 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ciência Florestal, Vol. 18, No. 3, 2008, pp. 353-361 COMUNIDADE DE FUNGOS MICORRÍZICOS ARBUSCULARES EM SOLO CULTIVADO COM EUCALIPTO, PINUS E CAMPO NATIVO EM SOLO ARENOSO, SÃO FRANCISCO DE ASSIS, RS COMUNITY OF ARBUSCULAR MYCORRHIZAL FUNGI IN CULTIVATED SOIL WITH EUCALYPT, PINUS AND NATIVE FIELD, IN SANDY SOIL, SÃO FRANCISCO DE ASSIS, RS Rodrigo Ferreira da Silva1, Zaida Inês Antoniolli2, Robson Andreazza3, João Kaminski4 1Engenheiro Agrônomo, Dr. Professor do Centro Universitário Franciscano, Rua dos Andradas 1416, CEP 97100-
000, Santa Maria (RS). rofesil@bol.com.br Recebido para publicação em 28/08/2007 e aceito em 11/08/2008. Code Number: cf08034 RESUMO O objetivo do trabalho foi caracterizar a comunidade de esporos de fungos micorrízicos arbusculares (FMAs) em cinco áreas sujeitas ao processo de arenização com cultivo de eucalipto (Eucalyptus grandis W. Hill ex Maiden), pinus (Pinus elliottii Engelm. var. elliottii) e campo nativo, localizada no município de São Francisco de Assis, RS. A análise da comunidade de esporos de fungos micorrízicos arbusculares foi mediante a identificação direta, indireta e da determinação do índice de diversidade. As áreas analisadas foram de campo nativo, bosque de eucalipto de 2 e 6 anos e de bosque de pínus de 2 e 12 anos de idade. Os resultados obtidos mostraram que os gêneros mais encontrados foram Acaulospora, Scutellospora e Glomus. A maior diversidade de espécies foi encontrada na área de campo nativo, seguido pela área de eucalipto com 2 anos de idade. A análise de agrupamento mostrou uma semelhança de, no mínimo, 70% para as espécies de FMAs identificados via esporos e 50% para as área estudadas. Palavras-chave: diversidade; Acaulospora; Scutellospora; Glomus. ABSTRACT The aim of this work was to evaluate the community of arbuscular mycorrhizal fungi spores in five areas cultivated with eucalipt (Eucalyptus grandis W. Hill ex Maiden), pinus (Pinus elliottii Engelm. var. elliottii) and native field, in arenization process, located in São Francisco de Assis-RS. The study of arbuscular mycorrhizal fungi spore community was carried out by direct and indirect identification and the diversity index determination. The analyzed areas were native field, 2 and 6 years-old eucalypt field and 2 and 12 years-old pinus field. The obtained results showed that the most frequent genera were Acaulospora, Scutellospora and Glomus. The highest diversity of species was found in area of native field, followed by 2 years-old Eucalyptus area. The Cluster analyses showed a similarity of the minimum 70 % to the species of FMAs identified through spores and 50 % to the areas being studied. Keywords: diversity; Acaulospora; Scutellospora; Glomus. INTRODUÇÃO O termo micorriza define uma simbiose especializada entre raízes e alguns fungos de solo (FRANK, 1885). Os fungos micorrízicos podem ser classificados em fungos micorrízicos arbusculares (endomicorrizas), ectomicorrizas, ectendomicorrizas, micorrizas orquidóides, micorrizas ericóides e micorrizas monotropóides (SMITH e READ, 1997). Os fungos formadores de micorrizas arbusculares (FMAs) pertencem à classe dos Zygomicotina. Os principais gêneros formadores de FMAs são: Acaulospora, Entrophospora, Scutellospora, Gigaspora, Glomus, Archaeospora e Paraglomus (INVAM, 2001). Esses fungos têm sido muito estudados por formar associação simbiótica com a maioria das plantas, desde as Briófitas e Pteridófitas até as