 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ciência Florestal, Vol. 20, No. 1, Jan-Mar, 2010, pp. 147-156 Tolerância de mudas de canafístula (Peltophorum dubium (Spreng.) Taub.) inoculada com Pisolithus microcarpus solo com excesso de cobre1 Tolerance of canafístula (Peltophorum dubium (Spreng.) Taub.) seedlings inocculated with Pisolithus microcarpus to copper contaminated soil Rodrigo Ferreira da Silva2 Zaida Inês Antoniolli3 Manoeli Lupatini4 Lineu Leal Trindade4 Alessandro Salles da Silva5
Recebido para publicação em 5/12/2007 e aceito em 11/12/2009. Code Number: cf10012 RESUMO A associa ção de fungos ectomicorrízicos com essências florestais nativas pode ser uma alternativa para revegetação de solos contaminados por metais pesados. O objetivo do trabalho foi determinar o comportamento de mudas de canafístula inoculadas com fungo ectomicorrízico em solo contaminado por cobre. O fungo ectomicorrízico utilizado foi o Pisolithus microcarpus UFSC Pt116, produzido em substrato composto por turfa-vermiculita (3:1). As mudas de canafístula foram desenvolvidas em areia lavada e transplantada para unidades experimentais quando apresentaram duas folhas definitivas. Como unidade experimental foi utilizado vaso com capacidade de 1 L contendo 1 kg de solo. A inoculação foi realizada no momento do transplante das mudas, sendo adicionado 2 g de inoculante por unidade experimental. O delineamento experimental foi inteiramente casualizado, num esquema bifatorial (2 x 4) qualitativo em A, com e sem inoculante do fungo, e quantitativo em D, sendo o controle, adição de 150 mg kg-1, 300 mg kg-1 e 450 mg kg-1 de cobre (CuSO4), com sete repetições. Determinou-se altura de planta, diâmetro do colo, massa seca da parte aérea, teor de cobre na planta, porcentagem de colonização micorrízica, área superficial específica radicular e comprimento radicular. O incremento das doses de cobre no solo reduziu o comprimento radicular, área superficial específica radicular, altura e diâmetro das plantas de canafístula. As mudas de canafístula inoculadas com fungo ectomicorrízico apresentam menor teor de cobre em seu tecido e reduzem a absorção de cobre na dose mais elevada. Palavras-chave: ectomicorriza; metal pesado; contaminação do solo. ABSTRACT The ectomycorrhizal fungi symbiosis with native florestal essences may be an alternative to revegetation of metal contaminated soils. The aim of this work was to determine canaf ístula seedlings behavior, inoculated to ectomycorrhizal fungi in soil with copper excess. The trial was conducted in greenhouse conditions. The ectomycorrhizal fungi used was Pisolithus microcarpus UFSC Pt116, cultivated on peat-vermiculite substrate (3:1). The canafístula seedlings were developed in washed sand and transplanted, when showed two definitive leaves. Vase of 1L with 1 kg of soil were used as experimental unit. The inoculation was accomplished in the seedlings transplant, added 2g of inoculates for experimental unit. The experimental design was entirely casual in bi factorial scheme (2 x 4) qualitative in A, with and without fungi inoculum