 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ciência Florestal, Vol. 20, No. 2, Apr-Jun, 2010, pp. 207-215 CRESCIMENTO E NUTRIÇÃO MINERAL DO NIM (Azadirachta indica A. Juss.) E CINAMOMO (Melia azedarach Linn.) SUBMETIDOS À SALINIDADE GROWTH AND MINERAL NUTRITION OF NEEM (Azadirachta indica A. Juss.) AND CHINABERRY TREE (Melia azedarach Linn.) SUBMITTED TO SALINITY Antonio Lucineudo de Oliveira Freire1 Gilberto Marinho de Sousa Filho2 José Romilson Paes de Miranda3 Patrícia Carneiro Souto4 Lúcio Valério Coutinho de Araújo5
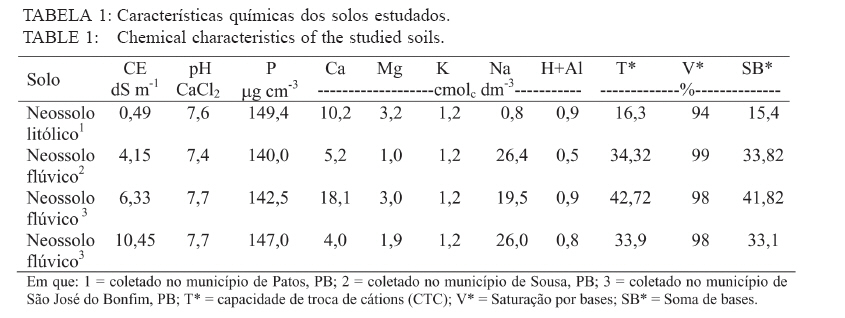
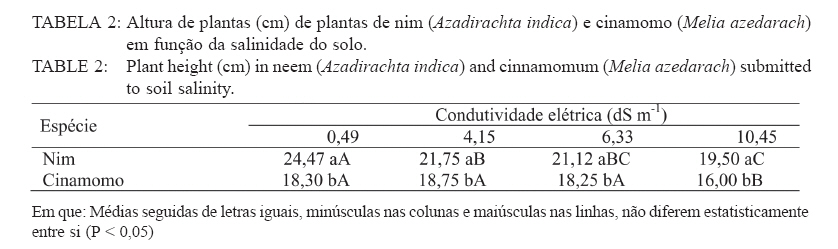
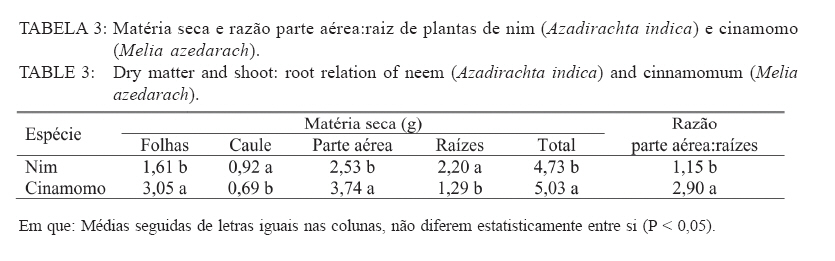
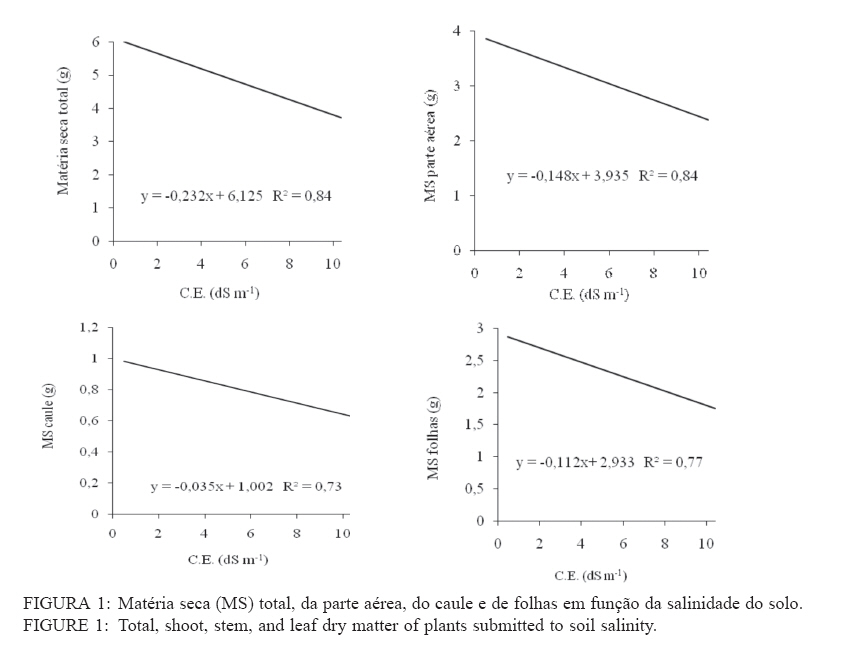
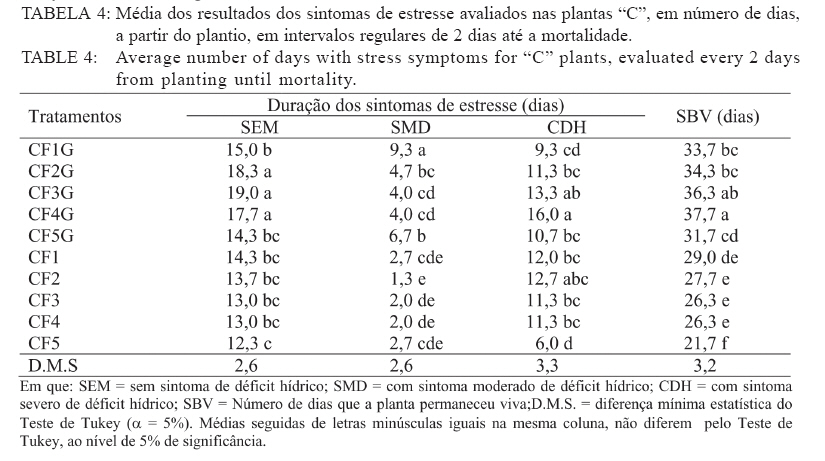
Recebido para publicação em 26/04/2005 e aceito em 05/04/2010. Code Number: cf10017 RESUMO Este trabalho teve como objetivo verificar os efeitos da salinidade do solo no crescimento e no acúmulo de macronutrientes e de sódio em nim (Azadirachta indica) e cinamomo (Melia azedarach). O experimento foi disposto em delineamento experimental inteiramente casualizado em arranjo fatorial 2 x 4, sendo duas espécies (nim e cinamomo) e quatro níveis de salinidade (Condutividade elétrica (C.E.) 0,49 (solo não salino); 4,15; 6,33 e 10,45 dS m-1), com 4 repetições. As plantas cresceram inicialmente em tubetes e, 60 dias após a emergência, foram transferidas para vasos contendo 3 kg de substrato [solo + esterco (2:1)], de acordo com o tratamento de salinidade. Após 45 dias foram avaliadas altura das plantas, matéria seca (folhas, caule, parte aérea (caule + folhas), raízes e total) e acúmulo de N, P, K, Ca, Mg, S e Na+ na parte aérea. A salinidade reduziu a altura das plantas em ambas as espécies, mas o efeito foi mais pronunciado no nim. Elevação na salinidade dos solos causou aumento no acúmulo de Na+ e redução no acúmulo de macronutrientes na parte aérea em ambas as espécies, principalmente no nim. O cinamomo foi mais tolerante aos níveis de salinidade dos solos do que o nim. Palavras-chave: estresse salino; tolerância à salinidade; Meliaceae. ABSTRACTThe objective of this study was to verify the effects of soil salinity on growth and nutrient and sodium accumulation