Gimnospermas e Angiospermas, ocorrendo em praticamente todas as famílias, com exceção daquelas exclusivamente ectomicorrízicas como, por exemplo, a família Pinaceae (SILVEIRA, 1992). A associação entre espécies vegetais e fungos micorrízicos pode ser importante na recuperação ou reabilitação de solos degradados. Fungos micorrízicos têm sido naturalmente encontrados em solos degradados (MELLONI et al., 2000; CASSIOLATO et al., 2001) e podem contribuir na produção de matéria seca da parte aérea (MELLONI et al., 2000), na sobrevivência das espécies vegetais (MATIAS et al., 2001), no aumento da capacidade de absorção e utilização de nutrientes (SMITH e READ, 1997), na absorção de água (LENY e KRIBUN, 1980), na tolerância a condições desfavoráveis do solo (WILKINS, 1991) e na resistência a microorganismos patogênicos (NEWSHAM et al., 1995). Desse modo, a inoculação de fungos micorrízicos adaptados às condições de solo degradado pode ser uma opção para reutilização dessas áreas. Os FMAs aumentam o vigor e a concentração de nutrientes na planta hospedeira, especialmente em solos deficientes em nutrientes (MOSSE, 1973). O efeito positivo dos fungos micorrízicos arbusculares na nutrição de espécies vegetais em solos degradados e seu efeito na estabilização dos agregados de grãos de areia têm sido relatado (CARNEIRO et al., 1999). Esse efeito pode estar relacionado a uma melhora na eficiência de absorção de P pelas plantas inoculadas com FMAs (CARNEIRO et al., 1999; ALVES et al., 2001). Desse modo, a utilização de FMAs pode ser uma importante opção na nutrição de espécies florestais como, por exemplo, o eucalipto, em áreas sujeitas ao processo de arenização, como as encontradas na região sudoeste do Rio Grande do Sul. As populações de FMAs nativas variam na composição de espécies, porque sua ocorrência é determinada pela vegetação e pelo ambiente. A diversidade biológica das populações nativas de FMAs em ecossistemas não-alterados é variável, apresentando número maior de espécies de FMAs quando o ambiente possuir maior diversidade de plantas. Ecossistemas naturais possuem uma vegetação diversificada, que diminui quando os sistemas de produção agrícola ou florestal são intensivamente manejados, nesse sentido, sistemas de monoculturas tendem a manter apenas uma espécie vegetal por área, apresentando menor diversidade de FMA (COLOZZI-FILHO e BALOTA, 1994). O manejo do solo ou prática agrícola pode alterar a população e diversidade de fungos micorrízicos arbusculares. Sabe-se que os cultivos podem selecionar espécies de FMAs e modificar a composição de sua população nativa (TRUFEM e BONONI, 1985). Portanto, o preparo do solo para plantio é uma prática agrícola que tem efeitos negativos sobre o potencial de inóculo natural do solo. Contudo, os efeitos de sistemas agrícolas sobre a composição e eficiência simbiótica dos FMAs precisam ser melhor estudados. Os FMAs são importantes na fase inicial de desenvolvimento das espécies florestais. Embora a maioria dessas espécies sejam colonizadas pelos fungos ectomicorrízicos, na fase de muda são as endomicorrizas que desempenham um importante papel na absorção de nutrientes (ROJAS e SIQUEIRA, 2000; SANTOS, 2001). Nesse sentido, tem-se observado efeitos benéficos de algumas espécies de FMAs. Por exemplo, Chu et al. (2001), observaram um efeito significativo no crescimento de gravioleira quando inoculada com Scutellospora heterogama e G. margarita. Aumento de crescimento na ordem de 800% foi também observado