 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
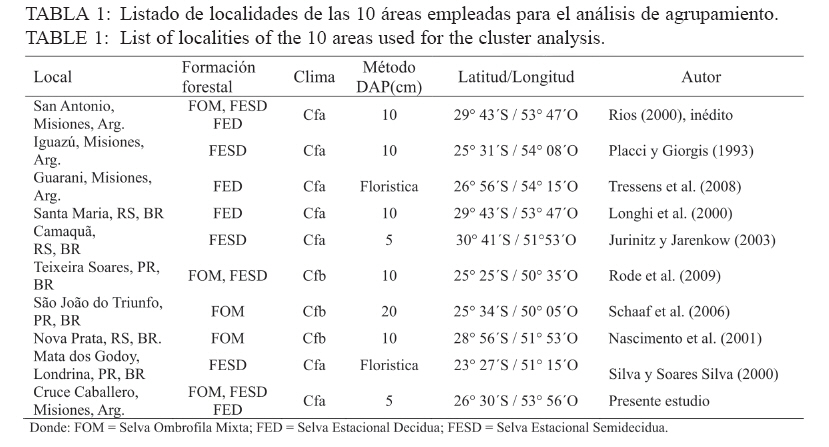
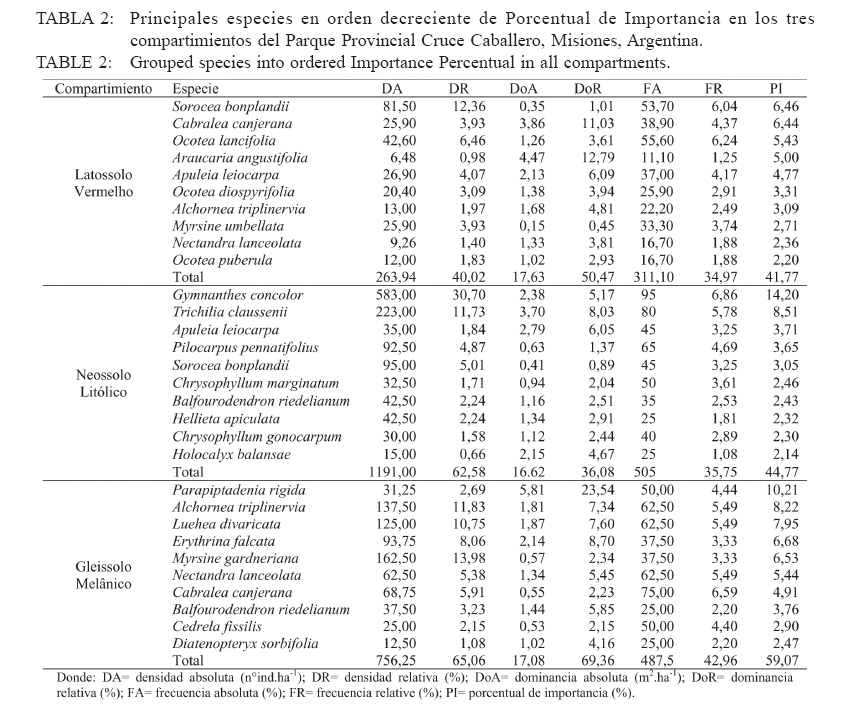
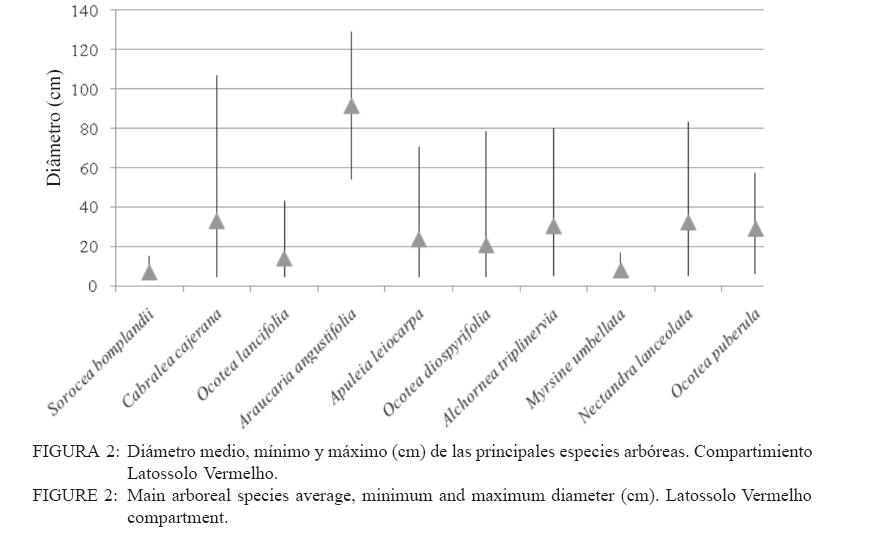
Ciência Florestal, Vol. 20, No. 2, Apr-Jun, 2010, pp. 193-206 ESTRUCTURA DE LAS PRINCIPALES ESPECIES ARBÓREAS EN EL PARQUE CRUCE CABALLERO Y SU SIMILITUD FLORISTICA CON AREAS DE ARGENTINA Y DE BRASILSTRUCTURE OF MAIN ARBOREAL SPECIES IN CRUCE CABALLERO PARK AND ITS FLORISTIC SIMILARITY WITH AREAS FROM ARGENTINA AND BRAZIL Román Carlos Ríos1 Franklin Galvão2 Gustavo Ribas Curcio3
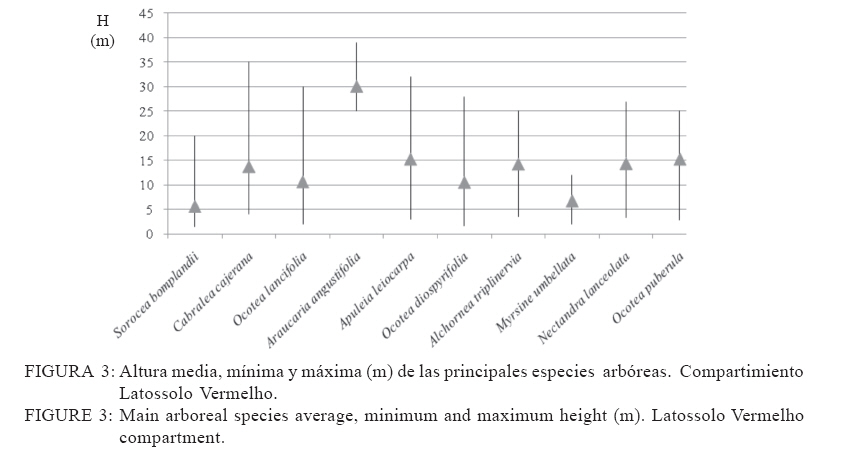
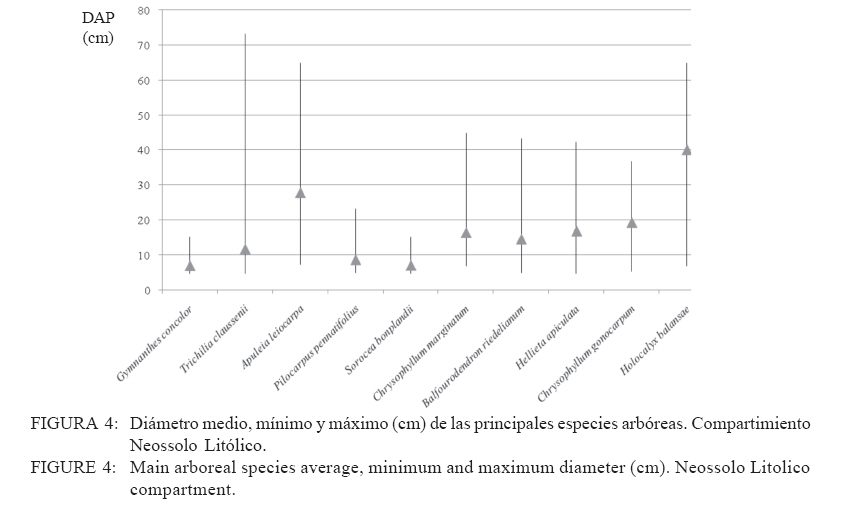
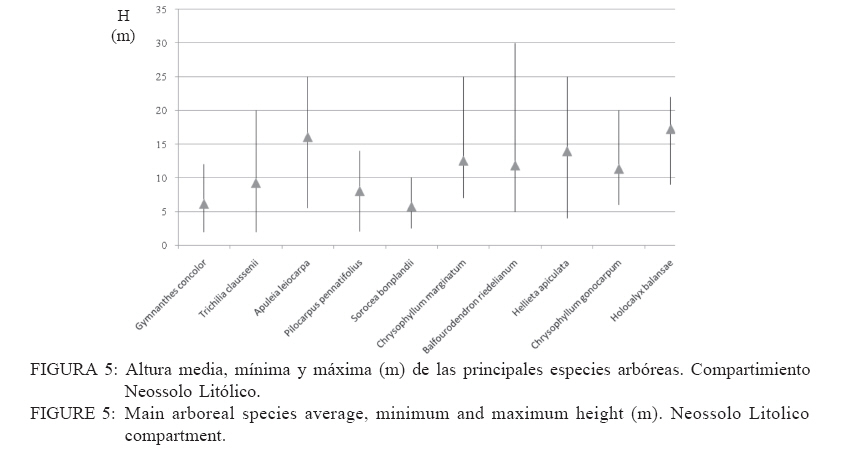
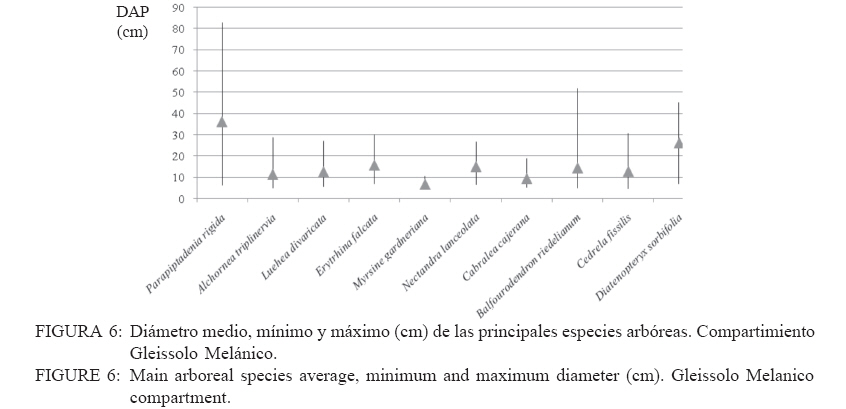
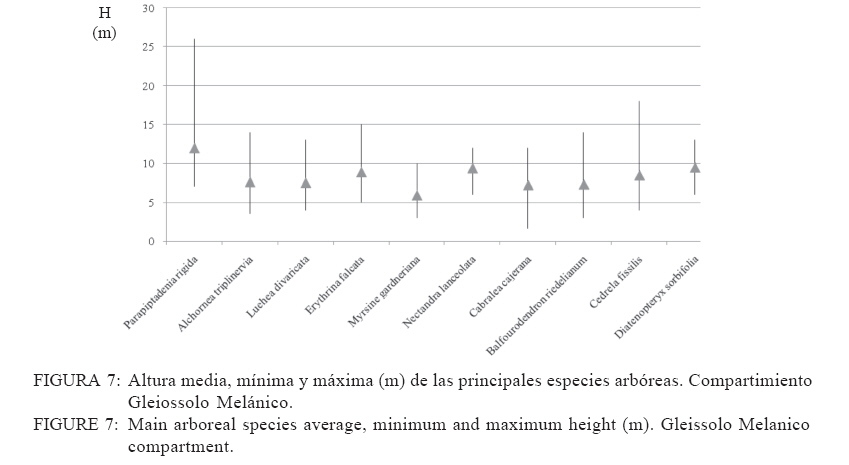
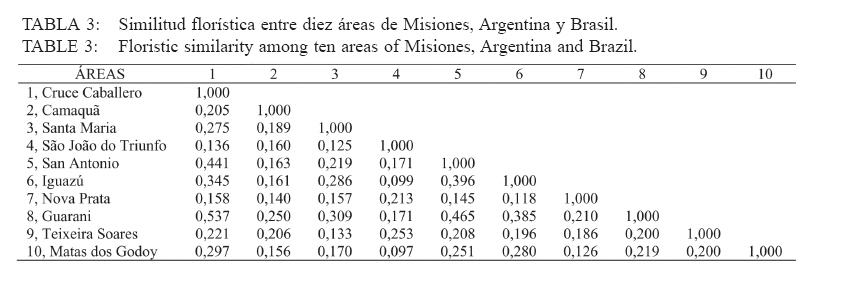
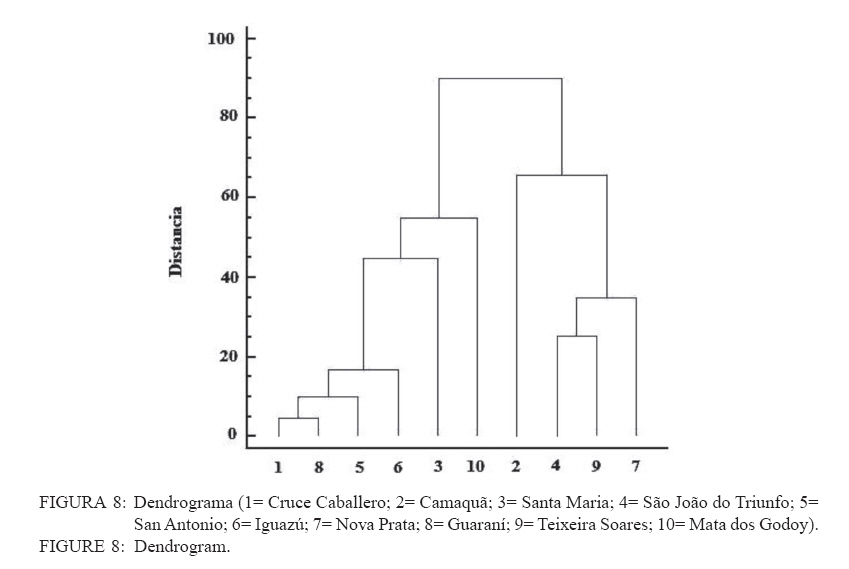
Recebido para publicação em 17/01/2008 e aceito em 05/04/2010. Code Number: cf10016 RESUMEN El presente trabajo tuvo como objetivo estudiar la estructura fitosociológica y la similitud florística entre áreas con selvas estacionales y mixtas de Misiones, Argentina y de Brasil. El Parque Cruce Caballero tiene 522 hectáreas donde se utilizó el método de las parcelas múltiples de 20 x 10 m para árboles a partir de 5 cm de diámetro a la altura de 1,30 (DAP). Se registraron e identificaron todas las especies arbóreas midiéndose el DAP y altura en tres compartimientos ambientales considerados: Latossolo vermelho, Neossolo litólico y Gleissolo Melánico. La Araucaria angustifolia (Bertol.) Kuntze domina el estrato superior en el primer compartimiento acompañada de Sorocea bonplandii (Baill.) W.C.Burger, Lanj. & Boer, Cabralea canjerana (Vell.) Mart., Ocotea lancifolia Mez y Apuleia leiocarpa J.F.Macbr.. La araucaria está ausente en el Neossolo Litólico donde predominan Gymnanthes concolor Müll.Arg., Trichilia claussenii C.DC., Apuleia leiocarpa, Pilocarpus pennatifolius Lem. y Sorocea bomplandii. La araucaria está escasamente representada en Gleissolo Melánico donde predominan Parapiptadenia rigida (Benth.) Brenan, Alchornea triplinervia Müll.Arg., Luehea divaricata Mart., Erythrina falcata Benth. y Myrsine gardneriana A.DC.. Comparando la similitud florística con otras áreas de la región se encontró que Cruce Caballero se asemeja a áreas con Selvas estacionales y presenta escasa similitud con a las selvas mixtas con araucaria de Brasil. Palavras-clave: Araucaria angustifolia; fitosociología; similitud. ABSTRACTThe present work aimed to study the fitossociologic structure and floristic similarity among seasonal and mixed forests of Argentina and Brazil. Cruce Caballero Park with 522 hectares where the method of multiple plots was adopted with (20 x 10 m). In each plot all individual tree of size equal or heigher than 5 cm diameter were registered, being measured the stem diameter and height into three environmental compartments, Haplorthox, Ustorthent and Udorthent. In Haplorthox, Araucaria angustifolia (Bertol.) Kuntze prevail at higher stratum and Sorocea bonplandii (Baill.) W.C.Burger, Lanj. & Boer, Cabralea canjerana (Vell.) Mart., Ocotea lancifolia Mez and Apuleia leiocarpa J.F.Macbr. follow it. In Ustorthent araucaria is absent and Gymnanthes concolor Müll.Arg., Trichilia claussenii C.DC., Apuleia leiocarpa, Pilocarpus pennatifolius Lem. and Sorocea bonplandii prevail. Aracuaria is scarce in Udorthent where Parapiptadenia rigida (Benth.) Brenan, Alchornea triplinervia Müll.Arg., Luehea divaricata Mart., Erythrina falcata Benth. and Myrsine gardneriana A.DC. prevail. A comparative analysis shown Cruce Caballero more similar to seasonal forests than mixed araucaria forest from Brazil. Keywords: Araucaria angustifolia; fitossociology; similarity. INTRODUCCIÓNEntre los diversos tipos forestales encontrados en Argentina, la Selva de Misiones (Selva Paranaense) es la más