 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ciência Florestal, Vol. 19, No. 4, Oct-Dec, pp. 375-382 Concentração e retranslocação de nutrientes em acículas de Pinus taeda L. Nutrients concentration and retranslocation in the Pinus taeda L. needles Márcio Viera1 Mauro Valdir Schumacher2
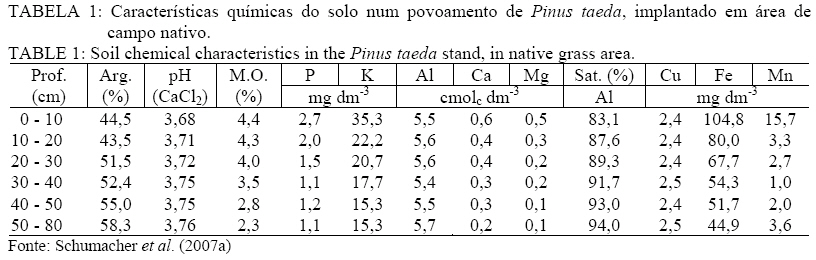
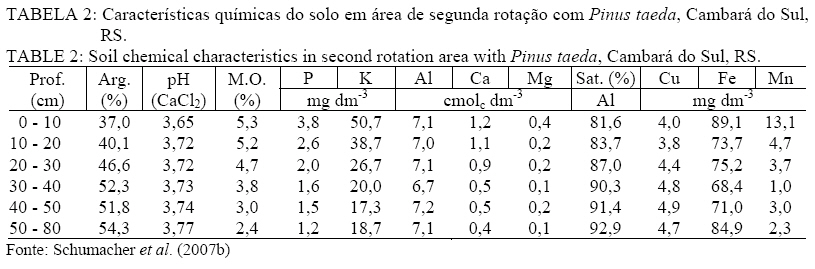
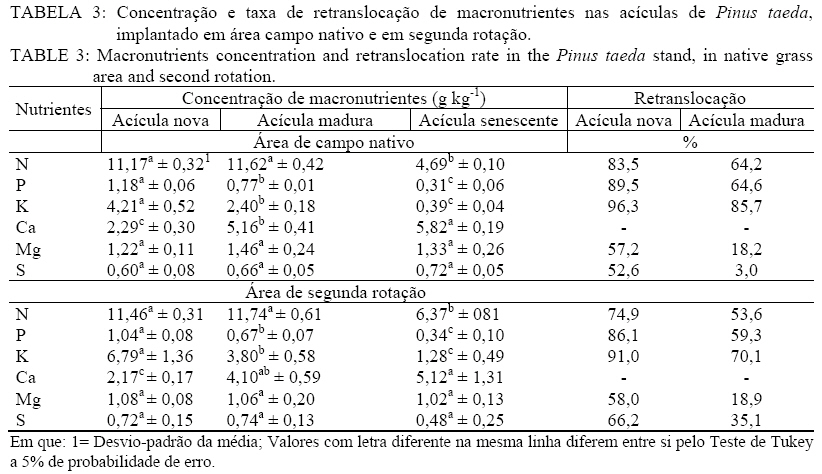
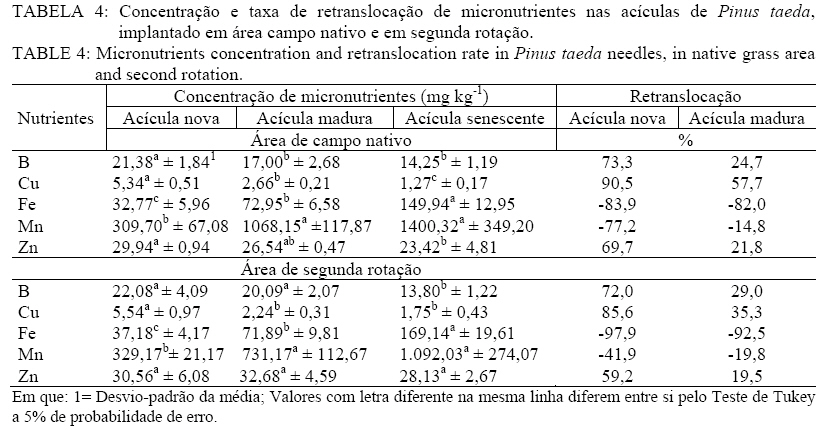
Recebido para publicação em 18/03/2009 e aceito em 19/10/2009. RESUMO Objetivou-se avaliar a concentração e retranslocação de nutrientes em acículas de Pinus taeda L.. O estudo foi realizado em dois povoamentos, um implantado em área de campo nativo e outro em área de segunda rotação, com mesma espécie e mesma idade (7,5 anos) no município de Cambará do Sul, RS. As acículas foram coletadas nas plantas nos quatro pontos ortogonais (sul, norte, leste e oeste), amostrando-se acículas jovens, maduras e senescentes. O material foi seco em estufa, moído e analisado quimicamente (macro e micronutrientes). As concentrações de N, P, K, B, Cu e Zn diminuíram, de Ca, Fe e Mn aumentaram e o Mg e S mantiveram-se constantes com o aumento da idade das acículas. A taxa de retranslocação (acícula senescente-nova) foi superior a 50% para a maioria dos nutrientes, com exceção ao Mn e Fe, que apresentaram efeito cumulativo e o Ca, usado como elemento referência pela sua baixa mobilidade. Palavras-chave: redistribuição de nutrientes; ciclo bioquímico; estádio fenológico. ABSTRACT Aiming at evaluating nutrients concentration and retranslocation in the Pinus taeda L. needles, this study was developed in two stands, in native grass area and in second rotation area, with same species and same age (7.5 years old) in Cambará do Sul, RS. The needles were collected in plants in four orthogonal points (South, North, East and West), sampled new needles, mature needles and old needles. The material was dried in a stove, milled and chemically analyzed (macro and micronutrients). The concentrations of N, P, K, B, Cu and Zn had decreased, of Ca, Fe and Mn increased, and the Mg and S have remained constant with the age of the needles. The retranslocation rate (old-new needles) was more than 50% for most nutrients, except for Mn and Fe, showed that cumulative effect and the Ca reference element. Keywords: nutrients redistribution; biochemical cycle; phenological stages. INTRODUÇÃO Após a promulgação da lei dos incentivos fiscais pelo Governo, nos anos 60, grandes áreas foram ocupadas com o plantio de florestas. Dentre as espécies mais plantadas, especialmente no Rio Grande do Sul, encontram-se o Pinus taeda L. e Pinus elliottii Engelm. var. elliottii, ocupando atualmente uma área de aproximadamente 182.000 hectares (ABRAF, 2007). Esses plantios em geral foram introduzidos em locais com solo de baixa fertilidade, mas mesmo assim, apesar da produção ser relativamente baixa, as plantas demonstraram uma capacidade extraordinária de adaptação com um bom uso dos escassos recursos nutricionais. A utilização de nutrientes está intimamente relacionado com a ciclagem de nutrientes, através do movimento de nutrientes dentro da planta (ciclagem interna) e desta com o solo por meio da queda de tecido foliar (SILVA et al., 1998). Dessa forma, relativa quantidade de nutrientes