em mudas de Colvillea racemosa quando inoculadas com uma mistura de FMAs, contendo: Glomus etunicatum, G. margarita e Acaulospora scrobiculata (ROJAS e SIQUEIRA, 2000). Caldeira el al. (1997) observaram maior taxa de sobrevivência de duas leguminosas arbóreas (Copaifera martii Heyne e Dimorphandra macrostachyaBenth.) quando inoculadas com Gigaspora margarita. Então, o conhecimento da ocorrência dos FMAs em áreas degradadas é importante para maximizar os efeitos da simbiose desses fungos com as mudas das espécies florestais usadas no reflorestamento. O conhecimento prévio da ocorrência e diversidade de fungos micorrízicos em áreas degradadas ou sujeitas ao processo de arenização é de fundamental importância nos trabalhos de recuperação ou para reutilização dessas áreas. Assim, este trabalho objetivou caracterizar a comunidade de fungos micorrízicos arbusculares em diferentes áreas sujeitas ao processo de arenização no município de São Francisco de Assis, RS. MATERIAL E MÉTODOS As áreas de estudo localizam-se no município de São Francisco de Assis, região oeste do estado do Rio Grande do Sul. As áreas selecionadas foram campo nativo com predominância de gramíneas, bosque de eucalipto (Eucalyptus grandis W. Hill ex Maiden) de 2 e 6 anos de idade e de pinus (Pinus elliottii var. elliottii Engelm.) de 2 e 12 anos de idade. O plantio das mudas foi realizado sob campo nativo em áreas em processo de arenização, sendo realizado adubação de base, entretanto não foi possível recuperar a informação sobre os valores da adubação aplicada. Determinaram-se os teores de fósforo por Murphy e Riley (1962) e pH em água de amostras de solo das cinco áreas estudadas (Tabela 1). TABELA 1:Valores de pH e fósforo nas amostras de solo das áreas de campo nativo (CN), pinus 2 anos (P2), pinus 12 anos (P12), eucalipto 2 anos (E2) e eucalipto 6 anos de idade (E6). São Francisco de Assis, RS, 2002. TABLE 1: Values of pH and phosphorus in soil sample in native field (CN), pine 2 years old (P2), pine 12 years old (P12), Eucalyptus 2 years old (E2) and Eucalyptus 6 years old (E6). São Francisco de Assis, RS, 2002.
A identificação dos fungos micorrízicos arbusculares foi realizada no Laboratório de Microbiologia e Biologia do Solo Prof. Marcos Rubens Fries do Departamento de Solos, Centro de Ciências Rurais, Universidade Federal de Santa Maria (CCR/UFSM). O processo de identificação constou de duas etapas, uma caracterizada como Identificação Direta (ID) e outra como Identificação Indireta (II). A extração de esporos do solo foi feita pelo método de peneiramento úmido e centrifugação em sacarose, conforme Gerdeman e Nicolson (1963). Para a identificação direta foi realizada uma amostragem composta representativa da área de campo, com 10 subamostras de solo coletadas na profundidade de 0 a 20 cm. Dessa amostra de solo, foi retirada uma amostra de 50 g para a identificação dos esporos. De cada área, foram realizadas cinco repetições, sendo cada repetição considerada uma unidade experimental. Outra parte da amostragem composta (500 g) foi utilizada para a identificação indireta de FMAs. Nessa etapa, instalou-se em casa de vegetação, um cultivo armadilha com trevo vesiculoso (Trifolium vesiculosum L.), com o objetivo da recuperação de espécies de fungos, que não estavam esporulando no momento da coleta, encontrando-se em estágio de hifa ou outro propágulo (SIQUEIRA e FRANCO, 1988). Os esporos extraídos tanto na identificação direta como indireta foram preparados em