mayor a 22ºC. Se trabajó con mapas digitalizados de suelos de la Región Pampeana a escala 1:50.000 (INTA, 1990) y se identificaron unidades que corresponden al Subgrupo de los Udipsamentes típicos y Hapludoles énticos (USDA, 2003). En la Tabla 1 se describen las características de las unidades taxonómicas de suelos donde se realizó el relevamiento. Si bien estos suelos ocupan una menor superficie en el total de la región, adquieren relevancia a los fines de la estimación de la biomasa para futuros inventarios de carbono debido a su buena aptitud forestal sin competir con la actividad agrícola-ganadera. TABLA 1: Principales características de las unidades taxonómicas en los sitios de muestreo. TABLE 1: Main characteristics of the taxonomy units in places of sampling.
Donde:P = posición; R = relieve; PE = pendiente; D = drenaje; E = escurrimiento; Per = permeabilidad. Mediciones forestales Se localizaron plantaciones de Eucalyptus viminalis Lab.de diferentes edades (Tabla 2) con densidad inicial homogénea (1111 pl.ha-1) y sin intervenciones silvícolas. La edad de las plantaciones fue obtenida del registro que poseen los productores. Los rodales muestreados cuentan con una superficie entre 1-3 has. En cada uno de ellos se instalaron, al azar, entre 2 y 4 parcelas circulares de 400 m2 y se realizó un inventario midiendo el diámetro a la altura del pecho (d) de todos los árboles y la altura (h) de 3 individuos: correspondiente al diámetro mayor, menor y promedio. En cada rodal, por fuera de las parcelas de inventario, se apearon entre 2 y 4 individuos promedio, asumiendo que estos representan adecuadamente a la plantación. Se muestrearon en total 21 individuos cuyos diámetros estaban comprendidos entre los 9,2-32,5 cm. Cada uno fue separado en los siguientes compartimientos: a) fuste hasta 5 cm de diámetro (fu); b) ramas verdes (rv); d) ramas secas (rs) y e) hojas y frutos (ho). A los fines del estudio no se separó esta última fracción porque los frutos no representan una proporción importante. El peso verde del fuste en las plantaciones de 4 años y las ramas, hojas y frutos de todos los individuos fue pesado in situ. Una muestra de cada componente fue llevada a estufa hasta peso constante (65-70ºC) para determinar la humedad y calcular la materia seca (MS). Para los individuos mayores a 4 años la determinación de la MS se realizó a través de la cubicación de la siguiente manera: Se midió la altura total del árbol (m), el diámetro en la base (cm) o altura del corte (entre los 10-15 cm de altura), a 1,30 m y cada 2 m hasta los 5 cm de diámetro de punta fina. En cada troza se midió el diámetro en ambos extremos en dos sentidos opuestos. Se calculó el volumen total con corteza (vtcc) y sin corteza (vtsc). Una vez calculado el volumen del fuste se lo multiplicó por el valor de densidad básica de la madera de Eucalyptus viminalis (δ: 505 kg. m-3) determinado según Normas IRAM por Repetti (1990). TABLA 2: Localización de los sitios de muestreo, tipo de suelos, edad de las plantaciones, número de parcelas, diámetro cuadrático promedio (dg) y altura promedio (ht). TABLE 2: Site location of sampling, tipe of soils, age of stand and number of plots.
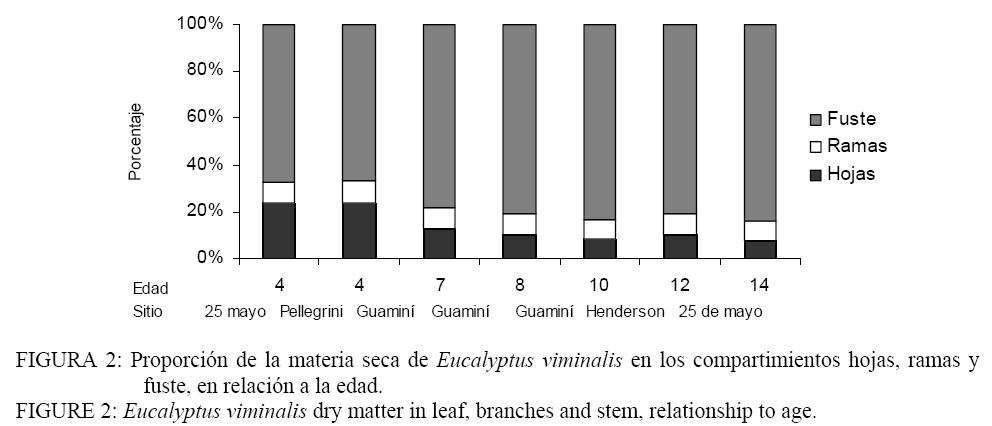
Mediciones de mantillo En cada parcela se tomaron 6 muestras de material vegetal, al azar, en una superficie cuadrada de 0,25 m2. El material se separó en a) mantillo (residuos de hojas, ramas, frutos, corteza de Eucalyptus viminalis Lab. y b) vegetación herbácea (pastos). Se separaron submuestras y se llevaron a estufa (65-70ºC, 72 hs), hasta peso constante. Los valores de MS por parcela fueron expandidos a hectárea. Análisis estadístico Se realizó un análisis exploratorio gráfico para detectar las relaciones existentes entre las variables observadas. Las variables explicatorias fueron: d y h, mientras quelas variables dependientes fueron volumen total con corteza (vtcc); volumen total sin corteza (vtsc), hojas (ho), ramas