retorna ao solo por meio da decomposição dos tecidos senescentes, ou é retranslocada para outros tecidos das plantas pelo ciclo bioquímico, o que explica a manutenção da fertilidade do solo sob floresta por longos períodos (MENEZES JÚNIOR et al., 2008), e a manutenção de teores adequados de nutrientes no tecido vegetal para o suprimento requerido por uma determinada espécie. O ciclo bioquímico envolve a retranslocação de um determinado elemento de um local de residência (órgão) para outro, como por exemplo, de uma folha velha para uma nova (MALAVOLTA, 1997), ou seja, refere-se à transferência de nutrientes no próprio interior da planta. A retranslocação de nutrientes durante a senescência foliar é um processo fisiológico importante para a retenção dos nutrientes na planta para suas possíveis necessidades (AERTS, 1996). Segundo Piatek & Allen (2000), as fontes primárias de nutrientes nos tecidos foliares são oriundos da oferta do solo e da retranslocação de tecidos senescentes. A taxa de crescimento das árvores, mais do que a disponibilidade de nutrientes no solo, é o principal fator controlador da retranslocação e que, em espécies de pinus, a retranslocação dos nutrientes dentro da planta constitui-se numa fonte de suprimento importante (NAMBIER e FIFE, 1987). Em estudo realizado por Finér (1994), com Pinus silvestrys, ele observou que as concentrações de K e Mg foram maiores nas acículas novas, o Ca, Fe e Mn, apresentaram teores mais elevados nas acículas mais antigas, já as concentrações de N, P e Zn não variaram com a idade das acículas. Mas cabe ressaltar que, geralmente a concentração de nutrientes nos tecidos foliares está associada às características biológicas da espécie (NARDOTO et al., 2006) e pelas condições edáficas e climáticas do sítio florestal (HAAG, 1985), e em solos de baixa fertilidade onde a demanda por nutrientes na planta é elevada, a ciclagem interna é bem mais acentuada (CALDEIRA et al., 1999). O presente estudo objetiva avaliar a concentração e a retranslocação de macronutrientes e micronutrientes das acículas jovens para as senescentes em plantas de Pinus taeda L. MATERIAL E MÉTODO Caracterização das áreas de estudo O estudo foi realizado em dois povoamentos distintos de Pinus taeda L., um em área pertencente à empresa Reflorestadores Unidos (povoamento 1) e outro pertencente à empresa Cambará S.A. (povoamento 2), ambos localizados no município de Cambará do Sul, RS. O povoamento 1 está situado nas coordenadas geográficas de 29°10’de latitude Sul e 50°19’ de longitude Oeste, numa altitude média de 893 metros e o povoamento 2 nas coordenadas geográficas de 28° 52’ de latitude Sul e 50° 08’ de longitude Oeste, numa altitude média de 1089 m. O município de Cambará do Sul localiza-se no nordeste do estado do Rio Grande do Sul, na região fisiográfica dos Campos de Cima da Serra (Figura 1). Segundo a classificação climática de Köppen, o tipo de clima fundamental predominante é o Cfb (temperado úmido), a precipitação média na região é de 1.700 mm, bem distribuída durante o ano. A temperatura média anual é de aproximadamente 15°C, sendo que a média das temperaturas máximas é de 22ºC e a média das temperaturas mínimas é de 8,5ºC (MORENO, 1961). Conforme Motta et al. (1971), os ventos dominantes na região são alísios, ou seja, sopram do mar para a terra, em