in neem (Azadirachta indica) and cinnamomum (Melia azedarach). The experimental delineation was completely randomized in a 2 x 4 factorial arrangement, with two species (neem and cinnamomum), four salinity levels (electrical conductivity 0.49 (non saline soil), 4.15, 6.33 and 10.45 dS m1) and four replications. Initially, plants were grown in tubes, and 60 days after emergence, they were transferred to pots containing 3 kg of substrate [soil + manure (2:1)], according to the saline treatment. After 45 days, plant height, dry matter (leaves, stem, shoot (stem + leaves), roots and total) and N, P, K, Ca, Mg, S and Na+ shoot accumulation were evaluated. Salinity reduced the plant height in both species, but the effect was more pronounced in neem. Increases in soil salinity caused an increase in the accumulation of Na + and reduced the accumulation of nutrients in shoots of both species, especially in neem. The cinnamomum was more tolerant to salinity levels of soils than neem. Keywords: salt stress; salt tolerance; Meliaceae. INTRODUÇÃOAs regiões áridas e semiáridas, que constituem cerca de 40% da superfície da Terra, caracterizam-se por períodos de seca prolongados e temperaturas altas, que limitam a produtividade das plantas. Somado a tais fatores, está a salinidade do solo e estima-se que cerca da metade dessas terras é afetada pela salinização (BRADBURY e AHMAD, 1990; SERRATO VALENTI et al., 1991), problema que está aumentando em virtude do emprego de técnicas agrícolas de maneira incorreta, como adubação excessiva e irrigação com água imprópria para tal finalidade, transformando terras férteis e produtivas em áridas (SERRATO VALENTI et al., 1991). Dentre os elementos que contribuem para a salinização dos solos, os principais são Ca, Mg, Na, K, Cl e S. Quando o teor de sódio é alto em relação aos demais cátions, este pode ser adsorvido pelo complexo de troca e, nesse caso, as partículas de argila podem dispersar-se e o solo perde sua estrutura, tornando-se impermeável (PIZARRO, 1978). O excesso de sais promove a retenção de água no solo, tornando-a cada vez menos acessível às plantas, além do efeito de alguns íons no protoplasma (JEFFREY e IZQUIERDO, 1983; MUNNS, 1993). A