 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ciência Florestal, Vol. 19, No. 3, July-Sept, 2009, pp. 267-278 Inoculação de Fusarium oxysporum e Fusarium solani e níveis de sombreamento na erva-mate: influência na severidade da podridão-de-raízes1 Fusarium oxysporum and Fusarium solani inoculation and shading levels in the erva-mate: severity influence of the root-rot Igor Poletto2 Marlove Fátima Brião Muniz3 Denise Ester Ceconi4 Maristela Machado Araujo5 Johnathan Rodrigues6 Ricardo Mezzomo7
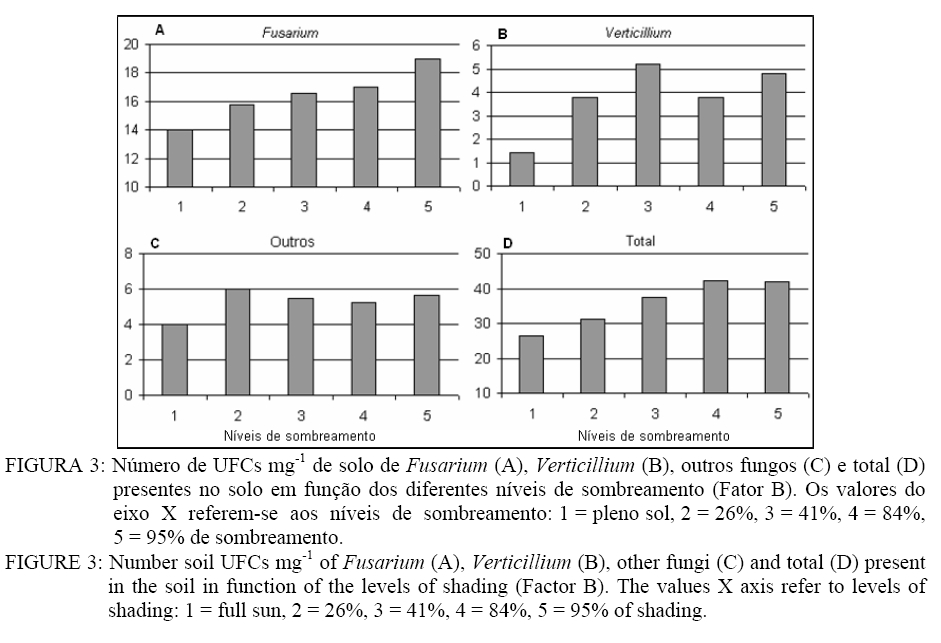
Recebido para publicação em 27/05/2008 e aceito em 31/01/2009. Code Number: cf09026 RESUMO A erva-mate (Ilex paraguariensis A. St.-Hill) é uma espécie amplamente cultivada na Região Sul do Brasil. Com o aumento da área plantada, sobretudo na última década, os problemas fitossanitários causados por pragas e doenças aumentaram e, dentre estes, a podridão-de-raízes vem se tornando um dos principais. Dentre os patógenos causadores dessa doença, Fusarium oxysporum Schlecht. e Fusarium solani (Mart.) Sacc são os principais. Esses fungos provocam sérios danos decorrentes da destruição do sistema radicular, ocasionando, nos casos mais graves, a morte da planta. Suspeita-se que a incidência e a severidade da doença são influenciadas pelo cultivo de ervais em diferentes níveis de sombreamento ou a pleno sol. Assim, foram instalados experimentos na área do Viveiro Florestal pertencente ao Departamento de Ciências Florestais/CCR/UFSM com o objetivo de confirmar tal hipótese. O experimento foi conduzido em um esquema fatorial (2 x 5), composto pela inoculação de Fusarium oxysporum e Fusarium solani e por níveis de sombreamento. Verificou-se que as plantas submetidas aos menores níveis de sombreamento ou a pleno sol ficaram predispostas à doença. Palavras-chave: estresse ambiental; fungos; luminosidade; patógenos de solo. ABSTRACT Ilex paraguariensis A. St.-Hill is a broadly cultivated species in the South of Brazil. With the increasing planted area, mainly in the last decade, fitosanitary problems caused by pest and diseases have increased in this crop, and the root-rot is becoming one of the main problems. Among the pathogens, Fusarium oxysporum Schlecht. and Fusarium solani (Mart.) Sacc are the main ones. These fungi provoke severe damages resulting in the destruction of the root system and, in the most serious cases, they cause the death of the plant. It is suspected that the incidence and severity of disease are influenced by cultivation of Ilex paraguariensis in different levels of shading or full sun. Therefore, experiments were conducted in the Forest Nursery area belonging to Forest Sciences Department/CCR/UFSM with the objective of confirming this assumption. The experiment was conducted in a factorial design (2 x 5), composed by Fusarium oxysporum and Fusarium solani inoculation and by shading levels. It was verified that the plants submitted to lower shading levels or full sun had their predisposed to the disease. Keywords: stresses environmental; fungi; luminosity; soil pathogens. INTRODUÇÃO A podridão-de-raízes é uma importante doença da erva-mate (Ilex paraguariensis A. St.-Hill) causando sérios problemas decorrentes da destruição do sistema radicular das plantas, acarretando em redução na produção na Região Sul do Brasil. Essa doença é causada por várias espécies do gênero Fusarium como: F. oxysporum Schlecht., F. solani (Mart.) Sacc., F. decemcellulare Brick, F. tabacinum (Beyma) W. Gams e F. tricinctum (Corda) Sacc., além de Pythium sp. e Rhizoctonia sp. (POLETTO et al., 2007; POLETTO et al., 2006). Porém, as espécies F. oxysporum e F. solani, são as mais virulentas e possuem uma maior área de abrangência entre as plantações infectadas. Na maioria das vezes, os sintomas se apresentam em reboleiras de tamanho variável, localizadas no meio do erval. Em alguns casos, erveiras mortas