and quantitative in D being the control addition of 150 mg Kg -1, 300 mg Kg -1 and 450 mg Kg -1 of copper (CuSO4), with seven repetitions. Plant height, colon diameter, dry matter, copper content, mycorrhizal colonization percentage, superficial roots area and roots length were assessed. The increase of soil copper levels reduced roots length, specific superficial roots area, height and plant diameter of canafístula. Canafistula seedlings inoculated to ectomycorrhizal fungi showed low cooper amounts in their tissue and copper absortion reduced in high copper doses. Keywords: ectomycorrhizal; heavy metal; contamination soil. INTRODUÇÃO A intensa demanda por produtos manufaturados tem contribu ído para contaminação do ambiente por metais potencialmente tóxicos, como zinco (Zn), cobre (Cu) e chumbo (Pb) (RIBEIRO FILHO et al., 1999). Esses metais podem contaminar os ecossistemas de forma irreversível (LEYVAL et al., 1997), causando redução da atividade microbiana do solo (DIAS JÚNIOR et al., 1998) e alterando a composição das espécies de fungos (NORDGREN et al., 1985; RÜHLING e SÖDERSTROM, 1990). Desse modo, quando uma área atinge níveis de contaminação muito elevados poderá ocorrer supressão da vegetação. Nesse sentido, a seleção de organismos tolerantes a esse tipo de ambiente, torna-se necessário na busca de alternativas para a revegetação dessas áreas. Embora o cobre seja um elemento essencial às plantas (SOLIOZ e STOYANOV, 2003), áreas contaminadas por esse elemento têm se tornado comum na Região Sul do Brasil, em consequência das atividades de mineração e em decorrência da aplicação de fungicidas à base de cobre em regiões vinícolas, (GIOVANNINI, 1997). A dificuldade no estabelecimento e desenvolvimento da vegetação nessas áreas implica no estudo de alternativas que envolvam plantas e organismos do solo que apresentem tolerância ao contaminante. As essências florestais nativas podem ser uma opção para revegetação de áreas contaminadas por cobre. Essas plantas, além de suas características fisiológicas desejáveis, como crescimento relativamente rápido, vigor e rusticidade, apresentam também grandes vantagens comerciais quanto à madeira, e subprodutos em relação às comercialmente mais utilizadas como o eucalipto e o pinus (SCHROEDER, 1991). A canafístula (Peltophorum dubium (SPRENG.) TAUB.) é uma leguminosa de ampla e expressiva dispersão, freqüente nas matas da bacia do rio Paraná, indicada para recuperar áreas degradadas (BACKES e IRGANG, 2004). No Rio Grande do Sul, embora as espécies florestais nativas tenham sido muito estudadas nos últimos anos, sobretudo em solos em processo de degradação, poucos estudos têm sido realizados com ênfase na ecologia e associações com outros organismos, como os fungos ectomicorrízicos, os quais podem favorecer o estabelecimento dessas plantas. Os fungos ectomicorrízicos, ao associar-se com as plantas, desenvolvem estruturas eficientes na absorção de água e nutrientes do solo, os quais são posteriormente transferidos para as plantas (SMITH e READ, 1997; SILVA, 2002; SILVA et al., 2003). Esses fungos se destacam