 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ciência Florestal, Vol. 19, No. 2, Abr-Jun, 2009, pp. 139-148 Aporte De Nutrientes E Decomposição Da Serapilheira Em Três Fragmentos Florestais Periodicamente Inundados Na Ilha Da Marambaia, RJ Litter Deposition And Decomposition In Three Forest Fragments Periodically Floodable in The Island Of Marambaia, RJ Ranieri Ribeiro Paula1, Marcos Gervasio Pereira2, Luiz Fernando Tavares de Menezes3 1Estudante de Engenharia Florestal, Universidade Federal Rural do Rio de Janeiro, BR 465, km 7, CEP 23890-000,
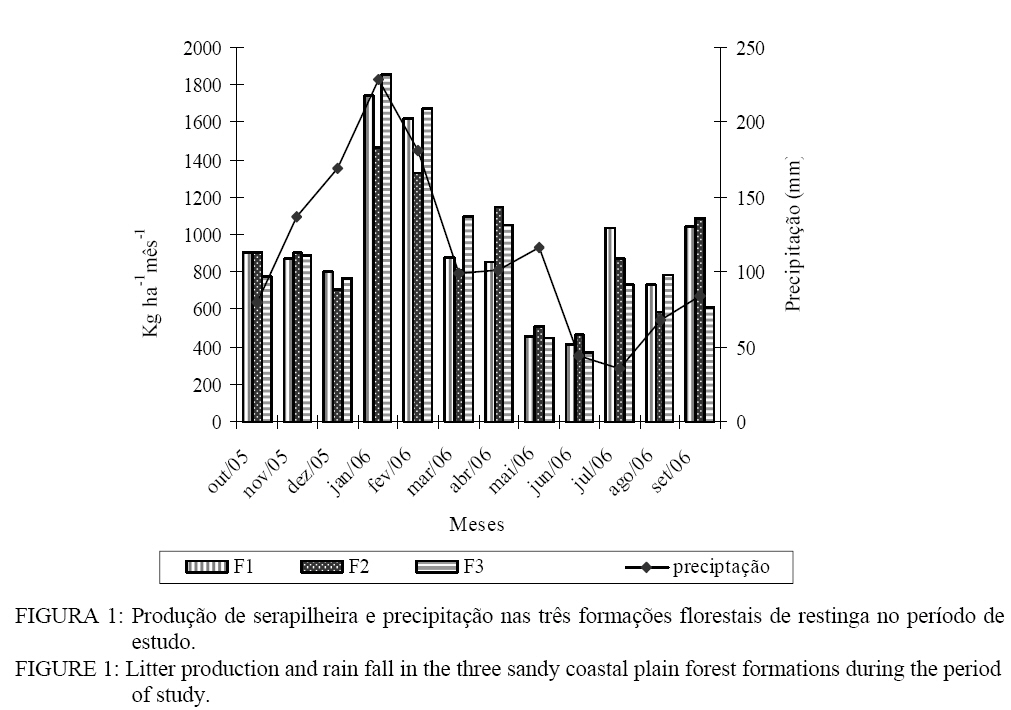
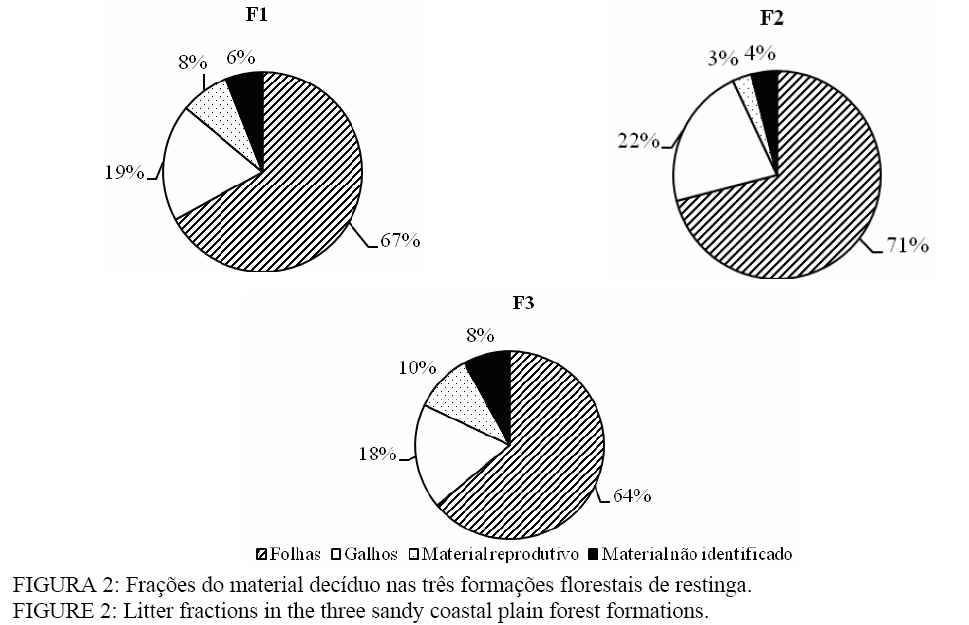
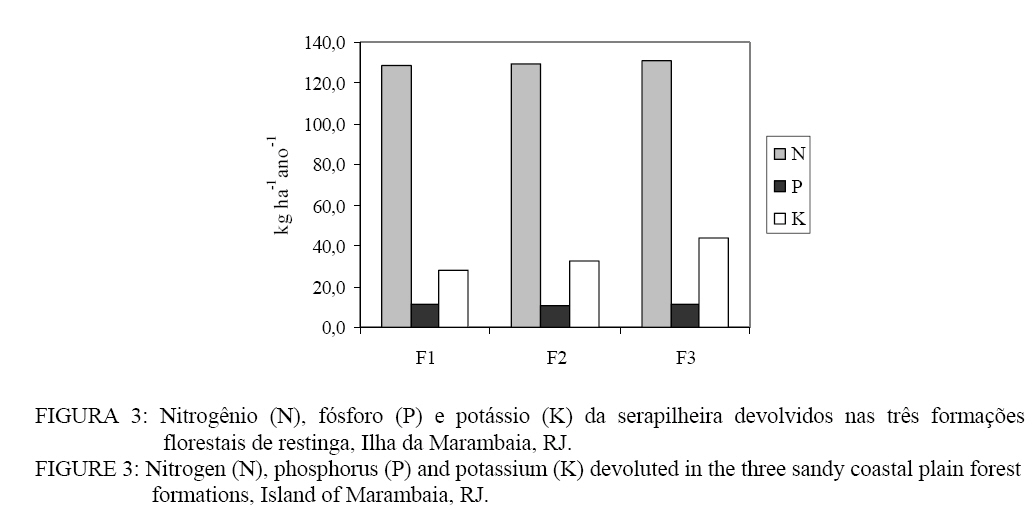
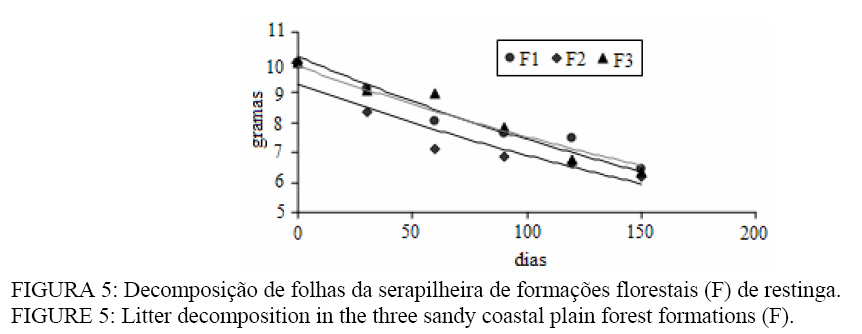
Seropédica (RJ). Bolsista FAPERJ. raniieri@hotmail.com Recebido para publicação em 29/04/2008 e aceito em 12/11/2008. Code Number: cf09014 Resumo O presente estudo foi realizado em três formações florestais de restinga que se encontram periodicamente inundadas, na Restinga da Marambaia, RJ. Para a quantificação da produção de serapilheira foram utilizados dez coletores cônicos por formação, durante outubro de 2005 a setembro de 2006. A avaliação da decomposição se deu por meio de sacolas de decomposição (litter bags), sendo distribuídas 15 sacolas por formação. A produção anual de serapilheira estimada para cada uma das formações florestais, variou de 11,3, 10,8 e 11,1 Mg ha-1ano-1, para F1, F2 e F3 respectivamente, com maiores valores de deposição na estação chuvosa. A fração folhas foi a mais representativa em todas as formações. O nitrogênio foi o nutriente devolvido em maior quantidade a esses solos, 130,8 kg ha-1 ano-1 em F3. A decomposição se apresentou como uma curva do tipo exponencial, sendo a constante de decomposição (k) e o tempos de meia vida foi de 0,0032 g g-1e 217 dias em F2, 0,0031 g g-1e 227 dias em F3 e 0,0028 g g-1; 247 dias em F1 Palavras-chave: folhedo; deposição; restinga; ciclagem de nutrientes. Abstract The litter production and decomposition of three sandy coastal plain forest formations periodically floodable in the Island of Marambaia, Mangaratiba, RJ, were studied from October 2005 to September 2006. For the litter deposition evaluation, 10 litter traps were installed and in each forest formation to quantify the litter decomposition 15 litter bags were allocated in each area. The annual litter production was 11.3, 10.8 and 11.1 Mg ha-1year-1, to F1, F2 and F3, respectively, the highest deposition was verified in the rainy season. To nutrients devolution, the highest value was observed to N, 130.8 kg ha-1 year-1 in F3. The fraction leaves were of higher contribution in relation to the others in all areas. The litter decomposition, evaluated by constant of decomposition (k) and half time life was 0.0032 g g-1and 217 days (F2), 0.0031 g g-1and 227 days (F3) and 0.0028 g g-1 and 247 days (F1). Keywords: litter; depositon; sandy coastal plain; nutrients cycling. Introdução O estudo da ciclagem de nutrientes minerais, via serapilheira, é de fundamental importância para o conhecimento da estrutura e funcionamento de ecossistemas florestais. Parte do processo de devolução da matéria orgânica e de nutrientes para o piso florestal se dá por meio da deposição da serapilheira, sendo esta considerada o meio mais importante de transferência de elementos essenciais da vegetação para o solo (VITAL