ou com sintomas aparecem espalhadas por todo o erval. As árvores apresentam sintomas de amarelecimento ascendente das folhas, estagnação do crescimento e queda excessiva das folhas mais velhas, morte parcial e, em casos mais avançados da doença, a morte total da planta. As raízes superficiais e profundas apodrecem, começando com manchas escuras na superfície. Em casos mais avançados da doença, é possível observar estruturas constituídas de micélio e conídios do fungo (POLETTO et al., 2006). O controle de patógenos de solo, como Fusarium spp., causadores de murchas-vasculares e podridão-de-raízes é muito difícil, pois se desenvolvem no solo e penetram no hospedeiro via sistema radicular. Além disso, são capazes de sobreviver no solo por longos períodos, o que dificulta sua erradicação (BEDENDO, 1995b). Fungicidas químicos não possuem um efeito eficaz e nem desejado pelos agricultores. Em consequência das inúmeras reações químicas que os produtos têm com o solo, não são eficazes e contaminam o meio ambiente e a própria erva-mate, que é consumida “in natura”, portanto, é necessário estudar métodos alternativos de controle desse tipo de patógeno, que visem a desfavorecer o patógeno e favorecer a planta hospedeira, adquirindo resistência à doença. Para Agrios (2005), as doenças resultam da interação patógeno versus hospedeiro sob influência do ambiente. Nas doenças do sistema radicular, a interação patógeno versus hospedeiro ocorre no solo, ambiente altamente complexo e dinâmico onde vários fatores bióticos e abióticos influenciam direta ou indiretamente as doenças. No solo, as variações são regra e não exceção. Essas variações podem ser causadas por fatores abióticos (temperatura, umidade, pH, teor de matéria orgânica, propriedades físicas e químicas) e bióticos (as plantas e a microbiota do solo) (MAFFIA e MIZUBUTI, 2005). As práticas culturais podem influenciar na sobrevivência de patógenos no solo e essas, juntamente com fatores climáticos, edáficos e de cobertura vegetal, são determinantes na distribuição e ocorrência de Fusarium spp. Além disso, nos mais variados patossistemas, as populações de micro-organismos apresentam comportamento dinâmico, e a regulação dessas populações depende das interações entre indivíduos e destes com o ambiente (SAREMI et al., 1999). Modificações no ambiente do erval são alcançadas mediante a adoção ou mudança de algumas práticas culturais como: aumento da cobertura vegetal do solo, consórcio com outras espécies florestais e alteração na quantidade de matéria orgânica do solo. Segundo Bressan (2004), em condições naturais ou agricultáveis, as plantas são frequentemente expostas a algum estresse ambiental, que é definido por um fator externo que exerce uma influência desvantajosa sobre a planta. Alguns fatores dessa natureza como a temperatura do ar, por exemplo, podem se tornar estressantes em poucos minutos, enquanto outros como o conteúdo de água no solo pode levar dias ou até semanas e, ainda, fatores como as deficiências minerais do solo necessitam de maior tempo para se manifestar. A adaptação e a aclimatação ao estresse ambiental resultam de eventos integrados que ocorrem em todos os níveis de organização, desde o anatômico e morfológico até o celular, bioquímico e molecular. As respostas ao estresse incluem mudanças no ciclo e divisões celulares, mudanças no sistema de endomembranas e vacuolização, bem como, alterações na arquitetura da parede celular. Bioquimicamente, as plantas alteram o metabolismo de várias maneiras para acomodar o estresse ambiental, incluindo a produção de compostos osmorreguladores, tais como prolina e glicina betaína (BRESSAN, 2004). A estabilidade de membranas celulares variadas é importante durante o estresse à alta temperatura, assim como durante o resfriamento e o congelamento. Sob temperaturas altas, ocorre um decréscimo na força das ligações de hidrogênio