lâminas com o reagente PVGL e identificados segundo suas características morfológicas (SCHENCK e PEREZ, 1987; INVAM, 2001). Na identificação indireta, utilizaram-se vasos de 1 litro como unidade experimental. Os vasos receberam em seu fundo, uma camada de aproximadamente 2 cm de areia esterilizada. Sobre essa camada, colocaram-se 500 g de solo e sobre esse solo, foi colocado outra camada de 2 cm de areia esterilizada. O uso de areia esterilizada objetivou evitar possíveis contaminações externas. Na areia localizada acima do solo, semearam-se as sementes de trevo previamente desinfestadas em solução de hipoclorito de sódio 5%, durante 1 min. A densidade de semeadura do trevo foi alta, pois o objetivo foi obter um grande volume radicular. Desse modo, não foi seguido nenhum parâmetro para definir a densidade de semeadura. O trevo foi inoculado, com cultura comercial de bactérias fixadoras de nitrogênio simbióticas, do gênero Rhizobium. Os trevos receberam, diariamente, 20 ml de solução nutritiva de Long Ashton (HEWITT, 1966). Além disso, foram aplicados 80 mg kg-1 de solo de nitrogênio na forma de uréia em duas aplicações, aos 40 e 80 dias após a emergência dos trevos. Após quatro meses, coletou-se uma amostra de 50 g de solo para a extração e identificação dos esporos. Após a extração dos esporos, fezeram-se a determinação do número total e a identificação dos principais gêneros e espécies. Determinou-se a diversidade de fungos micorrízicos arbusculares nas cinco áreas avaliadas pelo Índice de Diversidade de Simpson (BEGON et al., 1990). O delineamento experimental foi inteiramente casualizado com cinco repetições. Os resultados de número de esporos e porcentagem de colonização micorrízica foram submetidos à análise de variância e ao teste de comparação de médias (Teste de Tukey), utilizando o programa estatístico SOC desenvolvido pelo Núcleo Tecnológico para Informática NTIA/EMBRAPA (EMBRAPA, 1997). A matriz de similaridade de ocorrência entre as espécies e entre as áreas foi construída usando a análise de Cluster e o agrupamento dos dados foi por joining (tree clustering), por meio de 1–Pearson–r (Single linkage) (STATISTICA, 1993). RESULTADOS E DISCUSSÃO Os gêneros de FMAs, que apresentaram maior percentagem de indivíduos nas cinco áreas avaliadas, tanto na identificação direta quanto na indireta, foram Acaulospora, Scutellospora e Glomus (Tabela 2). A germinação dos esporos está relacionada com o pH do solo e varia entre os gêneros de fungos micorrízicos arbusculares. De uma forma geral, considera-se que os gêneros Gigaspora, Scutellospora e Acaulospora preferem pH mais ácido de 4,0 a 6,0, enquanto Glomus, na faixa de 6,0 a 8,0 (SILVEIRA, 1998). Esse comportamento dos fungos MAs em relação ao pH do solo também foi observado neste trabalho, pois as áreas avaliadas apresentaram pH entre 4,4 e 5,0 e o gênero Acaulospora ocorreu em todas as áreas avaliadas (Tabela 1 e 2). TABELA 2: Percentagem de gêneros micorrízicos arbusculares encontrados nas áreas de campo nativo (CN), pinus 2 anos (P2), pinus 12 anos (P12), eucalipto 2 anos (E2) e eucalipto 6 anos (E6), na identificação direta (ID) e identificação indireta (II), São Francisco de Assis/RS, 2002. TABLE 2: Percentage genera of arbuscular mycorrhizal fungi spores in native field (CN), pine 2 years old (P2), pine 12 years old (P12), Eucalyptus 2 years old (E2) and Eucalyptus 6 years (E6), in direct identification (ID) and indirect identification (II). São Francisco de Assis, RS, 2002.