et al., 2004). A ciclagem de nutrientes em ecossistemas florestais, plantados ou naturais, tem sido amplamente estudada com o intuito de se obter maior conhecimento da dinâmica dos nutrientes nesses ambientes, não só para o entendimento do funcionamento dos ecossistemas, mas também buscando informações para o estabelecimento de práticas de manejo florestal para recuperação de áreas degradadas e manutenção da produtividade de sítios degradados em recuperação (SOUZA e DAVIDE, 2001). Nesse sentido, a produção e a decomposição são parâmetros utilizados, em projetos de recuperação de áreas degradadas, como indicadores de restauração (Arato et al., 2003). A serapilheira é constituída por materiais vegetais depositados na superfície do solo, tais como folhas, cascas, ramos, troncos, gravetos, flores, inflorescências, frutos, sementes e fragmentos vegetais não identificáveis. Sua deposição introduz heterogeneidade temporal e espacial ao ambiente, podendo afetar a estrutura e a dinâmica da comunidade vegetal (Facelli e Pickett, 1991). Vários fatores bióticos e abióticos influenciam a produção de serapilheira, tais como: tipo de vegetação, altitude, latitude, precipitação, temperatura, regimes de luminosidade, relevo, deciduidade, estágio sucessional, disponibilidade hídrica e características do solo. Dependendo das características de cada ecossistema, um determinado fator pode prevalecer sobre os demais (Figueiredo Filho et al., 2003). Após a deposição do material decíduo, o seu acúmulo na superfície do solo será regulado pela taxa de decomposição (Haag, 1985). Alterações físicas no solo provocadas pela liberação de nutrientes e de compostos fitotóxicos podem modificar a atividade de organismos decompositores (Moorhead et al., 1998). A regulação das taxas de decomposição da matéria orgânica depende fundamentalmente das condições físicas e químicas do ambiente e da qualidade orgânica e nutricional do material que é aportado. Associado a esses fatores, a fauna edáfica se encontra inteiramente envolvida nos processos de fragmentação da serapilheira e estimulação da comunidade microbiana do solo (Correia e Andrade, 1999). Os três principais índices que têm sido usados para estimar a decomposição da serapilheira são: a respiração do solo, o valor K, que e a relação entre a quantidade de material que cai do dossel e a que está depositada sobre o solo, e avaliações diretas mediante medidas de perda de massa (litter bags e cover litter) (Correia e Andrade, 1999). Dentre os métodos empregados, apesar das criticas apontadas por alguns autores como Anderson e Ingram (1996) e Rezende et al. (1999), o método do litter bag é o que parece melhor representar com fidelidade a dinâmica de desaparecimento da manta orgânica. Em função do elevado grau de antropização que os remanescentes florestais da Mata Atlântica são submetidos, a restinga da Marambaia, em razão da sua riqueza de formações vegetais, ainda em um bom grau de conservação, apresenta-se com elevado potencial para o desenvolvimento de estudos ecológicos, que possam vir a contribuir para um melhor conhecimento do bioma e funcionar também como ferramenta no planejamento do manejo a ser adotado em programas de recuperação de áreas degradadas. O presente estudo teve por objetivo a avaliação temporal da deposição da serapilheira, bem como a quantificação do material decíduo aportado em formações