e das interações eletrostáticas entre os grupos polares de proteínas na fase aquosa da membrana. Assim, as temperaturas altas ou muito baixas modificam a composição e a estrutura da membrana e podem causar perda de íons. A ruptura das membranas também causa a inibição de processos como a fotossíntese e a respiração, que dependem da atividade de transportadores de elétrons e enzimas associadas a membranas (BRESSAN, 2004). A erva-mate habita naturalmente o sub-bosque de florestas, como por exemplo, as matas de araucária no Brasil, onde as variações ambientais, durante o ano, são minimizadas dentro do ambiente da floresta. Então, o plantio homogêneo da espécie fora de seu habitat natural pode proporcionar um estresse ambiental, mesmo que mantenha uma boa produção. Esses sintomas são visíveis em ervais puros e se apresentam na forma de alterações no sabor e na cor da folha, diminuição da área foliar e, em períodos de menor precipitação, observa-se estagnação do crescimento e perda excessiva de folhas, chegando até à morte parcial ou total de plantas, o que não ocorre em seu ambiente natural. O conhecimento das condições favoráveis à infecção e colonização é importante para o estabelecimento de medidas de controle, que visam, em última análise, desfavorecer o patógeno em quaisquer fases do estabelecimento da doença. Os fatores do ambiente determinam a distribuição geográfica, a incidência e a severidade da doença sendo, em muitos casos, específicos para o patógeno em questão. Dentre esses fatores, a temperatura é a mais frequentemente correlacionada com a epidemiologia da doença, seguida pela umidade e luz, além da nutrição e do pH do solo (AGRIOS, 2005). Em vista disso, o presente estudo foi desenvolvido com o objetivo de avaliar a severidade da podridão-de-raízes causada por Fusarium spp. em mudas de erva-mate submetidas a diferentes níveis de sombreamento, bem como a população fúngica do solo. MATERIAL E MÉTODOS Localização e características gerais do local dos experimentos: O ensaio foi realizado na área do Viveiro Florestal pertencente ao Departamento de Ciências Florestais (DCF) da Universidade Federal de Santa Maria (UFSM), localizada nas coordenadas 29º42' S e 53º42' W e com altitude de 95 m. Delineamento experimental O delineamento experimental utilizado foi blocos ao acaso em esquema fatorial 2 x 5 (Fator F: inoculação de Fusarium oxysporum e Fusarium solani; Fator B: níveis de sombreamento), totalizando dez tratamentos, com quatro repetições de duas mudas cada. Na Tabela 1, pode-se observar a composição dos níveis de cada fator. Material utilizado Fusarium oxysporum (isolado 12.4a) e Fusarium solani (isolado 4.4) utilizados neste estudo pertencem ao Laboratório de Fitopatologia do Departamento de Defesa Fitossanitária/CCR/UFSM. Esses isolados foram escolhidos pela sua patogenicidade e por estarem presentes em todos os ervais estudados por Poletto et al. (2006). Para obter os níveis de sombreamento, as mudas permaneceram sob capelas de sombrite com diferentes graus de permeabilidade de luz e expostas à luz solar durante todo o dia. A intensidade de sombreamento sob as capelas foi determinada com um luxímetro digital com cinco medidas, em diferentes meses que resultou nas porcentagens de sombreamento apresentados na Tabela 1. O solo (não classificado) utilizado no experimento foi coletado em um horizonte A (0-15 cm), peneirado (malha 0,5 cm) e seco à sombra. Depois de homogeneizado, foi coletada uma amostra e esta analisada quimicamente no Laboratório Central de Análises de Solo da UFSM. De posse dos resultados da análise (Tabela 2), optou-se em não aplicar adubação complementar, pois estava de acordo com a recomendação da Sociedade Brasileira de Ciência do Solo (2004) para a espécie. Instalação do experimento Para acondicionar o solo, foram utilizados sacos plásticos pretos com dimensões 18 x 25 cm. Em setembro de 