No campo nativo (ID + II), observou-se um predomínio das espécies S. heterogama, Acaulospora sp. e glomus aggregatum. As áreas de pínus com 2 e 12 anos de idade apresentaram maior porcentagem de A. scrobiculata. Nos eucaliptos com 2 e 6 anos de idade, foi encontrada uma maior porcentagem de S. heterogama, seguida do A. bireticulata no eucalipto com 2 anos e pelo Glomus sp. no eucalipto com 6 anos de idade, apresentando maior número de espécies na área de eucalipto com 2 anos de idade (Tabela 3). Na identificação indireta, observou-se que a área de campo nativo apresentou maior número de espécies (dez), seguido pela área de eucalipto 2 anos (sete). As espécies mais numerosas encontradas na identificação indireta, foram a Acaulospora scrobiculata e a Scutellospora heterogama (Tabela 3). TABELA 3: Percentual de esporos de espécies de fungos micorrízicos arbusculares encontrados nas áreas de campo nativo (CN), pinus 2 anos (P2), pinus 12 anos (P12), eucalipto 2 anos (E2) e eucalipto 6 anos (E6), na identificação direta (ID) e identificação indireta (II). São Francisco de Assis, RS, 2002. TABLE 3: Percentage of spores species of arbuscular mycorrhizal fungi spores in native field (CN), pine 2 years old (P2), pine 12 years old (P12), Eucalyptus 2 years old (E2) and Eucalyptus 6 years old (E6). São Francisco de Assis, RS, 2002.
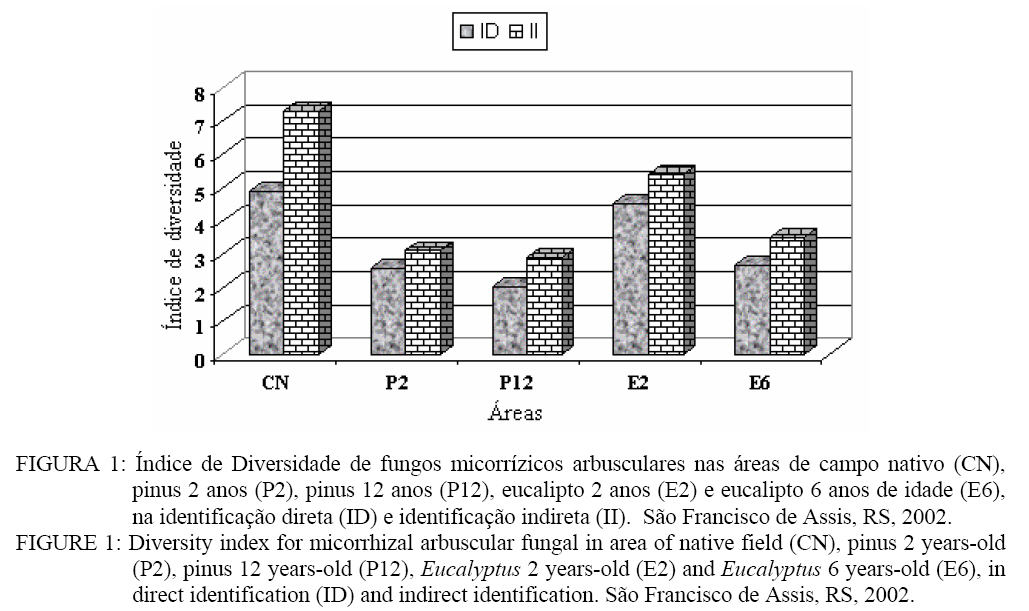
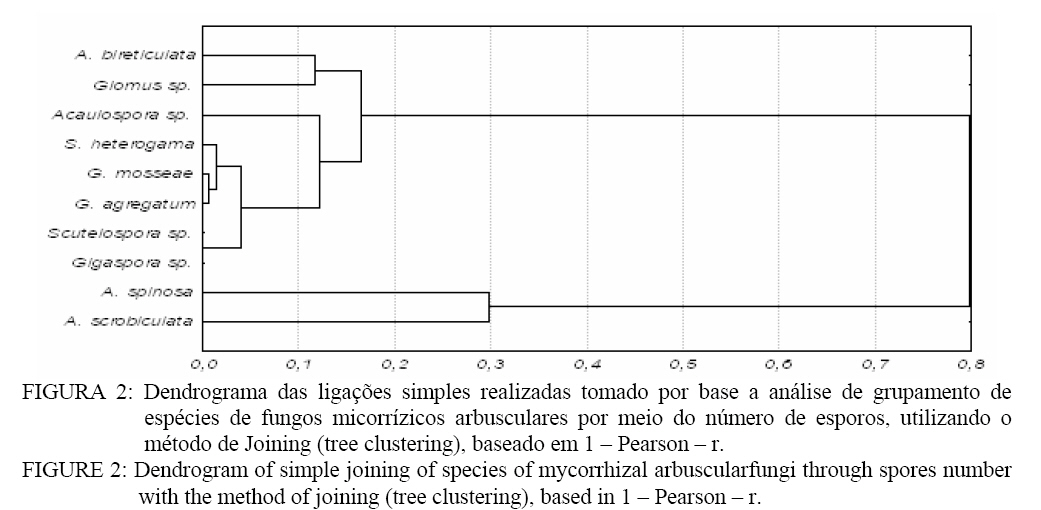
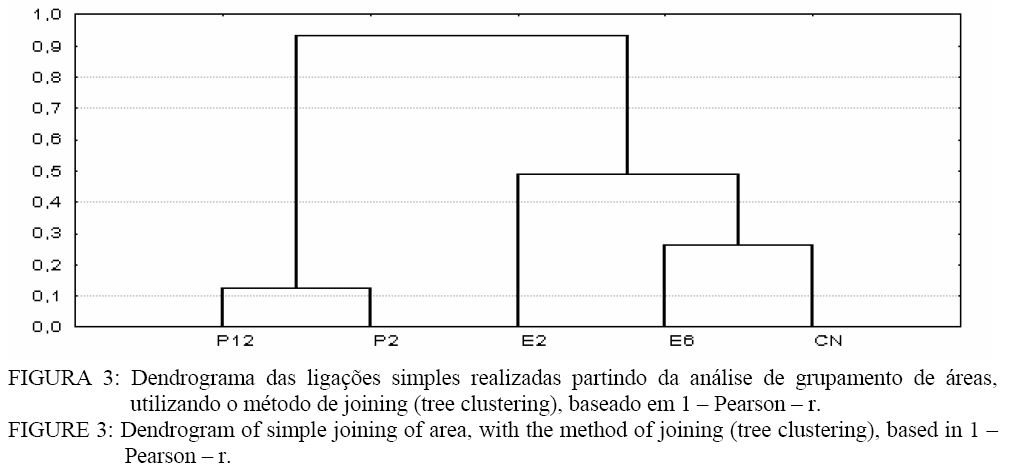
A área de campo nativo (II) apresentou maior porcentagem de S. heterogama, Acaulospora sp. e Scutellospora sp.. Verifica-se que A. scrobiculata apresentou um percentual superior às demais espécies nas áreas de pínus, seguido pela A. spinosa. Nas áreas de eucalipto, ocorreu maior percentual de Acaulospora bireticulata, A spinosa, Acaulospora sp., S. heterogama e Glomus sp..Observa-se ainda a ocorrência de Acaulospora sp. na área de eucalipto com 6 anos, que na identificação direta não havia sido constatada (Tabela 3). Isso indica a presença dessa espécie de FMA que, no entanto, no momento da coleta de solo, ainda não havia esporulado. Portanto, o cultivo de trevo proporcionou a formação e esporulação dos FMAs partindo de outra forma de propágulo. Fatores de solo, como o pH, podem influenciar a associação micorrízica. A variação no pH pode alterar a solubilidade de elementos como Al, Fe, Mn e Cu que, em níveis tóxicos, podem reduzir a germinação de esporos e o crescimento do tubo germinativo, reduzindo a esporulação de FMAs (LAMBAIS e CARDOSO, 1989). Embora