florestais com diferentes regimes de saturação hídrica na restinga da Marambaia, RJ. Material e Métodos Meio físico A restinga da Marambaia se localiza, em parte, na zona oeste do município do Rio de Janeiro, se estendendo através de um cordão arenoso até parte dos municípios de Itaguaí e de Mangaratiba. Os sítios estudados estão localizados na porção denominada Bravo VI (coordenadas 23°03’S e 43°36’ W). O clima da região classificado segundo Köppen (1948), enquadra-se no macroclima Aw (clima tropical chuvoso com inverno seco), apresentando médias mensais sempre acima 20,0°C e média anual alcançando 23,7º C. Em fevereiro, ocorre a maior temperatura média mensal (26,8° C) e em agosto, a menor (20,9° C). A precipitação é de 1239,7 mm, sendo que a maior parte desta ocorre no verão. Entre os meses de novembro a março os índices pluviométricos médios são sempre superiores a 100 mm, sendo que nessa época, a maior parte das precipitações, é de origem convectiva. Os meses de inverno são mais secos, entre julho e agosto a precipitação média mensal situa-se entre 40 e 55 mm. A umidade relativa do ar durante todo o ano fica próxima a 81%, diminuindo um pouco no inverno. Esses altos valores de umidade são conseqüência da posição geográfica, que é influenciada pela proximidade com o mar e das brisas locais (MATTOS, 2005). Os ventos predominantes na região são oriundos do norte (N), entre os meses de janeiro a julho, com velocidades médias entre 2,2 a 2,7 m s-1, embora, nesse período, ventos provenientes do sul (S) e em março do sudeste (SE) também sejam constatados. Entre agosto e dezembro os ventos provêm do sul (S), com velocidades um pouco superiores, chegando a atingir valores de 3,2 m s-1 na primavera. Durante a entrada de frentes frias ocorrem ventos provenientes do sudoeste (SW) com rajadas superiores a 13 m s-1, e podem ocorrer também ventos locais, os quais antecedem as precipitações convectivas nos meses de verão, podendo atingir velocidades próximas a 30 m s-1 (MATTOS, 2005). Para o estudo foram selecionadas três áreas de florestas periodicamente inundadas (Araújo e Henriques, 1984). Tal denominação é referente à influência do lençol freático nas formações vegetais, sendo que durante os períodos de maior pluviosidade as áreas tornam-se encharcadas. Essas florestas são encontradas nas depressões mal drenadas, sendo comum a ocorrência de Organossolos, relacionados a depósitos paludais ou turfáceos. Essas formações florestais podem ser encontradas ao redor das lagoas Vermelha e Lagoinha, podendo também ocorrer nas depressões do cordão arenoso interno, formadas pelos arrombamentos causados pelo mar e, neste caso, sendo inundadas pelo afloramento do lençol freático (MENEZES et al., 2005). Nessas formações, são encontradas, ocupando o dossel, espécies atingindo 20 m de altura, podendo também algumas espécies emergentes apresentar alturas superiores a 25 m. Dentre elas destacam-se: Tapirira guianensis Aubl. e Calophyllum brasiliense Cambess. Além dessas espécies, marcadamente dominantes do dossel, as mais comuns são Tabebuia cassinoides (Lam.) DC., Anaxagorea dolichocarpa Sprague & Sandwith, Myrcia acuminatissima O. Berg, Myrcia multiflora (Lam.) DC., Annona glabra L. Garcinia brasiliensis Mart., Diospyros ebenaster Retz. As formações florestais são caracterizadas especialmente por apresentarem menor diversidade de espécies, quando