compleja, biodiversa y a la vez una de las menos estudiadas. La situación geográfica privilegiada, desde el punto de vista de los corredores de dispersión de especies constituidos por los grandes ríos Paraná, Uruguay e Iguazú sumada a la heterogeneidad geomorfológica, hacen de Misiones un campo de pruebas biogeográfico para todas las especies de seres vivos. Así, el planalto noreste de la provincia es una continuación del planalto catarinense brasileño, con toda la carga de especies vegetales. La cobertura forestal natural de Misiones no superaría el millón de hectáreas en la actualidad las cuales 459.700 hectáreas están encuadradas como unidades de conservación en varias categorías de manejo (CHEBEZ y HILGERT, 2003). Según Morellato y Haddad (2000), 100 millones de brasileños viven en 3000 ciudades erigidas en tierras ganadas al Bioma Mata Atlántica. Algunas de ellas son las más grandes de América Latina, como São Paulo y Rio de Janeiro. En Brasil, el área de cobertura selvática no supera el 3% de la superficie original y en Paraguay se está repitiendo el mismo proceso de sustitución o conversión. El avance de la frontera agropecuaria y el incremento de superficies plantadas con monocultivos forestales (Pinus spp.) hacen prever disminuciones drásticas de la actual superficie de selvas en Misiones. De esta forma, se torna urgente la necesidad de evaluar y conocer la estructura y diversidad de todos los tipos de selva presentes en Misiones para poder dar soporte técnico-científico para futuros emprendimientos de restauración de fragmentos, con criterios que se basen en conocimientos amplios de la base forestal actual. Es muy importante conocer también el grado de similitud de las selvas de Misiones con otras de los estados vecinos de Brasil y Paraguay, es decir con un enfoque regional. Estudios comparativos de la florística entre fragmentos pueden ser realizados por medio de análisis estadísticos multivariados como el análisis de agrupamiento (Cluster) (MEIRA NETO y MARTINS, 2002). Tales métodos permiten la visualización de la proximidad florística entre diferentes tipos forestales, lo que resulta útil para el entendimiento de la fitogeografía local. Los objetivos del presente trabajo son analizar la estructura de las principales especies arbóreas que caracterizan a tres compartimientos ambientales dados por tipos de suelo diferentes y evaluar la similitud del Parque Cruce Caballero con otros fragmentos de la región. MATERIAL Y METODOS Características del área de estudioEste estudio fue realizado en el Parque Provincial Cruce Caballero, Misiones, Argentina (Figura 1), situado entre las coordenadas geográficas 26º30´S y 53º56´O, a una altitud media de 600 m.s.n.m. De acuerdo con la clasificación climática de Köppen, la región presenta un clima tipo subtropical o templado húmedo tipo Cfa. En la región predominan los suelos Latossolos Vermelhos, Neossolos Litólicos con menor superficie y los Gleissolos son los menos expresivos. MétodosSe adopto en este trabajo el método de muestreo en parcelas múltiples, el área de cada parcela fue de 200 m2 (10 x 20 m2) para la evaluación fitosociológica de las especies. Se establecieron tres compartimientos denominados conforme, al Sistema Brasilero de Clasificación de Suelos (EMBRAPA, 2006), Latosolo Vermelho Distrófico típico (LV), Neosolo Litólico Eutrófico típico (RL) e Gleissolo Melânico Distrófico típico (GM). En cada compartimiento, las unidades de muestreo fueron distribuidas de forma sistemática, siendo instaladas 54 en Latossolo Vermelho, 20 en el Neossolo Litólico y 8 en Gleissolo Melanico, totalizando 82 parcelas, lo que corresponde a un área muestreada de 16.400 m2 para la evaluación fitosociológica. En las parcelas fueron registrados los individuos con diámetro a la altura del pecho (DAP) mayores a 5 cm. Fue colectado material botánico de la