verdes (rv), copa(co) y fuste (fu). Para cada variable dependiente se ajustaron y testearon cinco modelos lineales: Modelo 1 : yij = β0 + β1d + β2 h + εij Modelo 2: yij = β0 + β1d + εij Modelo 3: ln(yij ) = β0 + β1 ln(d) + β2 ln(h) + εij Modelo 4: ln(yij ) = β0 + β1 ln(d) + εij Modelo 5: yij = β0 + β1d2 + εij Donde yij= variable dependiente; β0, β1y β2 son los parámetros del modelo y εij es el error aleatorio. Se asumió que los errores eran independiente e idénticamente distribuidos, con distribución Normal (0;σ ). Para la estimación de los parámetros de los modelos se utilizó el método de mínimos cuadrados (WEBER y SKILLINGS, 2000). Los cálculos estadísticos fueron realizados con R (DEVELOPMENT CORE TEAM R, 2003). Para cada modelo se realizó un diagnóstico de influencia de datos (FREUND y LITTELL, 1991) para detectar puntos influyentes y determinar su naturaleza, se utilizaron estadísticos de influencia que indican como se modifican los parámetros del modelo al substraer la i-ésima observación. Los criterios de evaluación estadística para analizar los modelos fueron: a) la equilibrada distribución de la suma de los cuadrados de los residuos, b) los altos R2 ajustados, c) el menor error promedio (EAP) que se obtiene como la media de la diferencia entre los valores absolutos medidos y estimados y d) la simplicidad de los modelos, o sea, que incluyan el menor número de variables regresoras. RESULTADOS Y DISCUSION Biomasa y volumen de árboles En la Figura 1 se muestra la relación entre la variable independiente diámetro (d) y las variables dependientes: MS en fuste, ramas, hojas y frutos, copa, altura (h) y volumen (vtcc y vtsc). El d es un parámetro que explica satisfactoriamente la variabilidad de la biomasa en diferentes compartimentos del árbol, como así también, las variables dasométricas que muestran el estado del rodal (ZIANIS y MENCUCCINI, 2003; SAINT-ANDRÈ et al., 2005 ; COLE y EWEL, 2006). Las asociaciones analizadas fueron buenas, excepto para la MS en ramas secas. En el compartimiento hojas y frutos se observó la mayor dispersión de los datos en relación a otras partes del árbol. Este comportamiento puede ser atribuido a la edad o a que algunos sitios muestreados presentaron variaciones en la estructura del rodal a causa de la densidad de la plantación y de los procesos de selección natural que se producen por la competencia (SAINT-ANDRÈ et al., 2005; PEICHL y ARAIN, 2007). La estrecha relación encontrada entre la biomasa en ramas verdes y el d no fue afectada cuando en los modelos se incluyeron las ramas secas. En los sitios más jóvenes la biomasa aérea de los individuos fue de 34,8 kg.árbol-1 (LC) y 24,6 kg.árbol-1 (PE). Las diferencias (30% aprox. inferior en PE) pueden atribuirse a mejores condiciones edáficas del sitio LC, localizado al norte de la región. Estos corresponden a la unidad taxonómica de los Hapludoles enticos (Tabla 1) son suelos más desarrollados y con un mayor nivel de fertilidad, comparado con la plantación del sitio PE que se encuentra sobre un Udispsamente tipico. La biomasa en hojas fue el componente que mayores diferencias mostró entre estos dos sitios y alcanzó al 37,7%; lo que significó en valores absolutos una cantidad de 12,2 kg de hojas.árbol-1 en LC y 7,6 kg de hojas.árbol-1 en PE. Los resultados podrían sugerir que a esta edad las condiciones de fertilidad afectarían en mayor proporción el desarrollo del componente fotosintético. En el fuste la variación fue del 22,4%, con valores de biomasa de 17,8 kg.árbol-1 y 13,8 kg.árbol-1 para LC y PE respectivamente. En el sitio AR, de 12 años, la biomasa total fue de 128,5 kg.árbol-1, de los cuales 11,7 kg.árbol-1 corresponden a las ramas, 14,4 kg.árbol-1 corresponden a hojas y 102,4 kg.