os fungos micorrízicos tenham sido encontrados em solos com pH variando de 2,7 a 9,2, ocorrem diferenças entre as espécies e isolados de fungos quanto à capacidade de germinar e colonizar o hospedeiro em função do pH do solo (MALUF et al., 1988; SIQUEIRA et al., 1986). A maior ocorrência de Acaulospora e Scutellospora, encontradas neste trabalho, pode estar relacionada à habilidade dessas espécies em desenvolver-se no pH desse solo que variou de 4,4 a 5,0 (Tabela 1 e 3). A presença de Scutellospora heterogama e das espécies de Acaulospora (Tabela 3) pode estar relacionada ao pH existente no solo das áreas avaliadas (Tabela 1). As espécies de fungos micorrízicos arbusculares pertencentes ao gênero Glomus apresentam maior taxa de germinação, colonização radicular e ocorrência natural em condições de pH próximos do neutro ou alcalino, enquanto que espécies pertencentes à Scutelospora, Acaulospora e Gigaspora desenvolvem-se bem na faixa de pH ácido. Espécies tais como G. margarita, Scutellospora heterogama, A. laevis e A. scrobiculata são predominantes em solos ácidos, germinando preferencialmente a pH inferior a 6,0 (SIQUEIRA e FRANCO, 1988; LAMBAIS e CARDOSO, 1988). Esses resultados se relacionam com as características originais do ambiente nativo, de modo que a acidez pode limitar a distribuição e abundância das espécies de fungos micorrízicos arbusculares, alterando o benefício da simbiose. O maior número de esporos (Tabela 3) foi encontrado na área de pinus com 12 anos de idade, tanto na identificação direta como indireta, sendo que a área com menor número de esporos na identificação direta foi a de eucalipto 2 anos e, na identificação indireta, a eucalipto 6 anos. Esses resultados podem evidenciar a preferência de alguns fungos micorrízicos em colonizarem determinadas espécies de plantas. Algumas plantas, por meio de seus exudatos radiculares, podem estimular a germinação de esporos e o crescimento micelial de FMAs (COLOZZI-FILHO e BALOTA, 1994). Os FMAs não ocorrem de maneira generalizada nas essências florestais, mas são de grande importância na fase inicial de estabelecimento dessas espécies (BELLEI e CARVALHO, 1992). Algumas espécies como, por exemplo, o eucalipto, podem apresentar tanto ectomicorrizas como endomicorrízas (BELLEI e CARVALHO, 1992), contudo, quando adultos, são sobretudo colonizadas essencialmente por fungos ectomicorrízicos (LE TACON et al., 1987). A importância desses fungos se relaciona à fase inicial de estabelecimento dessas plantas, o que torna interessante o estudo