comparadas a outras formações do bioma atlântico e um acentuado acúmulo de serapilheira sobre o solo, podendo as camadas atingirem até mais de 20 cm de espessura. Tal acúmulo e diversidade se devem, sobretudo, às condições de anaerobiose, decorrentes da saturação hídrica, causada pela influência do lençol freático. Os sítios de estudo, denominadas como (F1) Formação florestal 1, (F2) Formação florestal 2 e (F3) Formação florestal 3 possuem diferenças principalmente quanto ao grau de saturação hídrica. No sítio F1, nos meses mais secos, é observada a presença de poças esparsas, já em F2, nesse mesmo período, ainda costuma conservar uma lâmina d’água de aproximadamente 4 cm em toda a extensão da superfície do solo (SOUZA, 2007). Diferindo dos demais, o sítio F3 se apresenta saturado durante todo o ano. Métodos Aporte da serapilheira e determinação de N, P e K Em cada uma das áreas, foram instalados dez coletores cônicos, com 0,5 m de diâmetro e distando uma altura média de 0,4 m do solo. As coletas foram realizadas mensalmente durante o período de outubro de 2005 a setembro de 2006. Após coletado, o material foi seco em estufa de circulação de ar forçado a 65°C, por 48 horas, quando atingiu peso constante, sendo estratificado nas frações: folhas, galhos, material reprodutivo e resíduos (material não identificado). Posteriormente o material foi pesado e moído em moinho do tipo Wiley com malha < 2 mm. A produção de serapilheira foi estimada segundo Lopes et al. (2002), a partir da expressão: PS = (Σ PMS x 10.000)/Ac PS = produção de serapilheira (Mg ha-1 ano-1). PMS = produção mensal de serapilheira (Mg ha-1 mês-1). AC = área do coletor (m²). As análises químicas foram realizadas segundo Tedesco et al. (1995) e Malavolta et al. (1989). Com o objetivo de verificar a existência de diferenças significativas entre os aportes mensais de serapilheira nas formações, bem como nos conteúdos de N, P e K, foi uso o teste T de Bonferrone. Decomposição da serapilheira Para a avaliação da decomposição da serapilheira, foram utilizadas 10 gramas de folhas previamente secadas em estufa, sendo posteriormente acondicionadas em sacolas de decomposição (litter bags). Foram instalados 45 litter bags, sendo 15 por formação. Os litter bags foram distribuídos aleatoriamente nas áreas sendo fixados por meio de um fio de náilon a vegetação. A quantificação da taxa de decomposição foi feita mediante medidas de perda de massa, sendo as coletas dos litter bags, realizadas em intervalos regulares de 30, 60, 90, 120 e 150 dias durante os meses de março/2006 a julho/2006. De posse dos valores de perda de massa de cada formação ao longo do período de estudo, foi estimada a constante de decomposição k, conforme Thomas e Asakawa (1993) usando o modelo exponencial apresentado abaixo: Xt = X0 . e-kt. Em que: Xt = peso do material remanescente após t dias; X0 = peso do material seco originalmente colocado nos sacos no tempo zero (PI = 10g). Por meio desse modelo exponencial, foi obtido o valor da constante k para cada unidade de estudo, que indica a velocidade de decomposição. Esse modelo exponencial bem como as curvas que caracterizam a perda de peso (decomposição) da serapilheira foliar foram confeccionados com o auxílio do programa SIGMAPLOT. O tempo de meia-vida (t1/2) desse material foi calculado conforme Rezendeet al. (1999) por meio da equação: t1/2 = ln(2)/ k. Em que: k = constante de decomposição estimada pelo programa citado anteriormente. Resultados e discussão Caracterização da vegetação das áreas Como características gerais, as formações F1 e F2 possuem uma maior diversidade de espécies, sendo que algumas possuem maior valor de importância: Calophyllum brasiliense Cambess. (90,2%), Myrcia acuminatissima O. Berg (48,2%), Tapirira guianensis Aubl. (36,2%), Myrcia multiflora (Lam.) DC. (28,5%), Tabebuia cassinoides (Lam.) DC. (21,1%), em F1 e Calophyllum brasiliense Cambess. (58,5%), Tabebuia cassinoides (Lam.) DC. (44,9%), Tapirira guianensis Aubl. (40,9%), Diospyros ebenaster Retz. (38,0%), Myrcia acuminatissima O. Berg (28,2%), em F2. Já a formação F3, se encontra mais fortemente submetida à saturação hídrica, pois além da influência do lençol freático, ela também e interceptada por um pequeno curso d’água, apresentando provavelmente por esse fator, uma menor diversidade. Dentre as espécies de maior VI, destacam-se: Tabebuia cassinoides (Lam.) DC. (138,1%), Calophyllum brasiliense Cambess. (86,8%), Diospyros ebenaster Retz. (17,2%), Myrcia acuminatissima O. Berg. (16,1%), Inga subnuda Salzm. ex Benth. (10,8%). Produção de serapilheira O aporte de serapilheira e os índices pluviométricos ao longo do período de estudo estão apresentados na Figura 1. Verifica-se que, no período onde houve precipitação mais acentuada (janeiro), foi observada uma maior deposição de serapilheira, havendo correlação significativa entre esses parâmetros, para as diferentes áreas, sendo FF1 (0,65; p < 0,01), FF2 (0,61; p < 0,01) e FF3 (0,75; p < 0,01). Durante o período de estudo, foram observados maiores valores de produção nos meses de janeiro e fevereiro (estação chuvosa) com maior produção mensal em F3, depositando sobre o solo cerca de 1850,6 e 1676,3 kg ha-1mês-1 respectivamente, sendo seguido por F1 com 1742,6 e 1619,7 kg ha-1mês-1 e F2 com 1468,1 e 1328,8 kg ha-1mês-1. Os meses de maio e junho (estação seca) caracterizaram-se pela menor produção de serapilheira. Observando-se um comportamento aleatório na deposição nos outros meses, o que pode ser decorrente da ação de outros fatores. Resultados semelhantes foram obtidos por Pagano (1989) e César (1993) em florestas semidecíduas do estado de São Paulo. Herbohn e Congdon (1993) consideram a ocorrência de maior produção de serapilheira na estação seca influenciada, sobretudo, pelo estresse hídrico. Em florestas semideciduais, a maior queda das folhas é observada no fim da estação seca e início da chuvosa, sendo tal comportamento atribuído a vários fatores como disponibilidade de água (César, 1993; Poggiani e Monteiro Júnior, 1990; Oliveira, 1997), baixas temperaturas (Poggiani e Monteiro Júnior, 1990) e disponibilidade de nutrientes (Morellato, 1992). Burghouts et al. (1994) indicaram a umidade do solo como o principal fator determinante do padrão de deposição foliar verificada em uma floresta da Malásia. Durigan et al. (1996) consideraram indiscutível o efeito do estresse hídrico em floresta ciliar onde o lençol freático é profundo, ressaltando que em áreas onde o lençol é superficial haveria evidências de que a sazonalidade da caducifolia seria determinada por outros fatores, além da disponibilidade de água no solo. A produção total de serapilheira, no período estudado em cada uma das formações, foi de 11,3 Mg ha-1 ano-1, 11,1 