por interagir com as espécies vegetais formando uma associação simbiótica mutualística, que através da maior absorção de nutrientes e proteção das raízes, contribuem para o desenvolvimento das plantas (PEROTTO e BONFANTE, 1997). Esses fungos conferem às plantas maior tolerância aos metais pesados, indicando a possibilidade de uso dos mesmos em processos de biorremediação de solos poluídos por meio do acúmulo desses elementos nas plantas (LEYVAL et al., 1997; KHAN, 2001; SCHELOSKE et al., 2001). Os fungos ectomicorrízicos são benéficos ao crescimento da planta hospedeira, especialmente em situações em que fatores climáticos e edáficos são limitantes (SMITH e READ, 1997). Sabe-se que certos fungos podem, ainda, apresentar elevada tolerância a metais pesados (MEDVE e SAYRE, 1994). Esse efeito tem sido atribuído à habilidade dessas associações em reter os metais no micélio fúngico, evitando a translocação destes para a parte aérea da planta, aumentando sua tolerância (COLPAERT e VAN ASSCHE, 1993; AGGANGAN et al., 1998). Moreira e Siqueira (2002) atribuem essa habilidade a mecanismos que incluem processos externos às hifas, ligação a polímeros da parede celular e processos internos nas células dos fungos, em que os metais podem ser complexados, compartimentalizados ou volatilizados. Contudo, quando em excesso, os metais pesados podem influenciar negativamente os fungos ectomicorrízicos (DIXON e BUSCHENA, 1988), podendo inibir a colonização em plantas (BELL et al., 1988; GRAZZIOTTI, 1999). Rühling e Söderstrom (1990) observaram redução no número de frutificações e espécies de Basidiomicetos à medida que aumentava o nível de contaminação por As, Cd, Cu, Pb e Zn. Também é observado que os fungos ectomicorrízicos respondem de forma diferenciada a contaminação do solo, de acordo com a espécie e o tipo de metal envolvido (HARTLEY et al.,1997). Por isso, o estabelecimento de essências florestais nativas micorrizadas pode ser uma alternativa na revegetação e aproveitamento de áreas degradadas contaminadas por cobre. Nesse sentido, este trabalho teve como objetivo avaliar o comportamento de mudas de canafístula inoculadas com o fungo ectomicorrízico Pisolithus microcarpus UFSC Pt116 em solo com excesso de cobre. MATERIAL E MÉTODOS Solo O experimento foi desenvolvido em casa de vegeta ção da Estação de Pesquisas Florestais de Santa Maria – FEPAGRO-Florestas, localizada no distrito de Boca do Monte, Santa Maria, RS e teve duração de 180 dias. O solo utilizado no experimento é classificado como Argiloso Vermelho-amarelo distrófico típico (EMBRAPA, 1999). As amostras de solo, após coleta com trado calador, a 10 cm de profundidade, foram secas, destorroadas, passadas em peneira de 2 mm e em seguida esterilizadas em autoclave. O processo de esterilização constou em colocar o solo em sacos de pano e submeter à temperatura de 121ºC por 1 hora, sendo essa operação repetida por três vezes consecutiva. A caracterização da amostra utilizada é apresentada na Tabela 1. Espécie florestal A planta utilizada no experimento foi a canafístula (Peltophorum dubium (SPRENG.) TAUB). As