árbol-1 se localizaron en el fuste. La cantidad de hojas fue similar a la estimada en el sitio de menor fertilidad de 4 años (PE). La biomasa aérea fue la tercera parte de la estimada en los árboles de 14 años. En este sitio, además de presentar condiciones de suelo limitante por fertilidad (Udispsamente tipico), el rodal era habitualmente empleado como refugio de ganado, se encontró particularmente denso y con individuos con escasa rectitud de fuste si se lo compara con las restantes situaciones muestradas. En el sitio de mayor edad, (LC: 14 años) la biomasa aérea fue de 367,9 kg.árbol-1, siendo la biomasa en hojas 23,8 kg.árbol-1 y en fuste 296,9 kg.árbol-1. En este trabajo se observó que el cambio en la partición de la biomasa en los componentes del árbol estuvo relacionado con la edad (Figura 2). Este cambio se caracterizó por un importante aumento en la proporción de la biomasa total concentrada como madera en el fuste, que representó el 55% en los árboles más jóvenes y el 83% en los de mayor edad. La proporción de MS en las hojas varió del 33 % en los árboles de 4 años al 6 % en los de 14 años, en tanto que el porcentaje de ramas se mantuvo aproximadamente constante variando solo desde 12% al 9 % en el mismo período. Similares tendencias fueron reportadas por Montagu et al. (2005) en Eucalyptus pilularis, donde la proporción de la biomasa en el fuste aumentó con edad del 40 al 95 % y la biomasa en hojas disminuyó del 25 a menos del 5 %. Por el contrario, inversamente a lo encontrado en el presente trabajo, la partición de la biomasa en ramas disminuyó del 45 al 5%. De manera similar y trabajando con híbridos de Eucalyptus con edades entre 1 y 6 años, Saint Andre et al., (2005) encontraron que la proporción de la biomasa en hojas disminuyó del 36 al 10% y la proporción de biomasa en el fuste aumentó del 19% al 62%. Laclau et al. (2000), trabajando en plantaciones clonales de Eucalyptus en África con densidades iniciales de 666 pl.ha-1 muestran que a los 4 años el 80% de la biomasa aérea se localiza en el fuste. Este valor es notablemente superior al determinado en este estudio y puede ser explicado por el manejo intensivo aplicado (empleo de clones seleccionados con altas tasas de crecimiento, fertilización inicial y control de malezas). Comparado con otras especies, como Pinus radiata, Beets y Pollock (1987) reportaron tendencias similares a las de nuestro estudios, en plantaciones con edades comprendidas entre 2 y 12 años. La biomasa en hojas decreció del 40% a menos del 20%, el fuste incrementó del 35 al 60% y las ramas permanecieron constantes en el orden del 23%. Peichl y Arain (2007) analizando la partición de la biomasa en fuste en Pinus strobus L. encontraron que ésta aumentó del 25% a los 2 años hasta el 69% en individuos de 65 años. En cuanto a la partición de la biomasa en hojas y ramas vivas observaron un decrecimiento que fue del 34% hasta el 1,5% (hojas) y del 17% al 7 % (ramas vivas). El espesor de corteza promedio fue de 1,22 cm, significando el 22,5% del volumen del árbol. El espesor de corteza varió con la edad desde un mínimo de 0,525 cm a los 4 años (27,2%), hasta un máximo de 2,35 cm a los 14 años (20,8%). Se encontró una estrecha y significativa relación entre este estimador y el d (R2: 0,78; p: 0,01), en coincidencia con lo determinado para la misma especie por Schönau y Boden, (1982). Modelos de volumen y biomasa Los modelos matemáticos empleados para el ajuste de las funciones de volumen (Tabla 3) y biomasa (Tabla 4) son los de empleo frecuente en el ámbito científico y de ecólogos que estudian la dinámica de la biomasa en árboles. Las variables incorporadas (diámetro a la altura del pecho y/o la altura) son las más simples y son relevadas normalmente en inventarios forestales. El empleo de la forma ln o ln-ln, de aplicación corriente en este tipo de estudios expresa una proporcionalidad de los incrementos relativos entre las partes del árbol y además, es ampliamente consistente para las distintas formas de crecimiento. TABLA 3. Ecuaciones de volumen con y sin corteza para Eucalyptus viminalis Lab. TABLE 3. Volume equations with and without bark to Eucalyptus viminalis Lab.