2006, as mudas de erva-mate, doadas por um viveiro particular do município de Anta Gorda, RS, todas com o mesmo tamanho (quatro folhas e 5 cm de altura partindo do colo), foram repicadas e permaneceram por um mês em casa de vegetação e, em seguida, foram transferidas para as capelas onde permaneceram até o final do experimento. A irrigação foi realizada manualmente com água de poço artesiano. Produção do inóculo Os isolados de F. oxysporum ou F. solani utilizados neste estudo foram oriundos de plantas de erva-mate doentes e, identificados em estudos anteriores por Poletto et al., (2006), conservados, por dois anos, em frascos com sementes de sorgo e solo esterilizados, em temperatura ambiente. Porções dessas sementes foram retiradas dos frascos e transferidas para placas de Petri contendo batata-dextrose-ágar (BDA) e mantidas por 10 dias a 24°C com fotoperíodo de 12 h em câmara BOD. Porções das colônias crescidas nas placas foram transferidas para frascos de Erlenmeyer contendo arroz (sem casca, tipo parboilizado) previamente umedecido e autoclavado (30 min a 121°C a 1 atm). Depois de inoculados, os frascos permaneceram na câmara de incubação a 24°C com fotoperíodo de 12 h por 20 dias em câmara BOD. Após esse período, o arroz foi colocado em um recipiente com água esterilizada (1000 mL de água para cada 200 g de arroz) e agitado por 10 min, a suspensão resultante foi coada e contou-se o número de esporos, com auxílio da câmara de Neubauer em um microscópio. A suspensão apresentou 4,30 x 107 esporos mL-1 para F. oxysporum e 2,99 x 107 esporos mL-1 para F. solani. Após 5 meses, a repicagem das mudas foram inoculados 20 mL da suspensão de esporos por vaso (10 mL de suspensão para cada espécie), incorporados nos dois primeiros centímetros da superfície do solo, sem causar ferimento ás raízes e, imediatamente, este foi umedecido com água. Avaliações em plantas Após a inoculação do F. oxysporum e F. solani, até o final do experimento (9 meses após a inoculação), foram realizadas avaliações semanais analisando-se, visualmente, o aparecimento de sintomas na parte aérea de cada muda como: clorose, necrose, murcha, amarelecimento, encarquilhamento e queda de folhas, atribuindo notas para o grau de severidade de cada um (Tabela 3). As raízes de cada muda foram lavadas em água corrente e, em seguida, analisadas visualmente quanto à presença de necrose, atribuindo notas para o grau de severidade (Tabela 3). Foi retirada uma amostra do caule e da raiz de cada muda, colocada em câmara úmida nas caixas “gerbox” e mantida na câmara de incubação a 24°C com fotoperíodo de 12 h por 6 dias. Após esse período foi observado o desenvolvimento ou não de F. oxysporum e F. solani nas amostras, com auxílio de um microscópio estereoscópico. Avaliações no solo Coletou-se uma amostra de solo de 200 g, composta por todos os tratamentos, e encaminhou-se ao Laboratório Central de Análises de Solo da UFSM para análise química. Uma amostra (50 g) de cada tratamento (composta por uma alíquota das cinco repetições) foi coletada para a determinação e contagem da população fúngica do solo. Estas foram embaladas individualmente em sacos plásticos (dimensões 10 x 15 cm), que foram vedados e armazenados em geladeira a 4°C imediatamente após a coleta até sua análise. Para a determinação da população fúngica, 10 g de solo de cada tratamento foram diluídos em 90 mL de água destilada e esterilizada, juntamente com 2 gotas do emulsificante (Tween 80) para evitar a decantação de esporos, agitados por 2 min e, partindo dessa suspensão-mãe, foram feitas outras diluições seqüenciais 10-2, 10-3 e 10-4. Em cada diluição, foi retirada uma alíquota de suspensão e inoculada em três placas de Petri (0,5 mL placa-1) contendo meio BDA, suplementado com 4,0 mg de estreptomicina (concentração de 70%) /100 mL de meio, para evitar o crescimento de bactérias. As placas inoculadas foram