mayoría de las espécies encontradas. Las exsicatas fueron depositadas en el Herbário de la Escuela de Florestas de la UFPR. Los nombres científicos fueron establecidos, según APG II (2003). Los datos fueron procesados con auxilio del programa FITOPAC 1, desarrollado por el Dr. George John Shepherd, del Instituto de Botánica de la Universidad de Campinas. Los parámetros fitosociológicos considerados fueron los de densidad, frecuencia y dominancia, absolutas y relativas, y el porcentual de importancia (MUELLER DOMBOIS y ELLENBERG, 1974). Se analizó la distribución diamétrica de las principales especies arbóreas de cada compartimiento. Para la evaluación de la similitud entre fragmentos se utilizó el índice de Jaccard. Un análisis de agrupamiento fue realizado a partir de una matriz constituida por los valores de similitud de las diez áreas presentadas en la Tabla 1 y consideradas para lo que se utilizó el programa STATGRAPHICS PLUS versión 4.1 y siguiendo el ejemplo de Lopes et al. (2004), se utilizó el Método Aglomerativo Jerárquico, que permite la unión de áreas similares. Las secuencias de agrupamiento se presentan a través de un dendrograma utilizando el método Ward de ligación. La clasificación de la vegetación adoptada en este trabajo es la de Martínez Crovetto (1963), para Misiones, y del IBGE, según Veloso et al. (1991), para Brasil. Así, Martínez Crovetto (1963) clasificó a Misiones con los sectores forestales: el Misionero y el Planaltense. Siendo el sector Misiones dividido en distritos de los Laureles, del Palo Rosa y de los Helechos Arborescentes. Se intentó adaptarlos al sistema IBGE para hacer las comparaciones. RESULTADOS La riqueza de especies en el área estudiada es de 116, distribuidas en 46 familias y 84 géneros. Las familias de mayor riqueza de especies fueron Fabaceae (Leguminosae) y Myrtaceae, seguidas de Euphorbiaceae, Lauraceae, Meliaceae y Rutaceae. La presentación de los datos en forma diferenciada por compartimiento permite evaluar la estructura de las poblaciones arbóreas con mayor grado de detalle. Es importante destacar que los tres compartimientos presentan similitud florística principalmente el Latossolo Vermelho con Neossolo Litólico (RÍOS et al., 2008). En Latossolo Vermelho, la familia Fabaceae se mantiene con la mayor riqueza (16 especies), seguida de Meliaceae (6 spp.), Euphorbiaceae y Lauraceae (5 spp.) además de Myrtaceae y Rutaceae, con cuatro especies. En el compartimiento Neossolo Litólico, Fabaceae continúa con la mayor riqueza (9 spp.), seguida de Myrtaceae (7 spp.) y Meliaceae (6 spp.). La familia Lauraceae presento cinco especies, al igual que en el compartimiento anterior, además de Euphorbiaceae y Rutaceae, con cuatro especies. En el compartimiento Gleissolo Melánico, Fabaceae continua prevaleciendo en riqueza, pero sin ser marcada la diferencia con las demás familias y acompañada de Meliaceae con cinco especies. Las siguientes familias son Euphorbiaceae y Lauraceae, con cuatro especies, y finalmente Myrtaceae y Rutaceae, con dos especies. Teniendo en cuenta las diez principales especies según el porcentual de importancia (PI) de cada compartimiento, se considera que son representativas de la estructura poblacional de sus respectivos ambientes ya que la suma del PI del Latossolo alcanza el 41,7%, el Neossolo Litólico el 44,77% y el Gleissolo Melânico alcanza el 59,07%. En Latossolo Vermelho, las especies predominante en ordenación decreciente de PI son Sorocea bonplandii (Baill.) W.C.Burger, Lanj. & Boer, Cabralea canjerana (Vell.) Mart., Ocotea lancifolia Mez y Araucaria angustifolia (Bertol.) Kuntze (Tabla 2). Las mayores densidades absolutas fueron presentadas por Sorocea bonplandii, Ocotea lancifolia, Apuleia leiocarpa J.F.Macbr., sumando el 22,89% del total. Las tres primeras especies en orden de dominancia absoluta fueron Araucaria