Mg ha-1 ano-1e 10,8 Mg ha-1 ano-1, para F1, F3 e F2 respectivamente. Não foi observada diferença significativa pelo teste T de Bonferrone (p < 0,05) para o aporte de serapilheira entre as formações florestais. Resultados semelhantes foram encontrados por Vital et al. (2004), avaliando a produção de serapilheira em uma floresta semidecidual em zona ripária, com totais aportados em torno de 10,6 Mg ha-1 ano-1. Em estudo de uma restinga na Ilha do Mel, Pires et al. (2006) observaram valores de aporte de serapilheira da ordem de a 5,1 Mg ha-1 ano-1, porém segundo Silva e Britez (2005) essa variação em ambientes de restinga deve resultar da grande heterogeneidade ambiental, as quais apresentam distintas composições florísticas, estruturais e funcionais, conforme estejam estabelecidas sobre ou entre cordões litorâneos, além de fatores como o nível do lençol freático e da distância do oceano. Quanto às frações depositadas, observa-se que a maior contribuição para todas as formações foi da fração folhas, a qual apresentou percentuais de 71, 67 e 64% para F2, F1 e F3 respectivamente. Vários estudos demonstraram que um dos principais constituintes da serapilheira são as folhas, que podem contribuir em até 70%, ou mais do material adicionado (MORELATO, 1992; FIGUEIREDO FILHO et al., 2003), o que evidencia a importância desse compartimento na devolução de nutrientes ao piso florestal. Apesar da importância das folhas, dentro dos diferentes compartimentos de uma mesma árvore, podem-se observar diferenças nas concentrações e conteúdos de nutrientes, nos demais compartimentos vegetais, de acordo com sua função bioquímica. Estudando a distribuição de nutrientes em Eucalyptus grandis, Hill (ex-Maiden) Bellote (1979) encontrou maiores concentrações de nutrientes nas folhas, com exceção do cálcio, cobre e manganês, demonstrando que para alguns nutrientes, outros compartimentos podem contribuir significativamente no processo de ciclagem. Essa fração foi seguida em menores proporções pelas frações galhos e material reprodutivo. Na Figura 2, encontram-se as porcentagens para cada fração nas formações estudadas. Para a fração folhas, não foi verificada tendência de maiores valores na área F2. Essa diferença pode ter sido influenciada pelas características estruturais dessa floresta, pois ela apresenta um dossel “mais fechado” e uma formação vegetal mais densa em praticamente toda a sua extensão. Outra provável influência para o aporte observado nessa floresta, seria a relatada por Pires et al. (2006) que descreve a oligotrofia, como um possível impacto que ocorre em áreas próximas ao mar, no qual as gemas das plantas teriam seu desenvolvimento comprometido por causa da ação de areia e sal trazidos pelo vento, o que promoveria uma necessidade contínua de renovação das folhas. Apesar de todas as três áreas estarem submetidas a essas condições, a formação F2 e a que está localizada mais próxima do mar. Para o material reprodutivo os maiores valores foram verificados para as formações F1 e F3, que detém 8 e 10% do total aportado, respectivamente e F2 apenas 3%. Essa diferença pode ser atribuída ao maior número de indivíduos da espécie Calophyllum brasiliense Cambess, a qual, nessas duas formações (F1 e F3), apresenta maior IVI, quando comparadas a F2. Essa espécie detém elevado potencial produtivo de sementes que possuem maior massa, apresentando