mudas foram produzidas em areia lavada e esterilizada em autoclave. Posteriormente, foram transplantadas para os vasos de cultivo quando apresentavam dois pares de folhas definitivas. Não foi utilizada adubação de P e K, pois o solo apresentava teores satisfatórios desses nutrientes. Aplicou-se o equivalente a 30 kg ha-1 de N na forma de uréia como adubação de base e calagem com calcário dolomítico para elevar o pH do solo para 5,5. Os vasos de cultivo apresentavam capacidade de 1 L, nos quais foi colocado 1 kg de amostra de solo. Cada vaso foi considerado uma unidade experimental. A umidade do solo foi mantida a 80% da sua capacidade de campo, completando a diferença do peso dos vasos, com água destilada até 1,2 kg (com base no peso dos vasos inicialmente saturados). Inoculante ectomicorrízico O inoculante utilizado no experimento foi produzido tomando por base o fungo ectomicorrízico Pisolithus microcarpus UFSC Pt116. O fungo foi cultivado em 50 mL de meio líquido MNM (MARX, 1969) em erlenmeyer de 250 mL. Quando o fungo apresentou crescimento cobrindo toda superfície do erlenmeyer foi transferido para substrato, previamente esterilizado, contendo turfa-vermiculita (3:1), acrescido de 200 mL de meio MNM líquido. Após 40 dias o fungo apresentou crescimento por todo o substrato, foi então homogeneizado e aplicado 2 g dessa mistura em cada tratamento com fungo. Aplicou-se também 2 g do substrato turfa-vermiculita sem fungo nos tratamentos sem o inoculante. Delineamento experimental O delineamento experimental foi inteiramente casualizado num bifatorial (2 x 4) qualitativo em A (com e sem fungo ectomicorrízico) e quantitativo em D (controle, adição de 150 mg kg-1, 300 mg kg-1 e 450 mg kg-1), com sete repetições, totalizando 56 unidades experimentais. Parâmetros avaliados Avaliaram-se os seguintes parâmetros: altura de planta, diâmetro de colo, massa seca da parte aérea, comprimento radicular, área superficial específica radicular, conforme Tennant (1975) teor de cobre na planta (SILVA, 1999) e porcentagem de colonização radicular (GIOVANNETTI e MOSSE, 1980). Altura de planta e diâmetro do colo A altura de planta foi medida utilizando-se uma régua graduada de 50 cm de comprimento. Para evitar a interferência da irregularidade da superfície do solo contido nos potes, colocou-se uma pequena lâmina de isopor no ponto zero da régua formando um ângulo de 90º. Para a medida do diâmetro de colo, foi utilizado um paquímetro digital, marca Mitutoyo. Massa seca da parte aérea e do sistema radicular As plantas foram cortadas rente ao solo, separando a parte aérea do sistema radicular. As raízes foram separadas do solo, lavadas com água destilada, em seguida, a parte aérea das plantas e as raízes foram colocadas em sacos de papel, identificadas e levadas à estufa a 65ºC, onde permaneceram até atingirem o peso constante. Após, pesou-se em balança digital com três casas decimais, obtendo-se a massa seca da parte aérea. A massa seca radicular foi utilizada para determinação de cobre na raiz. Comprimento radicular Na análise do comprimento radicular, separou-se