angustifolia, Cabralea canjerana, Apuleia leiocarpa, que suman 29,91% del total y 10,46 m2 ha-1. Sorocea bonplandii predomina en PI a pesar de ser árboles del sotobosque, de pequeño diámetro por su presencia en la mayoría de las unidades de muestreo y por su alta densidad (Figura 2). Cabralea canjerana, con valor de PI apenas inferior que el anterior, presenta menor número de individuos por hectárea, pero tiene ejemplares de grandes diámetros. Un comportamiento similar presenta Araucaria angustifolia, con menor densidad absoluta que las anteriores, pero con la mayor dominancia absoluta del compartimiento. La Figura 2 representa la distribución de diámetros mínimos, máximos y medio de las 10 principales especies. Se puede observar el predominio de Sorocea bonplandii y Myrsine umbellata Mart. en el sotobosque por los diámetros pequeños y por las altura no mayores a 20 metros (Figura 3), en Sorocea bonplandii y Myrsine umbellata no superando los 12 metros. Ocotea lancifolia, Apuleia leiocarpa, Alchornea triplinervia Müll.Arg., Nectandra lanceolata Nees & Mart. y Ocotea puberula Nees presentan ejemplares desde los estratos inferiores hasta el dosel. Cabralea canjerana presenta algunos ejemplares en los estratos inferiores y otros con alturas de hasta 35 metros. El compartimiento Neossolo Litolico presenta como principales especies Gymnanthes concolor Müll.Arg., Trichilia claussenii C.DC., Apuleia leiocarpa y Pilocarpus pennatifolius Lem. (Tabla 2). Las mayores densidades absolutas fueron dadas por Gymnanthes concolor, Trichilia claussenii y Sorocea bonplandii que suman 47,44% del total. Las tres primeras especies en orden de dominancias absolutas fueron Trichilia claussenii, Apuleia leiocarpa y Gymnanthes concolor que suman 19,25% y 8,87 m2 ha-1. Gymnanthes concolor fue predominante del compartimiento, con árboles pequeños y con alta densidad, lo que se ve reflejado en las dominancias absolutas. Trichilia claussenii, con la mayor dominancia absoluta y alta densidad, caracteriza indiscutiblemente al compartimiento. Esta especie presentó ejemplares pequeños y de mediano porte. De la Figura 4 se puede apreciar el porte de Gymnanthes concolor, Sorocea bonplandii y Pilocarpus pennatifolius. Son árboles pequeños que dominan en el sotobosque, no superando los 25 centímetros de diámetro y los 15 metros de altura (Figura 5). Dadas las marcadas pendientes, este compartimiento no admite árboles de grandes diámetros, con el máximo alcanzado por Trichilia claussenii con 75 cm. Apuleia leiocarpa y Holocalyx balansae Micheli alcanzaron los 65 cm. Tanto Trichilia claussenii como Chrysophyllum gonocarpum Engl. alcanzaron el estrato intermedio de altura. Apuleia leiocarpa, Chrysophyllum marginatum Radlk., Hellieta apiculata Benth. y Balfourodendron riedelianum Engl. conforman el dosel de la selva. No se registraron especies con alturas superiores a los 30 metros. Las especies representativas en Gleissolo absoluta de 31 individuos.ha-1. Alchornea triplinervia Melánico fueron Parapiptadenia rigida (Benth.) y Luehea divaricata presentaron altas densidades y Brenan, Alchornea triplinervia, Luehea divaricata dominancias, considerando las condiciones limitantes Mart. y Erythrina falcata Benth. (Tabla 2). Las que del ambiente. Observando la Figura 6 se puede presentan las mayores densidades absolutas fueron afirmar que el compartimiento se caracteriza por tener Myrsine gardneriana A.DC., Alchornea triplinervia árboles pequeños y sin formar un dosel propiamente y Luehea divaricata, que sumaron 36,56% del total. dicho. Los individuos están aislados formando isletas Las mayores dominancias absolutas del de selva. La especie de mayor tamaño registrado fue compartimiento fueron dadas por Parapiptadenia Parapiptadenia rigida con un diámetro de más de rigida, Erythrina falcata y Luehea divaricata, que 80 cm y una altura superior a 25 