dessas comunidades de fungos em áreas cultivadas com plantas florestais como o pínus e o eucalipto. Aplicando o Índice de Diversidade de Simpson na população de esporos de FMAs, pode-se constatar que a área de campo nativo apresentou maior diversidade que as demais áreas avaliadas (Figura 1). Sistemas considerados mais estáveis como o campo nativo, por apresentarem maior diversidade na comunidade de plantas, oportunizam uma maior capacidade de associação para os fungos micorrízicos. Sabe-se que a comunidade de plantas pode alterar a composição de fungos micorrízicos de determinado local (SANDERS e FITTER, 1992). No entanto, a diversidade de fungos micorrízicos arbusculares não segue a de plantas (ALLEN et al., 1995). Assim pode ocorrer uma abundância de FMAs na presença de poucas espécies vegetais ou vice-versa. No cultivo de pínus e eucalipto, houve uma menor diversidade dos fungos micorrízicos em relação ao campo nativo. Sistemas com monocultura, ou com manejo intensivo reduzem a quantidade e a diversidade de FMAs (SILVEIRA, 1998). Dessa forma, áreas em cultivo permanente, como as de pínus e eucalipto, levam à seleção de espécies de fungos micorrízicos, de forma que a diversidade é reduzida, redução essa, que tende a aumentar com o tempo (Figura 1). Normalmente, as essências florestais são colonizadas predominantemente por fungos ectomicorrízicos e não por endomicorrizas (BELLEI e CARVALHO, 1992; LE TACON et al., 1987), o que contribui para uma menor diversidade nessas áreas. As modificações que ocorrem no solo, desde um simples cultivo até um processo de degradação erosiva, podem modificar a predominância de uma espécie fúngica na formação da associação micorrízica. A medida em que a severidade da modificação imposta ao solo aumenta, a diversidade dos FMAs, tende a diminuir (SILVEIRA, 1998). Essa afirmação é confirmada neste trabalho em que a área de campo nativo, onde o solo não sofreu alguma modificação, apresentou a maior diversidade. De acordo com a análise de agrupamento (Figura 2), observa-se a formação de agrupamento em três níveis. O primeiro engloba as ligações com distância inferior a 3% entre as espécies de Glomus mosseae, Glomus agreggatum, Scutellospora heterogama, isso permite inferir que a semelhança entre eles é de 97%. Outro nível é formado pelas espécies de A. bireticulata e Glomus sp. com uma semelhança de 88% e o terceiro, formado pelas espécies de A. spinosa e A. scrobiculata, com uma semelhança de 70%. Observa-se também que, ao excluir da análise o grupo formado pelas