uma amostra de 0,2 g de raízes cortadas em 1 cm e distribuiu-se em uma placa quadriculada de 1 cm x 1 cm e, em seguida, contou-se o número de intersecções das raízes com as linhas da placa. O comprimento radicular foi estimado seguindo-se o método de Tennant (1975), conforme a expressão: L = N° int er sec ções × Fc ; sendo L = Comprimento radicular; e Fc = Fator de correção (0,7857). Área superficial específica radicular A área superficial específica do sistema radicular foi estimada segundo Tennant (1975), conforme a expressão: S = 2π× R × L ; sendo S = Área Superficial Específica; R = Raio; e L = Comprimento Radicular.
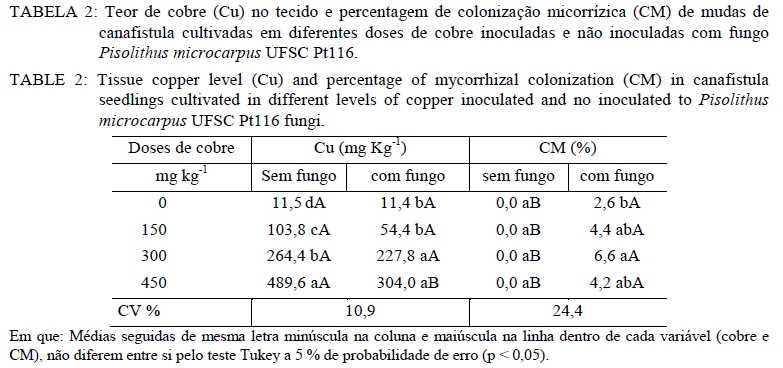
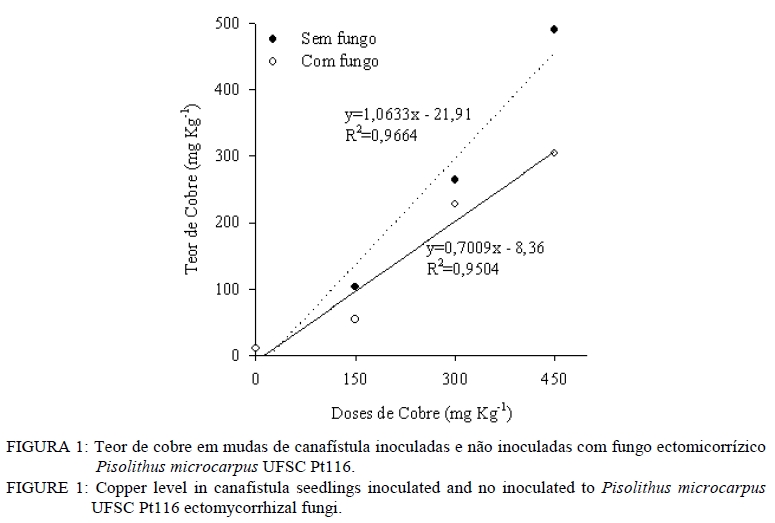
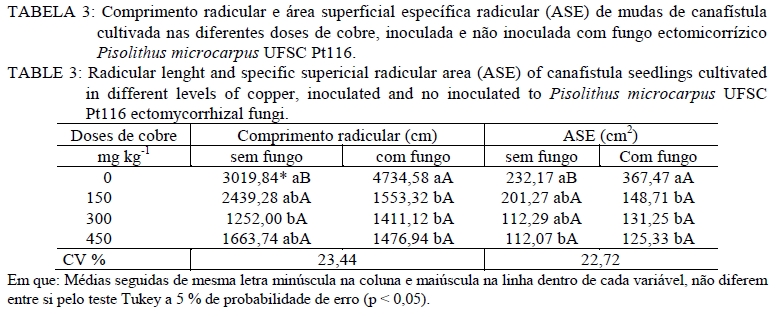
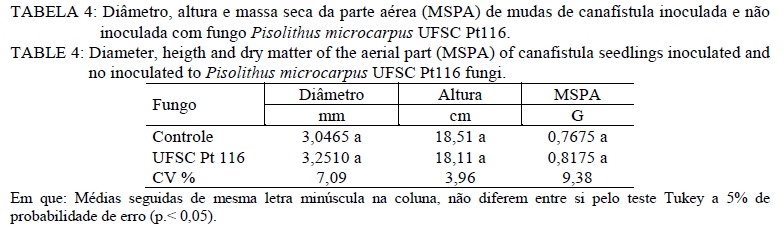
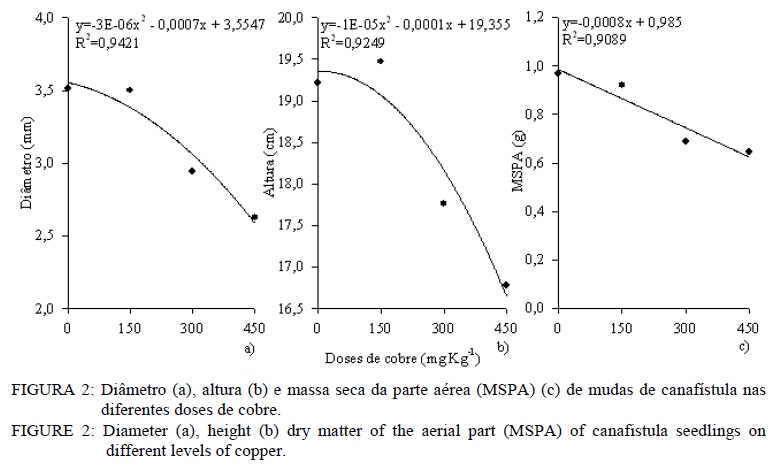
Análise química O material utilizado para essa análise foi à massa seca da parte aérea e massa seca radicular. Estas foram moídas em moinho tipo facas, passada em peneira de 2 mm e então submetida à análise química para determinar as concentrações de Cobre. Na análise de Cu empregou-se a digestão por via úmida com solução nitro-perclórica (HNO3 + HClO4) na proporção 3:1, segundo metodologia descrita por Silva (1999). As concentrações de Cu foram determinadas em espectrofotômetro de absorção atômica. Avaliação da associação micorrízica As amostras para determinação da porcentagem de raízes colonizadas por fungos ectomicorrízicos foram coletadas na ocasião da coleta do experimento. As raízes das plantas foram separadas do solo, através de peneiras e lavadas com água destilada e em seguida, retirou-se uma amostra de 0,1 g de raízes, as quais foram cortadas em 1 cm e armazenadas em solução com álcool comercial a 50%. No laboratório, essas raízes foram submetidas ao processo de clareamento e coloração. O procedimento de clareamento e coloração das raízes constou em deixar uma amostra de 0,1 g de raízes imersas em solução de KOH 10% a 80oC durante 1h30min. Após, lavou-se com água e posteriormente foram colocadas em HCl 0,1 mol L-1 durante 2 minutos. Lavou-as novamente com água e colocou-se em Trypan Blue (corante) a 80 oC por 30 minutos. Posteriormente, lavou-as novamente com água e armazenou-as em lactoglicerol, conforme Brundrett et al. (1996). A avaliação da percentagem de colonização micorrízica (CM) foi estimada pelo método da placa quadriculada (GIOVANNETTI e MOSSE, 1980), por meio da expressão: CM : (N° total de raízes colonizadas /N° total de raízes )×100 . Análise estatística Os resultados foram submetidos à análise de variância e quando da significância da análise as médias foram comparadas pelo teste de Tukey a 5% de probabilidade de erro. Equações de regressão foram ajustadas para os teores de cobre na planta, tomando-se como base os níveis de significância maiores que 95% (p < 0,05), utilizando o programa estatístico SISVAR (FERREIRA, 2000). RESULTADOS E DISCUSSÃO Os resultados obtidos mostraram que, quanto maior a dose de cobre aplicado no solo, maior é o teor de cobre no tecido das mudas de canafístula. Mesmo não