metros (Figura 7). sumaron 39,84% y 9,82m2 ha-1. La especie La mayoría de las especies no sobrepasan los 15 Parapiptadenia rigida alcanzó el primer lugar de PI, metros de altura y diámetros no mayores a 30 cm. Similitud florística Los valores de similitud florística entre Cruce Caballero y las 9 áreas consideradas (Tabla 3) presentan dos grandes grupos. Un primer grupo formado por las áreas Camaquã, São João do Triunfo, Teixeira Soares y Nova Prata, todas localizadas en los estados de Rio Grande do Sul y Paraná, en Brasil. El segundo grupo de seis áreas similares entre si, cuatro de ellas en Misiones y dos en Brasil. Cruce Caballero, Guaraní, San Antonio e Iguazú presentan mayor similitud siendo las áreas Santa Maria y Mata Godoy unidas al grupo con menor distancia o fuerza de similitud (Figura 8). DISCUSIÓN El predominio de las familias Fabaceae y Myrtaceae en Cruce Caballero coincide con otros estudios realizados en Misiones (PLACCI y GIORGIS, 1993; RÍOS, 2000 y TRESSENS et al., 2008). Pennington et al. (2000) mencionan, al describir las selvas estacionales del Neotrópico, que Fabaceae y Bignoniaceae dominan entre las especies arbóreas en diferentes grados según las áreas y comparten este dominio con Anacardiaceae, Myrtaceae, Rubiaceae, Sapindaceae, Euphorbiaceae, Flacourtiaceae y Capparidaceae en especies acuáticas o palustres. Oliveira Filho y Fontes (2000), buscando patrones de diferenciación florística del bioma Mata Atlántica asociadas con variables geográficas y climáticas y su relación con la Selva Amazónica y el Cerrado, registraron para la Selva Atlántica Costera el predominio de Myrtaceae (308 spp.), Fabaceae (246 spp.) y Melastomataceae (117 spp.); para la Selva Semidecidua el predominio en riqueza fue de Fabaceae (212 spp.), Myrtaceae (187 spp.) y Rubiaceae (84 spp.). Para la Selva Amazónica la familia de mayor riqueza fueron Fabaceae (252 spp.), Sapotaceae (88 spp.) y Lauraceae (79 spp.). En este estudio, Bignoniaceae está escasamente representadas con solo dos especies (Jacaranda micrantha Cham. y Tabebuia heptaphylla (Vell.) Toledo) y la familia ausente es Anacardiaceae. Similares representatividades de familias se presentan en el estado de Paraná en el Parque Estadual Mata dos Godoy en un estudio florístico realizado por Silva y Soares Silva (2000), donde detectaron 31 especies de Fabaceae y 24 especies de Myrtaceae. En Mata dos Godoy, la familia Bignoniaceae está escasamente representada con tres especies de los géneros Jacaranda, Tabebuia y Zeyheria, además de Anacardiaceae con solo dos especies. En el estado de Rio Grande do Sul, BR, Jarenkow y Weachter (2001) hallaron predominio de Fabaceae (7 spp.), seguida de Myrtaceae (6 spp.) y no registraron Anacardiaceae y Capparidaceae, al igual que la situación de Cruce Caballero. Familias no destacadas por Pennington et al. (2000) en sus descripciones son Meliaceae y Lauraceae, tal vez por ser más exigentes en ambientes húmedos. En Cruce Caballero fueron registradas con seis especies cada una, en la Mata dos Godoy Meliaceae (10 spp.), Lauraceae (11 spp.) y en una selva estacional en el estado de Rio Grande do Sul, Jarenkow y Weachter (2001) registraron Meliaceae y Lauraceae con cinco especies cada una. Spichiger et al. (2004) afirman que varias especies de Meliaceae, como Cedrela fissilis Vell., Trichilia elegans A.Juss. y Cabralea canjerana, son las más características en la región de selvas estacionales de Colombia, aclarando que estas especies son de amplia distribución y que se encuentran en la periferia de la región Amazónica (Arco Amazónico). El patrón de distribución actual de estas especies podría considerarse como remanentes de una mayor distribución pretérita, que sobrevive hoy solo en los márgenes de la Cuenca Amazónica. Por ejemplo, Cedrela fissilis tiene hoy una amplia distribución en todos los bordes de la Cuenca