um fruto de forma globosa, carnosa e de cor esverdeada, com diâmetro variando de 19 a 25 mm. A fração galhos foi mais expressiva em F2 correspondendo a 22% do total aportado. A explicação para esses valores pode ser a mesma atribuída para a fração folhas. Nessa formação, observa-se um dossel mais uniforme e denso, com maior número de galhos podendo, dessa forma, contribuir para um maior aporte dessa fração. Já a fração material não identificado, na qual são incluídas todas as partes vegetais não analisadas separadamente, inclusive animais que podem por ventura serem depositados nos coletores, apresenta uma ligeira diferença entre as formações F3 e F2. Essa variação pode ser decorrente da proporção de cascas encontradas nos coletores dispostos em F3 bem como resíduos orgânicos não identificáveis oriundos do alto do dossel. Essa área possui menor densidade de espécies, tendo também um dossel mais “aberto” estando mais influenciada pelas variáveis climáticas e deposição de materiais, o que pode estar contribuindo para os resultados encontrados. Pires et al. (2006) observaram a participação da fração folhas influenciando em 74,6% do total aportado, sendo seguidos pelas frações ramos (18,1%), órgãos reprodutivos (4,8%) e miscelâneas (2,5%). A distribuição dos percentuais encontrados neste estudo está de acordo com os valores observados na literatura por diferentes autores trabalhando com formações florestais distintas (Bray e Gorham, 1964; Ewel, 1976; Peres et al., 1983), em que a fração foliar é a principal constituinte do total de biomassa produzida, sendo seguida pela fração galhos, estruturas reprodutivas e miscelânea (nesse trabalho, caracterizou como a fração material não identificável). Aporte de N, P e K. Através da Figura 3, verifica-se que o N foi o elemento devolvido em maior quantidade nas diferentes formações. Valores de 130,8 kg ha-1 ano-1 em F3, seguidos por 129,7, kg ha-1 ano-1 em F2 e 128,5 kg ha-1 em F1. Para o potássio, foram quantificados valores de 43,6, 32,7 e 28,3 kg ha-1 ano-1, em F3, F2 e F1 respectivamente. Menores valores de deposição foram observados para o P com totais anuais de 11,7 kg ha-1 em F1, 11,4 kg ha-1 em F3 e 10,9 kg ha-1 em F2. Não
foram verificadas diferenças significativas pelo teste T de Bonferrone (p <
0,05) para o conteúdo de nutrientes nas diferentes formações florestais. Quanto
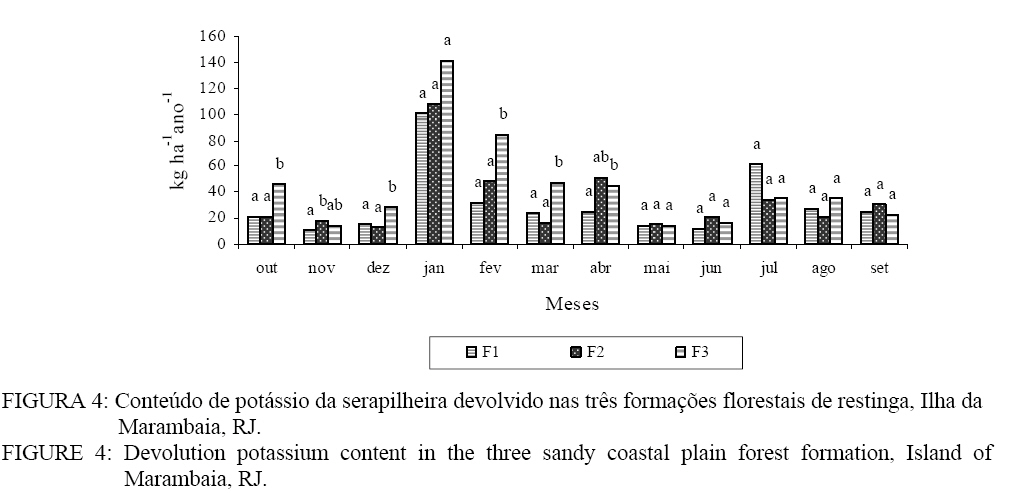
ao aporte mensal de nutrientes nas formações florestais, com exceção ao K, não
foram verificadas diferenças significativas pelo teste T de Bonferrone (p <
0,05). Para o K (Figura 4) foi observada diferença significativa entre F3 e as
demais formações estudadas. Essa diferença pode ser atribuída à influência do
lençol freático marinho, pois essa formação se encontra em uma depressão, o que
possibilita uma maior contribuição de K para as espécies presentes. Nessa
formação, no período seco, o lençol freático fica bem próximo da superfície e
sempre se encontra acima do solo nos períodos de maior precipitação (estação