apresentando diferença estatística, os teores de cobre no tecido das plantas foram menores em mudas colonizadas com o fungo ectomicorrízico (Tabela 2). Nas mudas com o fungo micorrízico, a dose 450 mg kg-1 apresentou diferença significativa em relação ao controle, indicando que a presença do fungo pode retardar a absorção de cobre pelas mudas de canafístula. (Tabela 2). O fungo micorrízico pode reter os metais em seu micélio, retardando a translocação destes para a parte aérea da planta, contribuindo assim, para o aumento da sua tolerância (AGGANGAN et al., 1998). Aos fungos ectomicorrízicos também tem sido atribuído o papel da imobilização de metais por meio de ligação a polímeros da parede celular e processos internos das células, nos quais os metais podem ser complexados, compartimentalizados ou até mesmo volatilizados (MOREIRA e SIQUEIRA, 2002). Observou-se diferença significativa entre os tratamentos com e sem fungo na colonização micorrízica e um efeito significativo das doses de cobre sob a colonização (Tabela 2). A maior colonização ocorreu com o acréscimo de cobre ao solo, sendo a dose de 300 mg kg-1 a que apresentou maior colonização, reduzindo na dose mais alta. Os metais podem expressar efeito deletério sob os fungos ectomicorrízicos, reduzindo a colonização micorrízica (GRAZZIOTTI, 1999). Nesse caso, embora o fungo UFSC Pt116 tenha reduzido a absorção de cobre pela planta, a associação pode ter sido influenciada pela toxidez deste metal. Além disso, tem se observado que o teor de fósforo no solo é tido como fator limitante para o estabelecimento da colonização micorrízica. Soares (1986) verificou alta taxa de colonização em Eucalyptus grandis Hill ex Maiden quando o substrato apresentava 4,5 mg kg-1 de fósforo extraível. Andreazza et al. (2004) observaram redução de 59% na colonização por fungos ectomicorrízicos também em Eucalyptus grandis, quando se adicionou 30 mg kg-1 de fósforo em solo arenoso com teor inicial de 8 mg kg-1 de fósforo disponível, encontrando uma média de 27% de colonização radicular com a adição do fósforo. Desse modo, a disponibilidade de fósforo no solo do experimento (12,6 mg kg-1) pode não estar favorecendo uma alta taxa de colonização micorrízica. No entanto, não há referência na literatura sobre a relação fungo ectomicorrízico e níveis de cobre em canafístula. A diferença significativa no teor de cobre no tecido em relação à presença do fungo na dose de 450 mg kg-1 e na colonização micorrízica (Tabela 2) evidencia que o fungo ectomicorrízico UFSC Pt116 pode reduzir a absorção de metais pelas plantas em doses elevadas. Os fungos ectomicorrízicos mostram habilidade em absorver metais pesados do solo podendo ser usado como atenuadores da fitotoxidez causada por esses (LEYVAL et al., 1997) e podem imobilizar os metais pesados, diminuindo a sua disponibilidade para o solo (GRAZZIOTTI et al., 2001). O manto fúngico e a rede de Harting são estruturas fúngicas que podem imobilizar os metais do solo, configurando um importante mecanismo de proteção às plantas (COSTA et al., 2003). Jones e Hutchinson (1986) observaram