Donde
d = diámetro; h = altura; vtcc
= volumen total con corteza (m3); vtsc = volumen total sin corteza
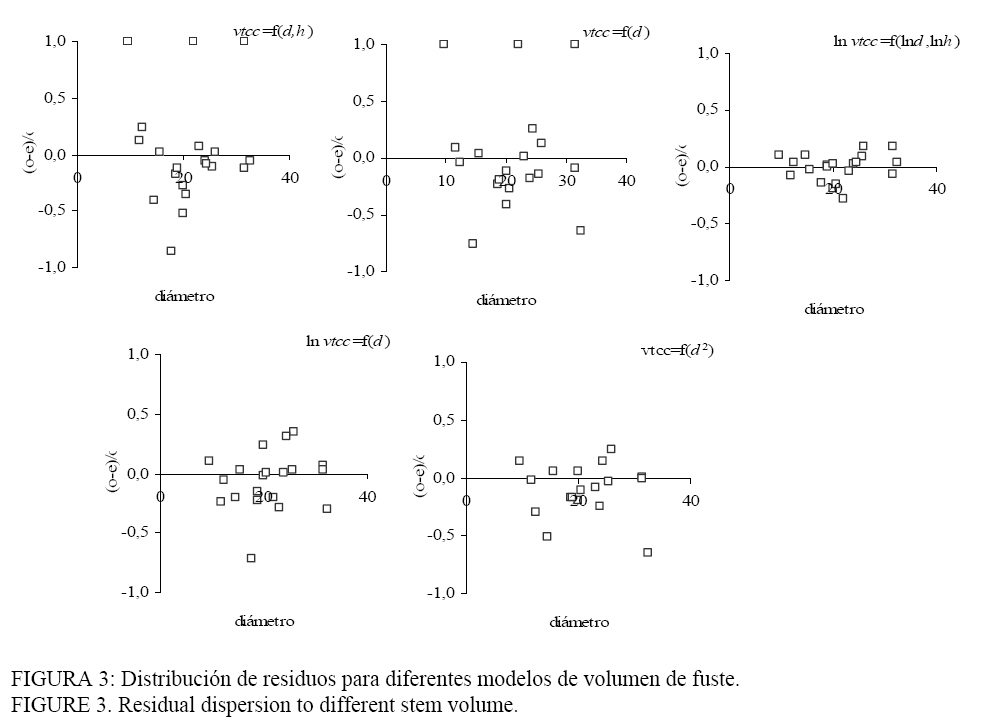
(m3); Todos los modelos de volumen (Tabla 3) presentaron coeficientes significativamente distintos de cero, con p < 0,001, elevados R2 y bajos errores absolutos promedio. En el caso del vtcc los R2 fueron superiores a 0,90, mientras que para el vtsc fueron superiores a 0,88. Los modelos con mayores coeficientes de determinación, bajos
errores absolutos y mejor distribución de residuos (Figura 3) se
correspondieron con las formas ln-ln (modelos 3 y 4), aunque estos
presentan los más altos El modelo (5), que incorpora el diámetro medio cuadrático resulta
ser el más consistente y robusto. Además de evidenciar alto R2, un
bajo Los modelos de volumen sin corteza presentaron un comportamiento
similar al volumen con corteza. Se encontró una mejora en la distribución de
los residuos en los modelos (1) y (2) con los más bajos R2 comparados con los restantes del grupo. Nuevamente las expresiones logarítmicas
(modelos 3 y 4) mostraron los mayores R2 y los residuos más
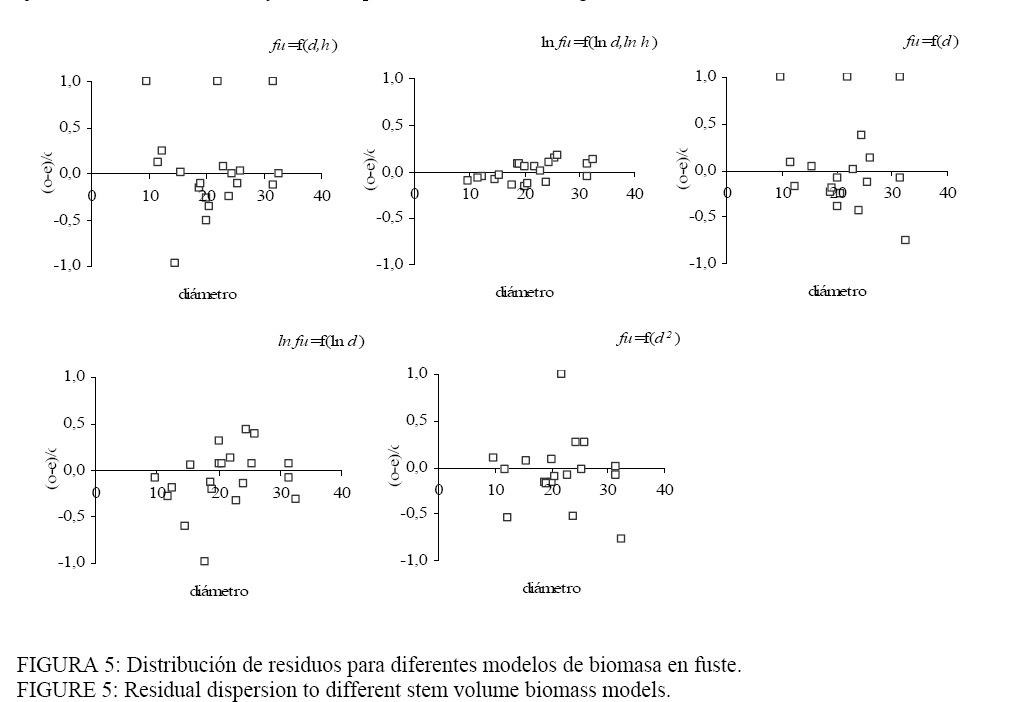
homogéneos, aunque presentaron los valores más altos del Se ha observado que existe una gran concordancia en la bibliografía en cuanto al uso de d como variable regresora. Por otro lado, contrariamente a nuestro trabajo, otros estudios muestran que las trasformaciones de las variables dependientes y/o independientes pueden resultar más adecuadas. Abbot et al. (1997) construyó modelos de estimación de volumen para árboles individuales de diferentes especies. La trasformación de las variables dependiente (volumen) e independiente (d) aplicada por los autores corresponde al log10, que presentó una mejor performance y fue la recomendada cuando se la comparó con la trasformación de raíz cuadrada del volumen y del d en su forma normal. Los coeficientes de determinación de las funciones de volumen para cada especie fueron superiores a 88%. Segura y Kanninen (2005) trabajando con diferentes especies del trópico presentan modelos de volumen del fuste en función del d con un desempeño menor en relación a los obtenidos en este trabajo. En este caso los coeficientes de determinación oscilaron entre 0,63-0,66 para las distintas formas testeadas. Para los autores, el modelo más adecuado (mayor R2, menor CV y menor CME) es el que incluye las variables d y h en su forma logarítmica. Salis et al. (2006) al ajustar funciones alométricas de volumen en especies del Pantanal de Brasil concluyen que la forma log-log es el método más simple y más seguro para estimar el volumen a partir del d como variable independiente. Los modelos de estimación de biomasa aérea (Tabla 4) muestran coeficientes de determinación medios a altos. Las funciones más débiles, relacionadas con los más altos errores absolutos promedios, resultaron ser las de biomasa en fuste y en la copa. TABLA 4: Ecuaciones de biomasa en hojas, ramas, copa y fuste para Eucalyptus viminalis en distintos compartimientos. TABLE 4: Biomass equations for leaf, branches, bole and stem of Eucalyptus viminalis in differents compartments.