mantidas em câmara de crescimento por 6 dias a 24ºC, com fotoperíodo de 12 h e, após esse período, foram quantificados e identificados os fungos presentes, em nível de gênero, pelas observações das placas diretamente no microscópio estereoscópico e com confecção de lâminas para observação em microscópio, com base na bibliografia especializada (BARNETT e HUNTER, 1999). O número de unidades formadoras de colônia (UFCs) de cada gênero fúngico presente foi determinado através das seguintes equações (CARTER, 1993): FD = Di × Ds ×Q Em que: FD = Fator de diluição; Di = Diluição inicial; Ds = Diluição subseqüente; Q = Quantidade colocada na placa. 1 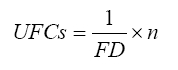
Em que: UFCs = Unidades formadoras de colônias por grama de solo; n = Número de colônias contadas na placa. Procedimento estatístico Os dados obtidos da contagem da população fúngica do solo foram transformados para √(x + 0,5) para aproximação da curva normal e submetidos à análise da variância para verificação da significância e, em caso positivo, foi aplicada análise de regressão ou comparação de médias com teste de Tukey a 5% de probabilidade de erro. Para essas análises, foi utilizado o software estatístico SANEST (ZONTA et al., 1984). Os valores das notas atribuídas a cada sintoma foram submetidos à análise da variância pelo teste de Friedman e comparação entre tratamentos pelo teste da Soma das Ordens de Wilcoxon (CARNELUTTI FILHO et al., 2001). Para essas análises, foi utilizado o software estatístico STATISTICA 6.0. RESULTADOS E DISCUSSÃO Propriedades químicas do solo A Tabela 4 apresenta a análise química do solo antes e ao final do experimento. Observa-se que houve pouca redução nos teores de nutrientes encontrados no solo do início para o final do experimento, portanto, não houve impedimentos nutricionais que poderiam interferir no bom desenvolvimento da espécie, estando os teores dentro do recomendado para a erva-mate. A redução nos teores de nutrientes, observados ao final do experimento, é decorrente da absorção e incorporação na biomassa da erva-mate. O fósforo, o potássio e o magnésio são considerados altos antes e ao final do experimento e, o Ca passou a ser médio. A quantidade de matéria orgânica é considerada média antes da instalação do experimento e continuou média até o final, com um valor muito próximo ao encontrado no início (SOCIEDADE BRASILEIRA DECIÊNCIA DO SOLO, 2004). Efeito dos tratamentos na severidade da doença Na análise dos sintomas da parte aérea e radicular apresentados pelas mudas, foram observadas respostas isoladas nos diferentes tratamentos. Na Figura 1, observam-se os gráficos das notas atribuídas aos sintomas necrose das folhas, clorose, amarelecimento, queda de folhas e necrose das raízes, nos diferentes tratamentos. A análise da variância pelo teste de Friedman mostrou que houve efeito significativo dos tratamentos para todos os sintomas observados. O teste da Soma das Ordens de Wilcoxon, para comparação entre tratamentos, não foi adequado para esse tipo de experimento. Por esse teste, não foi possível verificar diferenças na maioria dos tratamentos, provavelmente, em decorrência do pequeno intervalo de notas (0 a 5) atribuídas aos sintomas, e pelo fato de, na maioria das vezes, as cinco repetições de cada tratamento receberem notas heterogêneas. Para a necrose de folhas (Figura 1 A), os sintomas se manifestaram mais intensamente nos tratamentos com menores níveis de sombreamento, sendo os tratamentos do nível 1 com as maiores notas e, decrescendo sucessivamente, até o nível 5. A análise da variância mostrou que o efeito dos tratamentos foi significativo (p < 0,00008). O sintoma clorose das folhas (Figura 1 B) foi pouco evidente nos níveis 3 e 4 de sombreamento e, nos níveis 1 e 2, praticamente não ocorreu. Isso se deve, provavelmente, à maior severidade da necrose e da queda de folhas. Já para o