consequência da região situar-se próximo ao litoral norte do estado do Rio Grande do Sul. Quanto ao solo, o material de origem é rocha basáltica, resultante do derrame basáltico do Triássico Superior, predominando solos rasos com horizonte A de coloração escura, com baixa saturação de bases e teor elevado de alumínio trocável, portanto extremamente ácidos. O solo da região pertence à Unidade de Mapeamento Bom Jesus, sendo classificado como Cambissolo Húmico alumínico típico (STRECK et al., 2008). O povoamento 1 foi implantado em área de campo nativo, após preparo mínimo de solo com escarificação na linha de plantio, atingindo uma profundidade média de 30 cm. O plantio do povoamento decorreu-se no período de inverno do ano de 1999, com mudas clonais produzidas em tubetes e optando-se por um espaçamento de 3 m x 2 m. Por ocasião do plantio não foi realizado nenhum tipo de adubação. Não se efetuou manutenções no plantio (controle de matocompetição e de pragas). Aos 4 anos de idade as árvores sofreram poda (desrama) até 2,5 metros de altura. As características químicas do solo sob o povoamento 1 podem ser observadas pela Tabela 1. No entanto, o povoamento 2, foi implantado em área de corte raso, onde os resíduos do primeiro povoamento (Pinus taeda L.) foram enleirados. Para o plantio, utilizou-se coroamento (60 cm x 60 cm) como preparo mínimo de solo. O plantio do povoamento ocorreu no período de inverno de 1999, com mudas clonais produzidas em tubetes, e em espaçamento de 3 m x 2 m. Aos 5 anos de idade, todas as árvores sofreram poda (desrama) até 3,0 metros de altura. As características químicas do solo sob o povoamento 2 podem ser verificadas na Tabela 2. Metodologia de estudo Em condições de sítio semelhantes, foram alocadas quatro parcelas no povoamento 1 e três no povoamento 2, de 24 m x 20 m, nelas foram escolhidas aleatoriamente dez árvores por parcela, as quais apresentavam boas condições de sanidade (sem ataque aparente de pragas e doenças), danos mecânicos e bifurcações. Em cada uma das árvores avaliadas, foram coletados ramos do terço médio da copa, procedimento descrito por Bellote e Silva (2000), nos quatro pontos ortogonais (sul, norte, leste e oeste), neles, amostraram-se as acículas jovens, maduras e senescentes. Após a amostragem, misturou-se o material das dez árvores de cada parcela, formando uma amostra composta para cada estádio fenológico das acículas em cada parcela considerada. Os estádios fenológicos das acículas foram definidos com base na posição destas nos ramos. Foram denominadas de acículas jovens aquelas dispostas na parte superior (terminal ou lateral) dos ramos, com 2 a 3 meses de idade (crescimento na primavera). Por outro lado, as acículas maduras com aproximadamente um ano de idade se encontravam na parte mediana dos ramos (crescimento no período primavera-verão do ano anterior) e as senescentes das acículas recém-caídas. As acículas senescentes foram coletadas com auxílio de coletores instalados sob a projeção da copa das plantas, amostrando-se o material decíduo, depois de decorrido um período de 10 dias da instalação dos coletores. As coletas foram realizadas na primeira quinzena de janeiro de 2007, em ambos os povoamentos. O material amostrado foi acondicionado em embalagem plástica, devidamente identificado e enviado ao Laboratório de