espécies de A. spinosa e A. scrobiculata, o restante dos FMAs apresentaram uma distância nas suas ligações inferior a 18%, desse modo, essas espécies teriam uma semelhança de, no mínimo, 82%. Contudo, ao analisar todas as ligações, observa-se que a distância entre as ligações é inferior a 30%. Assim, a semelhança entre o número de esporos nas espécies é de, no mínimo, 70%. Na análise de Cluster para as áreas (Figura 3), observa-se a formação de grupos em dois níveis quanto ao número de esporos. No primeiro nível, encontram-se as áreas de pinus 2 e 12 anos com uma semelhança entre as duas áreas de 88%. No segundo, encontram-se as áreas de campo nativo e eucalipto 6 anos apresentando 75% de semelhança. De uma forma geral, todas as áreas apresentaram ligações inferior a 50%, o que permite dizer que a diferença entre elas é de, no mínimo, 50%. Assim, o trabalho mostra que na área em que o homem teve menor influência, como no campo nativo, a diversidade de FMAs foi maior em relação às outras áreas, podendo estar relacionado a uma maior diversidade de espécies vegetais. Nesse sentido, observa-se também que as áreas mais velhas de pinus e eucalipto tendem a selecionar espécies de FMAs, diminuindo sua diversidade. CONCLUSÕES Os gêneros Acaulospora, Scutellospora e Glomus foram os mais encontrados no solo das áreas de campo nativo e talhões de pínus com 2 e 12 anos e eucalipto com 2 e 6 anos de idade. As espécies encontradas em solo de campo nativo foram: Acaulospora sp., Acaulospora bireticulata, Acaulospora spinosa, Acaulospora scrobiculata, Scutellospora sp., Scutellospora heterogama, Gigaspora sp., Glomus sp., Glomus mosseae e Glomus aggregatum. No solo com pínus, encontroaram-se Acaulospora sp., Acaulospora bireticulata, Acaulospora spinosa, Acaulospora scrobiculata, Scutellospora heterogama e Glomus sp., e nos solos com eucalipto, encontraram-se Acaulospora sp., Acaulospora bireticulata, Acaulospora spinosa, Acaulospora scrobiculata, Scutellospora heterogama, Glomus sp., Glomus mosseae e Glomus aggregatum. A área de campo nativo apresentou a maior diversidade de espécies de FMAs, seguido pelo plantio de eucalipto com 2 anos. A análise de agrupamento revelou uma semelhança de, no mínimo, 70% para as espécies de FMAs identificados via esporos e 50% para as áreas estudadas. REFERÊNCIAS BIBLIOGRÁFICAS
Copyright 2008 - Ciência Florestal The following images related to this document are available:Photo images[cf08034f2.jpg] [cf08034f3.jpg] [cf08034f1.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}