nível 5, houve ausência do sintoma. A análise da variância mostrou efeito significativo dos tratamentos para este sintoma (p < 0,03935). Quanto ao sintoma amarelecimento das folhas (Figura 1 C), a maior severidade foi verificada nos tratamentos dos níveis 2, 3 e 4 de sombreamento. No nível 1, o sintoma quase não foi observado, provavelmente, pela maior severidade da necrose e queda de folhas. Nas mudas submetidas ao maior sombreamento (nível 5), o sintoma de amarelecimento esteve presente, com baixa intensidade, quando foi inoculado com os isolados de Fusarium. A análise da variância mostrou que houve efeito significativo dos tratamentos (p < 0,00087). A maior diferença na severidade deste sintoma foi observada no nível 4 de sombreamento, com nota 85% maior no tratamento inoculado em relação ao não inoculado. A maior severidade de queda de folhas ocorreu nos tratamentos do nível 1 de sombreamento. Nesse nível, as mudas inoculadas com Fusarium chegaram a apresentar nota 5 (100% de queda de folhas), diminuindo sucessivamente nos níveis 2 e 3 até, praticamente, não ocorrer sintoma nos níveis 4 e 5 (Figura 1 D). A análise da variância mostrou que houve efeito significativo dos tratamentos (p < 0,00012). A severidade dos sintomas necrose de folhas, clorose, amarelecimento e queda de folhas, observados nos tratamentos inoculados com Fusarium foi, em média, 24, 56, 38 e 21% respectivamente, maior que nos tratamentos não inoculados. Na análise do sistema radicular, observou-se necrose e as maiores notas foram atribuídas aos tratamentos do nível 1 de sombreamento, diminuindo, sucessivamente, à medida que o sombreamento aumentou, chegando a nota 0 no nível 5 (Figura 1 E). Em média, as notas atribuídas à severidade do sintoma nos tratamentos com inoculação de Fusarium foram 40% maiores que os não inoculados. Pela análise da variância verificou-se que houve efeito significativo dos tratamentos (p < 0,00017). Na Figura 1 F, observa-se o gráfico representando a soma total dos sintomas. Nota-se que os tratamentos do nível 1 apresentaram a maior severidade, diminuindo sucessivamente com o aumento do sombreamento. O tratamento inoculado com Fusarium e submetido ao nível 5 de sombreamento praticamente não apresentou sintomas, recebendo a nota 0,4 e sua testemunha, sem inoculação, não apresentou nenhum sintoma. Em média, os tratamentos com inoculação de Fusarium spp. obtiveram notas 32% maiores que os não inoculados. Houve efeito significativo dos tratamentos (p < 0,0001) pela análise da variância. Observa-se que o melhor desenvolvimento morfológico das mudas ocorreu nos maiores níveis de sombreamento e ao contrário, nos menores níveis, estas apresentaram desenvolvimento reduzido e sintomas de estresse ambiental (dados ainda não publicados). No geral, observou-se uma tendência de aumento da severidade dos sintomas causados por Fusarium dos maiores para os menores níveis de sombreamento, indicando que as plantas estressadas ficam mais suscetíveis ao ataque do patógeno. Esses resultados vêm ao encontro aos de Bedendo (1995a), afirmando que o ambiente é um componente relevante na interação patógeno-hospedeiro-ambiente, podendo, inclusive, impedir a ocorrência da doença, mesmo na presença de hospedeiro susceptível e do patógeno. Doenças altamente destrutivas, em ambiente favorável, podem passar despercebidas, sob certas condições ambientais. A influência da luminosidade e da temperatura também foi observada na severidade de Fusarium graminearum Schwabe no biocontrole de Egeria densa Planch. e E. najas Planch. O aumento da temperatura ambiental, associada ao menor período de luminosidade, aumentou a severidade da doença (BORGES NETO et al., 2004). Para Bedendo (1995a), os fatores do ambiente podem determinar o grau de predisposição do hospedeiro. Por outro lado, tais fatores também podem ter efeito direto ou indireto sobre o patógeno favorecendo