Donde: d
= diámetro; h = altura; ho = materia seca de hojas (kg); RV = materia
seca de ramas verdes (kg); Co = materia seca en la copa (hojas + ramas verdes
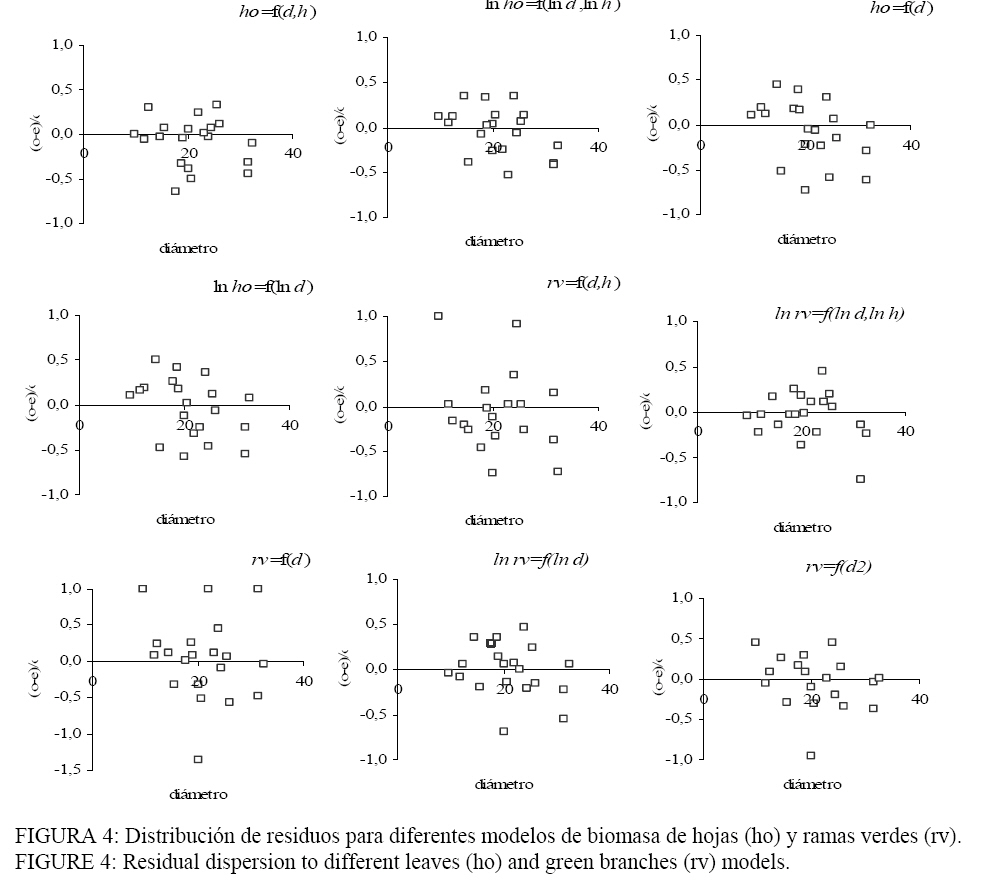
en kg); Fu = fuste materia seca de tronco (kg); De todos los componentes ajustados, la biomasa en hojas fue la fracción con coeficientes de determinación menos satisfactorios (R2: 0,66-0,84), pero aceptables considerando la cantidad de individuos muestrados. Los modelos predictivos de la biomasa en ramas presentaron mejores ajustes que las hojas (R2: 0,82-0,91) mientras que los mayores coeficientes de determinación se encontraron con los modelos de fuste (R2: 0,90-0,99). En los modelos de biomasa de hojas, las expresiones logarítmicas de la variable dependiente e independiente d y h (modelo 2) arrojó lo más altos R2 y menor EAP. Si se elimina el ln h (modelo 4) la precisión del modelo disminuye, pero se logra una ecuación más simple y económica. A pesar de que este componente resultó ser el más variable (WANG, 2006), se observó una distribución homogénea de los residuos de todos los modelos probados (Figura 4) indicando que la dispersión de los datos se mantiene independientemente del tamaño del individuo. En estos casos, los residuos no serían un criterio de se selección y deberían optarse por analizar los restantes parámetros de decisión. Nuestro trabajo concuerda con lo informado por Wang, (2006) y Peichl y Arain (2007) quienes indican que la estimación de la biomasa en hojas es más dificultosa de lograr con sólo el d como variable independiente. Los coeficientes de determinación mencionados en los estudios presentan variaciones similares a los de este trabajo. Veiga et al. (1998) y Salis et al. (2006) en Brasil también destacan que los modelos de biomasa en hojas fueron los de menor ajuste, y destacan las formas doble logarítmicas como las mas apropiadas. Las funciones para la estimación de la biomasa en ramas resultaron
más precisas y con EAP similares. De manera análoga a lo observado en el
componente hojas, los modelos ln-ln fueron los más adecuados, con una