Ecologia Florestal, pertencente ao Departamento de Ciências Florestais da Universidade Federal de Santa Maria. No laboratório, as amostras foram secas em estufa de circulação e renovação de ar a 70ºC, por um período de 72 horas, sendo posteriormente moídas em moinho do tipo Wiley com peneira de 30 mesh. Nas amostras, foram determinados os teores de nitrogênio pelo método Kjeldahl, fósforo e boro por espectrometria visível, potássio por fotometria de chama, enxofre por turbidimetria e o cálcio, magnésio, cobre, ferro, manganês e zinco por espectrofotometria de absorção atômica, seguindo a metodologia descrita por Tedesco et al. (1995). A retranslocação de nutrientes nas acículas foi estimada segundo a metodologia de Vitousek e Sanford (1986), a qual leva em conta a retranslocação de Ca, elemento praticamente imóvel, que pode servir de referência. Para este estudo, utilizou-se a fórmula adaptada por Chuyong et al. (2000), como segue abaixo: %NutRe = {1 – [(Nut./Ca(senescente))/(Nut./Ca(verde))]} x 100 Em que: %NutRe = porcentagem do nutriente retranslocado; Nut./Ca(senescente) = relação entre os teores foliares dos nutrientes em avaliação e o teor de Ca nas acículas senescente; Nut./Ca(verde) = relação entre os teores foliares dos nutrientes em avaliação e o teor de Ca, nas acículas verde. Para a análise estatística, o delineamento experimental adotado foi o inteiramente casualizado com três tratamentos e quatro repetições para o povoamento 1(campo nativo) e três repetições para o povoamento 2 (segunda rotação). Os tratamentos constituíram-se das acículas coletadas, divididas em jovem, madura e senescente. As análises estatísticas foram realizadas com o auxílio do programa estatístico Genes (CRUZ, 2001) ao nível de 5% de probabilidade de erro, para a separação dos contrastes de médias utilizou-se o teste de Tukey. RESULTADOS E DISCUSSÃO Com base na Tabela 3, pode-se verificar a concentração e a taxa de retranslocação de macronutrientes nas acículas em plantas de Pinus taeda implantadas em área de campo nativo e em segunda rotação. Dentre todos os nutrientes, o Ca e o N foram os de maior teor nas acículas maduras e senescentes, já nas acículas novas o K e novamente o N foram superiores, em ambos os povoamentos. O alto teor de Ca, nas acículas maduras e senescentes, pode ser decorrente, de uma liberação mais lenta desse elemento e pela baixa taxa de retranslocação (FIFE e NAMBIAR, 1991; PADRO, 2008), sendo considerado um elemento imóvel na planta, fazendo parte da lamela média da parede celular (MALAVOLTA et al., 1997), ficando armazenado em forma de cristais na folha, permanecendo nela mesmo em sua senescência (NEVES et al., 2001; DIAS et al., 2002), como verificado neste estudo, no qual o teor nas acículas novas foi de 2,29 e 2,17 g kg-1 e nas senescente de 5,82 e 5,12 g kg-1 respectivamente para os povoamentos em área de campo nativo e em segunda rotação. Os teores de macronutrientes nas acículas maduras foram inferiores aos valores considerados adequados para a cultura do pinus (Malavolta et al., 1997), exceto para o cálcio. Mas cabe ressaltar que os teores de nutrientes nos tecidos vegetais podem variar dentro da mesma espécie, dependendo da idade das plantas, época de coleta, posição de amostragem, tipo de solo, fatores climáticos, dentre outros (HAAG, 1985; NAMBIAR e FIFE, 