ou desfavorecendo sua sobrevivência Na literatura, os trabalhos encontrados, envolvendo o sombreamento relacionado à incidência e severidade de doenças, são escassos e envolvem patógenos da parte aérea. Não foi possível encontrar trabalhos que envolvam patógenos radiculares para discutir com os resultados encontrados no presente estudo. Efeito dos tratamentos na população fúngica do solo A quantidade de radiação solar exercida sobre o solo determina variações diárias e sazonais na temperatura deste, sobretudo nas camadas superficiais. Assim, várias pesquisas têm demonstrado a existência de uma estreita correlação entre atividade biológica e a temperatura do solo. O efeito da luz sobre os microorganismos do solo é exercido diretamente para aqueles presentes na superfície, ou indiretamente, por meio das plantas, para aqueles que habitam as camadas mais profundas do solo, na região da influência rizosférica ou em simbiose com as raízes, dependendo dos produtos fotossintetizados que a planta destina à raiz, seja diretamente ou como exsudatos (TSAI et al., 1992). Quanto à população fúngica no solo ao final do presente experimento, os gêneros identificados nas diluições foram: Fusarium, Penicillium, Aspergillus, Verticillium, Rhizopus, Rhizoctonia, Pseudobotrytis, Nigrospora, Paecilomyces, Epicoccum, Monilia, Trichoderma, Gliocephalis, Curvularia e Dichobotrys. Segundo Stamford et al. (2005), os fungos são encontrados no solo com variação entre 104 a 106 UFCs g-1 de solo. Além da grande importância como responsáveis pela maior parte das doenças das plantas, realizam funções como: imobilização, adição de matéria orgânica, solubilização de nutrientes, agregação do solo e ação predatória. Segundo os mesmos autores, os fungos mais comuns encontrados no solo são dos gêneros Aspergillus, Cladosporium, Fusarium, Helminthosporium, Mucor, Penicillium, Phytophthora, Pythium, Plasmodiophora, Rhizopus, Sclerotium, Scopulariopsis, Thielaviopsis e Trichoderma. Por se apresentarem em maior número, as populações de Fusarium e Verticillium, encontradas neste estudo, foram analisadas estatisticamente, as demais foram somadas compondo a variável “outros”, a variável “n. de gêneros” foi composta pela contagem do número de gêneros presentes em cada tratamento e a variável “total” foi somado o número de UFCs de todos os gêneros presentes. Na Tabela 5, são apresentados os valores de significância pela análise da variância para essas variáveis. Observa-se, na Tabela 5, que as variáveis Fusarium, outros e número de gêneros não apresentaram significância para os fatores F (inoculação ou não de Fusarium) e B (níveis de sombreamento), nem na interação entre ambos, diferentemente da população de Verticillium, que apresentou significância em ambos os fatores e também na interação. Para a variável população total, a significância estatística foi somente para o fator B. A população de Fusarium (Figura 3 A) apresentou uma tendência de resposta linear positiva aos níveis de sombreamento (Tabela 6). Em média, a população desse fungo apresentou 16,5 UFCs mg-1 de solo, sendo que, nos tratamentos inoculados, o número de UFCs foi 16,6% maior que nos não inoculados, em média. Nesse caso, pode-se afirmar que o efeito dos níveis de sombreamento teve pouca influência sobre a população de Fusarium spp. do solo. Uma hipótese para esse resultado, seria a de que, mesmo com as diferenças ambientais proporcionadas pelos níveis de sombreamento, o ambiente do solo não foi afetado suficientemente para favorecer ou desfavorecer Fusarium spp., ou então, esse fungo possui uma alta plasticidade e/ou capacidade de adaptação a ambientes diversos. Em contrapartida, os maiores níveis de sombreamento, proporcionaram uma condição ambiental que favoreceu a erva-mate perante o ataque de Fusarium spp. Para a população de Verticillium (Figura 3 B), houve diferença significativa, pelo teste