Amazónica, lo que se podría deber por el aumento de temperatura y precipitaciones en el Holoceno. Esta variación climática pudo haber causado que las especies generalistas y tolerantes a la sequía, como esta especie, puedan desplazarse a los bordes de la Cuenca Amazónica por ser reemplazada en las áreas más húmedas por elementos sensibles a la sequía. Reis et al. (2007), analizando la composición florística de fragmentos de Minas Gerais, BR, todos ellos con Selva Estacional Semidecidua, encontraron el impresionante valor de 45 especies de Fabaceae, acompañadas de Lauraceae (33 spp.), y luego Myrtaceae con 29 especies. Las Meliaceae presentaron 11 especies y Bignoniaceae siete especies. El compartimiento Latossolo Vermelho está caracterizado por ser un suelo profundo, bien drenado y de escasas pendientes. El predominio de Fabaceae es acompañada de Meliaceae, la que ocupa el lugar de las Myrtaceae, pero sin dejar estas de ser importantes. Este cambio se debe a que las Meliaceae prefieren ambientes mésicos y las Myrtaceae prefieren suelos con buena disponibilidad hídrica. Considerando las principales especies según PI, el juego de especies por familia cambia marcadamente. Lauraceae demuestra tener mayor injerencia en el ambiente. Considerando las especies más representativas, Sorocea bonplandii, Cabralea canjerana, Ocotea lancifolia y Araucaria angustifolia, las dos primeras son clímax tolerantes a la sombra y las otras dos son clímax exigentes en luz. Considerando las diez primeras especies, a las dos tolerantes a la sombra les siguen en importancia ocho exigentes en luz (Tabla 2). La alta densidad de Sorocea bonplandii se explica por su característica de ser una especie pequeña de sotobosque con gran tolerancia a la sombra. Otros estudios con igual criterio de inclusión también registraron Sorocea bonplandii como la especie predominante (RIOS et al., 1999, 2008, en Eldorado, Argentina; SILVA et al., 2005, en Parque do Turvo, Brasil; CAMPOS et al., 2006, en una Selva Estacional Semidecidual, Brasil.). Otros estudios en cambio, registraron Sorocea bonplandii con menor importancia en la comunidad, como por ejemplo Vilela et al. (2000), que la registró en el puesto 81° del valor de importancia en una Selva Semideciual Aluvial y Montana en Minas Gerais. Esta falta de protagonismo puede ser indicada a cuestiones ambientales limitantes para esta especie. Algunos autores la catalogan como especie indiferente en cuanto a condiciones de suelo (LORENZI, 1998). Similar situación encontró Longhi et al. (2000), en una Selva Decidua, en Rio Grande do Sul. Ocotea lancifolia tiene una cita para Misiones. En la zona sur (Instituto Darwinion Arg. Web) es conocida en la región como laurel pimienta. França y Stehmann (2004) la registraron en la Selva Ombrófila Densa Altimontana en Minas Gerais como principal especie en importancia, con alta densidad absoluta (167) y alta dominancia absoluta (19 m2 ha-1). En Cruce Caballero, la densidad absoluta (42) es inferior y muy diferente, la dominancia absoluta es de 1,26 m2 ha-1. Nascimento et al. (2001) la registraron en la Selva Ombrófila Mixta de Rio Grande do Sul en el puesto 16° de valor de importancia, con densidad y dominancia absoluta muy bajas comparadas con Cruce Caballero. S á (2004), en un relevamiento florístico en el estado de Paraná, en un área de confluencia de unidades fisionómicas, registró Ocotea lancifolia; Barbosa y Moraes (2007) la citan para el estado de Espírito Santo. Vasconcelos y D´Angelo Neto (2009) la citan como una de las principales especies en una Selva Ombrófila Mixta de Minas Gerais, junto a Cabralea canjerana y escasos individuos de Araucaria angustifolia, situación general muy parecida a Cruce Caballero en el estrato considerado. En este estudio, Araucaria angustifolia ocupa el 4° lugar de importancia, lo que responde a sus altos valores de