1991; AERTS, 1996; CALDEIRA et al., 1999; ALBAUGH et al., 2000; PIATEK e ALLEN, 2000; NARDOTO et al., 2006; NAMBIAR e SAUR, 2008). Segundo Borém e Ramos (2002), para o Ca, Mg, K e P, a quantidade contida na serapilheira (material senescente) está diretamente relacionada à quantidade disponível desse elemento no solo, nos primeiros 10 centímetros. Sendo que, no caso do K e do Mg, eles podem sofrer lixiviação através da água da chuva que atravessa o dossel e permeia a serapilheira, o que poderia explicar os teores baixos desses elementos nesta. A ordem relativa da concentração de macronutrientes nas acículas foi de: N > Ca > K > MG > S = P, segundo Haag (1985) essa tendência é encontrada na maioria dos estudos com diferentes formações florestais. Os macronutrientes mais móveis (P e K) apresentaram-se estatisticamente com teores superiores (p < 0,05) nas acículas jovens, seguido pelas acículas maduras e senescentes. Comportamento inverso e significativo foi verificado para o Ca, que possui baixa capacidade de redistribuição, em que ocorreu um aumento por causa do efeito de concentração. As acículas novas e maduras apresentaram teores de N similares, diferindo significativamente das acículas senescentes. Já os teores de Mg e S mantiveram-se estatisticamente iguais (p > 0,05) em todos os estádios fenológicos sem apresentar um padrão de variação definido. Zhang e Allen (1996) observaram, num povoamento de Pinus taeda, que o aumento no conteúdo de nutrientes nas acículas novas foi associado com a diminuição do teor de nutrientes das acículas velhas, sugerindo que ocorra a transferência de nutrientes para o desenvolvimento das folhas. Esse mesmo autor infere que a amostragem de acículas para a avaliação da deficiência de nutrientes, as amostras foliares devem ser coletadas no terço médio da copa em acículas de um ano de idade durante o período de rápido crescimento de novos brotos. Em relação à taxa de retranslocação, estimou-se que 83,5 e 74,9% do N; 89,5 e 86,1% do P; 96,3 e 91,0% do K; 57,2 e 58,0% do Mg e 52,6 e 66,2% do S foram redistribuídos das acículas senescentes para acículas novas respectivamente para o povoamento em área de campo nativo e em segunda rotação. Dessa forma, a magnitude de retranslocação foi de: K > P > N > MG = S, sendo essa sequência igual à estimada por Albaugh et al. (2008) e Zhang e Allen (1996) para plantas de Pinus taeda com 8 e 11 anos de idade respectivamente e similar a citada por Landsberg (1986) em sua revisão para as mais diversas formações florestais. Segundo Albaugh et al. (2008) essas elevadas taxas de retranslocação de nutrientes móveis indica a importância desse recurso para o crescimento e sobrevivência das plantas de Pinus taeda. Teores elevados de Fe e Mn (Tabela 4) podem ser decorrentes das altas concentrações de tais elementos no solo (Tabela 1 e 2) segundo o Manual de Adubação e Calagem-RS/SC (2004). A magnitude de concentração dos micronutrientes foi a mesma nos três estádios fenológicos (Mn > Fe > Zn > B > Cu). Os teores de micronutirentes nas acículas maduras foram inferiores aos valores considerados adequados para a cultura do pinus (Malavolta et al., 1997), exceto para o ferro e manganês, sendo que o último possui concentração bem superior ao intervalo considerado adequado (200-300 mg kg-1). O Cu, B (em ambos os povoamentos) e o Zn (em campo nativo) mostraram uma diminuição significativa (p < 0,05) das concentrações em acículas senescentes, indicando uma possível mobilidade desses micronutrientes no tecido foliar. Segundo Helmisaari (1990), Lehto e Mälkönen (1994), Brown e Shelp (1997), Lehto et al. (2000), Lehto et al. (2004) e Konsaeng et al. (2005) a mobilidade do boro está restrita às espécies que produzem alcoóis de açúcar (polióis) que se ligam com o boro (poliol-B), formando complexos com maior estabilidade, tornando-o prontamente translocavel. Por isso, o B é considerado o único nutriente essencial que é móvel em algumas espécies e imóvel em outras. Lehto et al. (2000) e Lehto et al. (2004) relatam que em algumas espécies de coníferas (Pinus sylvestris e Picea abies) o B é retranslocado pelo floema. Já para a espécie do presente estudo (Pinus taeda) não se verificou na literatura indicativos da mobilidade ou não do boro, nesse caso Perica et al. (2001) salienta a importância da verificação da presença de polióis nas plantas para a indicação da mobilidade do boro. Já o Mn e o Fe apresentaram comportamento similar ao Ca, apresentando valores estatisticamente superiores (p < 0,05) nas acículas senescente e maduras em relação às acículas novas, demonstrado baixa capacidade de redistribuição. A taxa de retranslocação de micronutrientes foi de 73,3 e 72,0% do B; 90,5 e 85,6% do Cu; -83,9 e -97,9% do Fe; -77,2 e -41,9% do Mn e 69,7 e 59,2 do Zn foi redistribuído das acículas senescente para acículas novas, respectivamente para o povoamento em área de campo nativo e em segunda rotação. Com uma magnitude de retranslocação de: Cu > B > Zn > Mn > Fe. Cabe ressaltar, que para o Mn e Fe, suas taxas negativas referem-se ao seu efeito acumulativo nas acículas senescentes em relação às novas. A dinâmica de nutrientes nas árvores varia em função da espécie, da idade, condições edafoclimáticas do sítio e práticas de manejo adotadas. Assim, observa-se que a concentração de alguns elementos nas folhas aumenta, enquanto as de outros decresce. Há, dessa maneira, uma translocação de nutrientes de órgãos senescentes para regiões de crescimento da árvore. Todos esses aspectos evidenciam a grande importância da ciclagem interna dos nutrientes para a manutenção do balanço nutricional das plantas (HAAG, 1985; NAMBIAR e FIFE, 1991; AERTS, 1996; COLIN-BELGRAND et al., 1996; SILVA et al., 1998; CALDEIRA et al., 1999; ALBAUGH et al., 2000; PIATEK e ALLEN, 2000; NARDOTO et al., 2006; NAMBIAR e SAUR, 2008 ). CONCLUSÕES As concentrações de N, P, K, B, Cu e Zn diminuíram significativamente (p < 0,05) com o aumento da idade das acículas, já o Ca, Fe e Mn aumentaram significativamente (p < 0,05) e o Mg e S mantiveram-se constantes; A taxa de retranslocação (acícula senescente-nova) foi superior a 50% para a maioria dos nutrientes, com exceção ao Mn e Fe, que apresentaram efeito acumulativo. AGRADECIMENTOS Os autores agradecem às empresas Cambará S/A e Reflorestadores Unidos S/A, por cederem área particular para desenvolver o presente estudo e pelo apoio logístico-financeiro. REFERÊNCIAS BIBLIOGRÁFICAS
Copyright © 2009 - Ciência Florestal The following images related to this document are available:Photo images[cf09036t4.jpg] [cf09036f1.jpg] [cf09036t3.jpg] [cf09036t1.jpg] [cf09036t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}