crescimento superior a 54%, em mudas de Betula papyrifera Marsh. colonizadas por Scleroderma flavidum Pers. na presença de 85 mol L-1 de Ni, e atribuíram esse comportamento a retenção dos metais no micélio. Os resultados do presente trabalho indicam a capacidade do fungo ectomicorrízico UFSC Pt116 em promover a retenção de cobre em seu micélio fúngico evitando que seja absorvido pelas raízes das mudas de canafístula. O teor de cobre no tecido das plantas revelou comportamento linear nas mudas de canafístula inoculadas e não inoculadas com o fungo ectomicorrízico UFSC Pt116 com adição de cobre ao solo (Figura 1). Contudo, observa-se que na presença do fungo ectomicorrízico, o teor de cobre no tecido das mudas foi menor. Trabalhos de pesquisa têm demonstrado estreita relação entre a produção de pigmentos extracelulares por fungos e a bioadsorção de metais (GADD, 1993; FOGARTY e TOBIN, 1996). A melanina, por exemplo, contém grupos carboxílicos, fenólicos, hidróxidos e aminas, os quais fornecem sítios potenciais de bioadsorção, inclusive para o cobre (GADD e DE ROME, 1988). Desse modo, o fungo ectomicorrízico UFSC Pt116 além de reter metais no seu micélio, pode também estar produzindo pigmentos extracelulares capazes de adsorver o cobre evitando que seja absorvido pelo sistema radicular das mudas de canafístula. Observou-se decréscimo no comprimento radicular e área superficial específica com adição das doses de cobre no solo (Tabela 3). O excesso de cobre pode exercer efeitos deletérios sobre a estrutura e funções das raízes (JENSEN e ADALSTEINSSON, 1989; ADALSTEINSSON et al., 1997), reduzindo o crescimento ou causando necrose nas raízes absorvente (SOARES et al., 2000). O fungo ectomicorrízico foi benéfico para o comprimento radicular e área superficial específica radicular das mudas de canafístula apenas na dose 0 mgL-1 de Cu, não exercendo efeito nas demais doses. Embora os fungos ectomicorrízicos também possam alterar o sistema radicular, proporcionando redução da dominância apical (SMITH e READ, 1997), estima-se que a superfície da área explorada por ectomicorrízas é mil vezes superior àquela de raízes sem a presença dos fungos ectomicorrízicos (HARLEY, 1969). Assim, mesmo reduzindo os parâmetros radiculares, espera-se maior capacidade de absorção de nutrientes e água pelas plantas micorrizadas, cultivadas em solo com excesso de cobre. Na análise estatística para diâmetro de caule, altura de planta e massa seca da parte aérea observouse que não houve interação significativa entre o fungo testado e as doses de cobre (Tabela 4). Contudo, espera-se que as plantas possam apresentar crescimento significativo em relação ao controle sem fungo, pois os fungos ectomicorrízicos também contribuem para aumentar a absorção de nutrientes e água pelas raízes (SMITH e READ, 1997; GRAZZIOTTI et al., 2001). Nesse caso, pode ser que as espécies florestais, como a canafístula, necessitem de maior tempo de crescimento para expressar o efeito benéfico das ectomicorrízas. Analisando apenas o efeito do cobre sobre os parâmetros sem interação, observa-se que o diâmetro