distribución de residuos más uniforme y los mas bajos Para la estimación de la biomasa en la copa, como síntesis de los anteriores compartimentos, los modelos (2) y (4) serian los más robustos y consistentes. El fuste es uno de los componentes de mayor interés por que a edades adultas concentra la mayor parte de la biomasa. En la mayor parte de los sitios, la estimación de la biomasa del fuste se realizó de forma indirecta mediante el empleo de la densidad de la madera y del volumen obtenido por cubicación. Todas las expresiones ajustadas mostraron R2 muy satisfactorios pero se observaron discrepancias en los EAP de los modelos, en la distribución de los residuos (Figura 5) y en los cuadrados medios del error. Los modelos (1) y (3) muestran una distribución de residuos no homogénea, que mejora levemente con el modelo (5). Estos tres modelos presentan los mayores cuadrado medio del error. Las expresiones logarítmicas salvaron estos inconvenientes mostrando una caída sustancial en el cuadrado medio del error y una mejora en la distribución de los residuos. Comparando los modelos (2) y (4) se observa la mayor precisión lograda con el empleo en la función (2) con dos variables independientes; los residuos muestran una distribución mas ajustada, con un elevado R2 y un EAP que solo alcanza los 10 kg. Fernandez Tschieder et al. (2004) también reportaron los mejores ajustes en los modelos de fuste al trabajar con plantaciones en la región subtropical de nuestro país. En este caso los autores también incorporaron el d, la altura o ambas variables. Merino et al. (2003) trabajando con Pinus radiata en la región de Galicia, España encontraron similar comportamiento. Por razones prácticas se eligió el modelo que solo incluye el d, como lo presentado por Verwijst y telenius, (1999) y en la forma de logaritmo natural linealizada, lo que concuerda con una gran parte de los estudios de biomasa (SCHLEGEL, 2001). PEICHL y ARAIN (2007) sugieren la incorporación de la altura como segunda variable cuando las ecuaciones se aplican a árboles de diferentes clases sociales (dominantes, intermedios o dominados). Los resultados obtenidos en los modelos son consistentes si se considera que el número de árboles muestreados es relativamente bajo para la estimación de la biomasa. A pesar de ello, Clark (1979) apud Snowdon et al. (2000) sugiere entre 20 y 40 árboles para una adecuada aproximación a la estimación de la biomasa en árboles individuales. Eficiencia de los modelos Para evaluar la fiabilidad de los modelos de biomasa y volumen se partió aleatoriamente la muestra en dos grupos balanceados y se ajustaron nuevamente los modelo utilizando solo el d (SILVA AYÇAGUER et al., 2001) obteniendo los nuevos estimadores que se presentan en la Tabla 5 para cada grupo. Como se advierte, no se observan variaciones importantes en los estimadores si compara con los detallados en las Tablas 3 y 4, que fueron ajustados con todos los individuos. TABLA 5: Nuevos parámetros para la estimación de biomasa del fuste. TABLE 5: New parameters to estimate stem biomass.