chuvosa). Estudando a produção e conteúdo de nutrientes na serapilheira em duas florestas tropicais brasileiras, sendo uma delas caracterizada como floresta de restinga, Moraes et al. (1999) encontraram aportes para N, P e K, respectivamente, 28,0, 1,0 e 7 kg ha-1 ano-1. Decomposição da serapilheira Na figura 5, são apresentados os valores de perda de massa da serapilheira foliar durante o período de avaliação. Foi observado em F2 a maior constante de decomposição (k), sendo decomposto 0,0032 g g-1 (p< 0,01; R² = 0,8917) da serapilheira. Seguidos respectivamente por F3 e F1, apresentando valores de k 0,0031 (p < 0,01 R² = 0,9592) e 0,0028 g g-1 (p < 0,01; R² = 0,9643). Para o T1/2 verifica-se que o material de F1 se mostrou mais resistente quanto à decomposição, apresentando um T1/2 maior, em torno de 247 dias. Para F2, foi verificado um menor tempo, 217 dias e em F3 de 223 dias para que seja decomposta metade da serapilheira foliar. Em uma floresta de sucessão secundária espontânea, Fernandes et al. (2006) encontraram valores da constante de decomposição (k) em torno de 0,00194 g g-1 dia e sendo necessários 357 dias para o desaparecimento de metade desse material. Vital et al. (2004), em seu estudo, observaram que eram necessários 150 dias para que ocorresse a decomposição de 50% da serapilheira e 639 dias para que houvesse o desaparecimento de 95% desse material. Foi observado em F2 que a decomposição ocorreu de forma mais rápida no primeiro mês, em que 16,9% do material foliar foi decomposto. Esse comportamento não foi observado nas outras formações, sendo que F3 apresentou 9,5% e F1 somente 8,8% de decomposição no primeiro mês de avaliação. A rápida decomposição que ocorreu no primeiro mês, sobretudo na formação F2, pode ser atribuída ao período em que o material estava mais palatável aos detritívoros (Swift et al., 1979). Já em F1 e F3 esse comportamento não foi observado com grande expressão. As constantes de decomposição se correlacionaram significativamente com a precipitação para todas as áreas, com níveis de significância para F1 (0,57; p < 0,01), F2 (0,54; p < 0,01) e F3 (0,75; p < 0,01). A curva que representa a decomposição é do tipo exponencial, verificando-se que decorridos 150 dias, para todas as formações, a decomposição da serapilheria não ultrapassou os 40% da quantidade inicialmente condicionada nos litter bags. Supõe-se que, nessas formações, a decomposição seja mais lenta, sobretudo em razão da qualidade química do material, como estruturas mais resistentes e compostos mais estáveis e de difícil decomposição como lignina e outros compostos fenólicos (Swift et al., 1979) e as condições de anaerobiose as quais são submetidas essas formações. Tal comportamento pode ser uma estratégia, adotada por esse ecossistema para maximizar o aproveitamento na absorção dos nutrientes e diminuir a perda dos mesmos por lixiviação. Conclusão Os maiores valores de deposição de serapilheira na restinga da Marambaia, RJ ocorreram durante a estação chuvosa (janeiro e fevereiro) e as menores deposições nos meses de maio e junho, caracterizado pela estação seca. A fração folhas é o componente mais representativo, na serapilheira da restinga da Maramabaia, RJ atingindo maior percentual médio em F2 (71%). O N foi o nutriente com maiores valores de aporte nas folhas em todas as áreas de estudo. Foi observada uma baixa taxa de decomposição da serapilheira, sendo verificada correlação entre a decomposição e a precipitação. Referências Bibliográficas
Copyright © 2009 - Ciência Florestal The following images related to this document are available:Photo images[cf09014f4.jpg] [cf09014f3.jpg] [cf09014f5.jpg] [cf09014f1.jpg] [cf09014f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}