e altura das mudas de canafístula apresentam a tendência de resistir ao efeito do cobre até a dose 150 mg kg-1 (Figura 2). Sabe-se que o uso de plantas pode alterar as formas dos metais pesados, reduzindo sua mobilidade e disponibilidade no solo (TORDOFF et al., 2000). As plantas podem, ainda, contribuir indiretamente na remoção de metais pelo efeito rizosférico, por meio da microbiota biodegradadora ou acumulando o metal em suas raízes (MOREIRA e SIQUEIRA, 2002). Desse modo, as mudas de canafístula podem estar apresentando certo grau de tolerância até a dose de 150 mg kg-1 de cobre. Torna-se, então, recomendável que se avalie o comportamento dessas plantas em doses intermediárias a 150 mg kg-1 de cobre, para que se obtenham informações mais conclusivas. Entretanto, observa-se que as doses testadas apresentam efeito negativo para massa seca da parte aérea das mudas de canafístula (Figura 2). Grandes concentrações de cobre causam necrose das folhas, desfolhamento precoce e diminuição do crescimento (FILHO, 2007). Tais sintomas estão diretamente relacionados com a redução das raízes, que diminui a capacidade de absorção de água e nutrientes (PANOU-FILOTHEU et al., 2001). O cobre é um elemento essencial para planta e participa do metabolismo de carboidratos, do nitrogênio, da síntese de lignina e de clorofila (MARSCHNER, 1995; FILHO, 2007). Contudo, quando em altas concentrações na solução do solo o cobre, pode proporcionar alterações nos tecidos vegetais, em nível bioquímico e fisiológico, os quais podem resultar em perdas consideráveis do potencial produtivo das plantas (FERNANDES e HENRIQUES, 1991). Quando plantas são expostas ao excesso de cobre, estas apresentam alteração na atividade da enzima nitrato redutase, reduzindo o N total na planta e o tamanho da planta por causa da redução na produção de clorofila (LUNA et al., 1997). Contudo, esse efeito não foi observado na massa seca da parte aérea das mudas de canafístula (Tabela 4) e pode ser atribuído a alterações na estrutura morfológica das plantas, que apresentavam mais ramificações, com a maior dose de cobre. CONCLUSÕES A presença de cobre no tecido vegetal de mudas de canafístula é baixa quando há associação com o fungo Pisolithus microcarpus UFSC Pt116, embora o nível de fósforo encontrado no solo pode ter influenciado negativamente a associação micorrízica. O isolado de fungo ectomicorrízico Pisolithus microcarpus UFSCPt116 reduz significativamente a absorção de cobre pelas mudas de canafístula na dose de 450 mg kg-1 . O excesso de cobre no solo reduz o desenvolvimento das mudas de canafístula. AGRADECIMENTOS Ao Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) e à Fundação de Amparo à Pesquisa do Rio Grande do Sul (FAPERGS), pelo auxílio financeiro. REFERENCIAS BIBLIOGRÁFICA
Copyright 2010 - Ciência Florestal The following images related to this document are available:Photo images[cf10012t3.jpg] [cf10012t2.jpg] [cf10012f1.jpg] [cf10012t1.jpg] [cf10012f2.jpg] [cf10012t4.jpg] |
| |||||||||

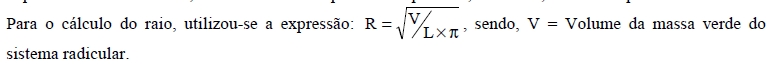
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}