dominancia ya que la densidad alcanzada es extremadamente baja (6 individuos ha-1). Esta situación corroboraría lo afirmado por Martinez Crovetto (1963) en su trabajo titulado Fitogeografía de Misiones, al decir que la Selva Subtropical (Selva Estacional Decidua y Semidecídua) va eliminando poco a poco al pino Paraná. Comparativamente, esta especie en áreas centrales de distribución (Selva Ombrófila Mixta) presenta predominio absoluto en importancia de esta especie, como registrado por Longhi (1980), Rondon Neto et al. (2002), Schaaf et al. (2006). Baja densidad de araucaria en Selva Ombrófila Mixta fue registrada por Nascimento et al. (2001) en Nova Prata y con un 3° puesto en importancia. Apuleia leiocarpa sigue en valor de importancia a la araucaria en Cruce Caballero, además de sumar altos valores de dominancia junto a esta lo que auxiliaría en el carácter estacional de la selva de Cruce Caballero. El Neossolo Litólico es un suelo raso, bien drenado de altas pendientes donde al considerar las diez principales especies de importancia, el juego de familias botánicas cambia, siendo Rutaceae la de mayor riqueza (3), seguida de Fabaceae y Sapotaceae. Se observa el predominio de Gymnanthes concolor y Trichilia claussenii. Esta situación se enmarca en el concepto de dominancia ecológica acuñada por Richards (1952) y que es definida por la marcada superioridad en importancia de pocas especies arbóreas en una comunidad. En las selvas tropicales es común que la dominancia ecológica sea más pronunciada en los extremos de alta y baja disponibilidad de recursos (CARVALHO et al., 2005). El predominio de Gymnanthes concolor en áreas de Selva Estacional Semidecidua y de pendiente pronunciada fue registrada por varios autores en estados vecinos de Brasil, como Jarenkow y Weachter (2001), Jurinitz y Jarenkow (2003), Silva et al. (2003). Esto indicaría una preferencia por suelos rasos y bien drenados. Sin embargo, esta especie fue indicada para ambientes aluviales (SILVA et al., 2007). Trichilia claussenii y Apuleia leiocarpa son árboles que alcanzan grandes dimensiones y, según López y Little (1987), Apuleia leiocarpa prefiere suelos bien drenados. Las mayores densidades de tres especies puede deberse a la dominancia ecológica que determinan por ser el compartimiento escaso en nutrientes y agua. Esta situación fue encontrada por Carvalho et al. (2005), en una selva de Tres Marias, Minas Gerais, donde el Neossolo Litólico presenta mayores densidades de individuos bajos y finos. El compartimiento Gleissolo Melánico fue el que presentó mayor riqueza de familias botánicas, pues las diez principales especies se reparten en ocho familias. Las limitaciones ambientales impuestas por el anegamiento semipermanente o permanente no lograron que se de la dominancia ecológica de pocas especies como detectada en el compartimiento Neossolo Litólico. La primera especie en importancia, Parapiptadenia rigida, se ve beneficiada por su alta dominancia, pero su valor de densidad es bajo. Algunos autores reportan a esta especie como exclusiva de suelos bien drenados y profundos (BRACK y WEIK, 1993), características no halladas en este compartimiento. Según Silva et al. (2007), la ocurrencia de especies arbóreas en áreas inundables puede seguir varios patrones de distribución geográfica, siendo que Luehea divaricata fue la especie encontrada con mayor frecuencia en el sur y sudeste de Brasil. Estudios en áreas inundables del sureste de Brasil, registraron Parapiptadenia rigida como preferencial de ambientes aluviales, es decir de inundaciones no permanentes. En Cruce Caballero esta especie se halla en ambientes anegados y Alchornea triplinervia y Luehea divaricata se presenta como no preferenciales, pues Alchornea triplinervia se encontró en todos los compartimientos, ya Luehea divaricata se la encontró en dos compartimientos