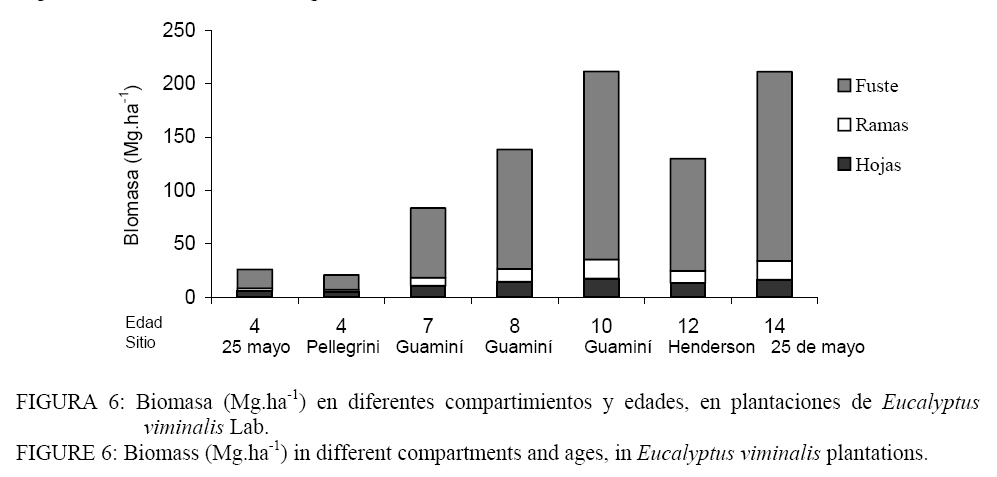
El efecto de remoción al azar de muestras demostró la consistencia o robustez de las funciones ajustadas para el escaso número de individuos muestrados. Biomasa y volumen del rodal Para reconstruir los valores de biomasa de las 22 parcelas de inventario se utilizaron los modelos que incluyen solo el d debido a su comportamiento satisfactorio y a su practicidad de relevamiento. La cantidad de biomasa aérea acumulada por ha (fuste + ramas + hojas) varió desde 20,9 Mg.ha-1 a los 4 años hasta 211,6 Mg.ha-1 a los 14 años de edad (Figura 6). Las observaciones realizadas a nivel de árbol individual se extienden al rodal. Comparando las plantaciones de 4 años (LC y PE) se observó una diferencia del 20% de la biomasa a favor del sitio LC. Otras diferencias importantes atribuidas a condiciones de sitio se observaron en la plantación de 12 años localizada en HE. La biomasa acumulada fue de 138,6 Mg.ha-1, un 40% menos que el rodal de 10 años ubicado en Guaminí (211,6 Mg.ha-1). Las condiciones más desfavorable que presenta HE quedan expuestas al compararlo con los obtenido por POGGIANI (1985). El autor estimó que la biomasa total de Eucalyptus saligna de 11 años en un sitio de baja fertilidad alcanzó las 185 Mg.ha-1. Para calcular el volumen a nivel de rodal se seleccionó la ecuación (5), que incluye el d2 como variable independiente. El volumen acumulado con y sin corteza fue inferior a los 50 m3. ha-1 en las plantaciones de 4 años. Las mayores producciones se lograron en el sitio con 10 años con un volumen superior a los 300 m3. ha-1 ; algo más alto a lo obtenido en la plantación de 14 años. Mantillo y sotobosque La cantidad de MS sobre el suelo varió desde 2,56 Mg.ha-1 en la plantación de 4 años en el sitio LC, hasta 22,7 Mg.ha-1 en la de 14 años (Tabla 6). Se hace particularmente notable el aumento del mantillo, constituido por hojas, ramas y frutos de Eucalyptus, a partir de los 7 años donde ya se ha producido el cierre del dosel. La menor cantidad de mantillo en la plantación de 4 años en el sitio PE, comparado con LC, corresponde claramente a condiciones de fertilidad del sitio, donde las plantas de Eucalyptus se desarrollan lentamente y tienen una cobertura de suelo inferior, con una copa poco desarrollada si se las compara con LC. En estos ambientes de médanos, previo a la plantación suele sembrarse pasto llorón para evitar la erosión, lo cual explicaría la mayor abundancia de sotobosque en esta situación de menor fertilidad. En los sitios DI y AR no se encontró sotobosque debido a la alta densidad de plantación que impide la entrada de luz al estrato inferior. La baja productividad del mantillo en AR se correlaciona con los más bajos valores de biomasa y volumen descriptos previamente. Estos valores de mantillo son comparables a los informados por Pogiani (1985) para un sitio de baja fertilidad. La cantidad de hojas en el piso alcanzó 7,9 Mg.ha-1, algo mayor a AR. Los sitios de mayor producción de biomasa aérea (DI10 y LC14) mostraron una diferencia de ≈ 5 Mg .ha-1. Las diferencias mas importantes fueron encontradas en la presencia de sotobosque que se explica por la mayor apertura de dosel del sitio LC. Se encontró una asociación significativa entre el diámetro medio de la parcela y la MS del mantillo (r: 0,90; p < 0,001) así como también