de Tukey, na média entre os níveis do fator F (inoculação ou não de Fusarium spp.). Os níveis inoculados apresentaram uma população 51% menor que os tratamentos não inoculados. Houve uma resposta linear positiva do número de UFCs aos níveis de sombreamento (Tabela 6). Em média, o número de UFCs aumentou de 1,4 UFCs mg-1 de solo nos tratamentos a pleno sol, para 4,8 UFCs mg-1 de solo nos mais sombreados, com um aumento de 70%. Nota-se que a população de Verticillium, diferentemente de Fusarium, tem sua população reduzida partindo do nível 3 de sombreamento, sendo influenciada significativamente. A soma da população dos demais fungos (outros, Tabela 6) apresentou significância somente no fator F (inoculação ou não de Fusarium spp.). A comparação de médias pelo teste de Tukey, mostrou que houve superioridade no número de UFCs dos tratamentos não inoculados em comparação aos inoculados, em média 39%, (4 UFCs mg-1 de solo e 6,5 UFCs mg-1 de solo respectivamente). O número de gêneros identificados não apresentou significância em resposta aos fatores F e B, nem na sua interação. O mínimo e o máximo de gêneros identificados nos tratamentos foram de 3 e 9, respectivamente. A população fúngica total do solo (Figura 3 D) somente apresentou significância para o fator B, apresentando resposta linear positiva aos níveis de sombreamento (Tabela 6), obtendo em média, 26 UFCs mg-1 de solo nos tratamentos a pleno sol para 42 UFCs mg-1 de solo, nos mais sombreados, um aumento de 37%. Esses resultados indicam que, em geral, a população fúngica do solo desenvolveu-se melhor em uma condição de maior sombreamento. Para sobreviverem no solo, ambiente em constante modificação, alguns fungos se adaptam para utilizar as mais diversas fontes de energia e sob as mais diversas condições ambientais como, concentrações de oxigênio, umidade, temperatura, etc. (NEVES, 1992). A ação do ambiente sobre o patógeno é exercida de diferentes formas, podendo interferir nos processos de sobrevivência, disseminação, infecção colonização e reprodução. Além dessa interferência, o ambiente pode atuar indiretamente, alterando populações ou atividades microbianas que ocorrem no solo e que possam ter uma relação sinergística ou antagônica em relação a um agente patogênico (BEDENDO, 1995a). Nas amostras de raízes e parte aérea, colocadas em câmara úmida, observou-se que em todos os tratamentos inoculados ocorreu crescimento de Fusarium, partindo dos tecidos internos das mudas, após 6 dias de incubação. Os maiores níveis de sombreamento proporcionaram efeito positivo no desenvolvimento da erva-mate e na resistência ao patógeno, mas não proporcionaram impedimentos para este penetrar e colonizar os tecidos internos da planta. Nos tratamentos sem inoculação, não houve desenvolvimento de Fusarium spp. O sombreamento pode ser uma prática cultural que pode ser adotada com a finalidade de criar uma condição ambiental que desfavoreça o patógeno ou favoreças o hospedeiro, com o objetivo de impedir o aparecimento de doenças ou de mantê-las em baixos níveis (BEDENDO, 1995a). Em plantações de erva-mate, o sombreamento pode ser alcançado por meio do consórcio com outra essência florestal ou em enriquecimento de mata nativa. CONCLUSÕES Pelos resultados obtidos, pode-se concluir que: O patossistema erva-mate – Fusarium spp. é influenciado pelos níveis de sombreamento. A erva-mate submetida a pleno sol ou aos menores níveis de sombreamento apresentou maiores níveis de sintomas de podridão-de-raízes. O maior sombreamento proporcionou um aumento da população total de fungos no solo. REFERÊNCIAS BIBLIOGRÁFICAS
Copyright © 2009 - Ciência Florestal The following images related to this document are available:Photo images[cf09026t4.jpg] [cf09026t5.jpg] [cf09026t2.jpg] [cf09026t1.jpg] [cf09026t3.jpg] [cf09026f1.jpg] [cf09026f3.jpg] [cf09026t6.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}