pero con marcada abundancia en el Gleissolo. Silva et al. (2007) encontraron predominio de Luehea divaricata en ambientes aluviales e inundables, pero con mayores áreas aluviales. Erythrina falcata fue catalogada como no preferencial y en Cruce Caballero se presento en dos compartimientos pero en mayor número de Gleissolo. El patrón de similitud florística resultante del análisis de agrupamiento evidenció la existencia de dos grandes grupos. El primero formado por cuatro áreas de Brasil, tres de ellas conformadas de Selva Ombrófila Mixta (4-9-7) y una con Selva Estacional Semidecidua (2). El segundo grupo formado por cuatro áreas de Argentina (1-8-5-6) y dos de Brasil (3-10) (Figura 8). Cruce Caballero mostró mayor similitud con Guaraní que con San Antonio, a pesar de ser esta última área la que comparte la presencia de araucaria junto a Cruce Caballero. Guaraní no registra la presencia de la especie. Esta situación corroboraría la afirmación hecha por numerosos autores referente a las selvas con araucaria de Argentina de que esta formación arbórea es un ecotono con las selvas estacionales. El Parque Nacional Iguazú, al norte de Misiones, está muy cercana a las anteriores en su composición florística. Más alejadas o mas disimilares están Santa María y Mata dos Godoy. Santa Maria con Selva Decidua de Rio Grande do Sul y Mata dos Godoy con Selva Semidecidua del estado de Paraná. Es notable observar como Camaquã, presentando Selva Semidecidua, es disimilar a la Mata dos Godoy, que también posee Selva Semidecidua, pero tiene su explicación si consideramos la gran distancia geográfica que las separa. El 77% de las especies de Cruce Caballero están presentes en Guaraní, a excepción de araucaria, que se presume fue extirpada (BORGES com. Pers.) del área y más teniendo en cuenta que en el predio vecino (Parque Caa Yarí) se encuentra la especie con algunos ejemplares adultos. Comparada con San Antonio, comparte el 56,6% de sus especies y con Iguazú comparte el 48,6%. Santa María presenta Selva Decidua y contiene el 33,6% de las especies de Cruce Caballero. La Mata de Godoy contiene el 64,6% de las especies de Cruce, el doble de lo que contiene Santa Maria. Podríamos esperar que Mata Godoy debiera estar a menor distancia o debería ser más similar a Cruce Caballero si no fuera que Guaraní, San Antonio e Iguazú son más similares a Santa María que a Mata Godoy. Esta situación hace que Santa Maria se posicione a menor distancia de las cuatro áreas de Argentina. CONCLUSIONESEl Parque Provincial Cruce Caballero presenta una vegetación arbórea con especies diferenciadas según los tres compartimientos considerados. El Latossolo Vermelho fue caracterizado por Sorocea bonplandii y Cabralea canjerana. La Araucaria angustifolia presento ejemplares adultos con alta dominacia absoluta, pero con escasa densidad absoluta. El Neossolo Litólico presenta dominancia ecológica de Gymnanthes concolor y Trichilia claussenii y ejemplares pequeños, dadas las limitantes ambientales del compartimiento. Gleissolo Melánico presentó predominio de Parapiptadenia rigida. La selva del Parque Cruce Caballero se asemeja florísticamente a las selvas estacionales de Brasil y no presenta similitud con la Selva Ombrófila Mixta de Brasil. Las mayores similitudes se establecieron con Guaraní, San Antonio, Iguazú, Santa Maria (Brasil) y Mata dos Godoy (Brasil). Se concluye que la selva de Cruce Caballero es un ecotono entre las selvas estacionales Decidua y Semidecidua con la Selva Ombrófila Mixta de Brasil. REFERENCIAS
Copyright 2010 - Ciência Florestal The following images related to this document are available:Photo images[cf10016t2.jpg] [cf10016f3.jpg] [cf10016f7.jpg] [cf10016f1.jpg] [cf10016f8.jpg] [cf10016f2.jpg] [cf10016t1.jpg] [cf10016f6.jpg] [cf10016f4.jpg] [cf10016f5.jpg] [cf10016t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}