con la MS del sotobosque (r: -0,65; p < 0,001). A edades tempranas, cuando la plantación aún no cerró el dosel e ingresa luz, existe una mayor abundancia de pastos; por el contrario, a edades mayores, hay acumulación de material aportado por la plantación. Los valores de MS encontrados en el presente trabajo se aproximan a lo informado por O´Cconnell y Sankaran (1997) quienes indican que la MS acumulada sobre el suelo (mantillo +sotobosque) en plantaciones tropicales de Eucalyptus spp con edades entre 2 y 27 años varía entre 0,8 y 21,6 Mg.ha-1. En bosques nativos de Eucalyptus en Australia las cantidades fueron desde 12,2 hasta 22,2 Mg ha-1. TABLA 6: Materia seca (Mg.ha-1) en piso forestal de plantaciones de Eucalyptus viminalis. TABLE 6: Dry matter (Mg.ha-1) in forest floor of Eucalyptus viminalis plantations.
Donde: Los valores entre paréntesis indican el desvío standart de la muestra. CONCLUSIONES Este estudio provee una serie de modelos para la estimación del volumen y de la biomasa para Eucalyptus viminalis en una región donde no se dispone de información previa. Esta información resulta fundamental como punto de partida para la diagramación de proyectos relacionados a la cuantificación del carbono. Las ecuaciones de volumen de mejor comportamiento son basados en modelos lineales en su forma normal; con la mejor expresión en aquella que incluye el diámetro en su forma cuadrática. Sin embargo, para la estimación de la biomasa se optan por los modelos en forma de logaritmo natural linealizado. El uso del diámetro como única variable independiente en los modelos de volumen asegura estimaciones confiables además de simplificar la recolección de datos a campo. Los modelos de biomasa, si bien alcanzan buenos comportamientos con el diámetro mejoran con la incorporación de la altura. El análisis de la eficiencia de los modelos demostró que si bien se trabajó con una amplia región geográfica es posible emplear en estas etapas un solo modelo general independientemente del ambiente. La distribución de la biomasa en los individuos sigue el patrón de desarrollo que se encuentra en la bibliografía. La proporción de la biomasa en la copa disminuye con la edad, contrariamente a lo sucedido con el fuste. Se observaron diferencias de producción en función a las unidades de suelo estudiadas. Esto hace notar que si bien se han obtenidos modelos fiables, es necesario ampliar la base de datos en cuanto a edades y distribución de sitios en el área de trabajo. Para futuros estudios de estimación de stock de carbono en el ecosistema los relevamientos deberían incluir la biomasa subterránea (raíz) y el carbono orgánico almacenado en la materia orgánica del suelo. AGRADECIMIENTOS El INTA agradece la colaboración brindada por técnicos de las Agencias de Extensión, a profesionales independientes y a los propietarios de las plantaciones, por el apoyo recibido en tareas de campo y localización de plantaciones. El trabajo fue parcialmente financiado por la Cooperativa de Electricidad CRECES y por el proyecto INTA PNFOR2215. BIBLIOGRAFÍA
Copyright 2008 - Ciência Florestal The following images related to this document are available:Photo images[cf08027f5.jpg] [cf08027f2.jpg] [cf08027f1.jpg] [cf08027f6.jpg] [cf08027f3.jpg] [cf08027f7.jpg] [cf08027f4.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}