habilidade das plantas em sobreviver sob condições salinas é importante para sua distribuição geográfica e para a agricultura nas regiões salinizadas. É necessário que se utilizem espécies que tolerem essa condição e, se possível, que sejam capazes de melhorar as características físicas e químicas desse solo, o que pode ser conseguido por meio do plantio de espécies arbóreas de crescimento rápido, tolerantes à salinidade. No entanto, para que se obtenha sucesso, é necessário que se conheçam os efeitos da salinidade no desenvolvimento da espécie a ser empregada e o seu grau de tolerância a essa condição adversa. Em termos de potencial madeireiro de grande porte, as áreas semiáridas do nordeste brasileiro apresentam baixo potencial quando comparadas com espécies nativas de porte arbóreo-arbustivo, utilizadas como fonte de matéria-prima para fins energético e forrageiro. O uso de madeira no estado da Paraíba para fins energéticos é intenso e de importante contribuição, tendo em vista que a lenha e o carvão vegetal se incluem entre as principais fontes de energia para o uso doméstico e industrial. Diversas espécies florestais exóticas foram introduzidas na região, podendo-se destacar a algaroba (Prosopis julifora) e a leucena (Leucaena leucocephala) entre as espécies potenciais. No entanto, deve-se dar atenção a outras espécies madeireiras exóticas capazes de se adaptar às condições climáticas e edáficas da região e que sirvam como opção para reduzir as pressões sobre a vegetação nativa. O nim ou amargosa (Azadirachta indica A. Juss.), pertencente à família Meliaceae, é uma espécie subtropical, nativa das regiões áridas da Ásia e África e encontra-se distribuída também na Austrália e América (SCHMUTTERER, 1990; MORDUE e BLACKWELL, 1993). Prefere climas tropicais e subtropicais, com precipitação pluvial anual entre 400-800 mm; é tolerante a altas temperaturas, resistindo a longos períodos secos (NEVES et al., 2003). A madeira tem coloração avermelhada, é dura e resistente a ataques de térmitas ou apodrecimento. O cerne é muito rico em tanino e sais inorgânicos de cálcio, potássio e ferro (NEVES e NOGUEIRA, 1996). Em plantios na região semiárida do Nordeste brasileiro, a madeira de nim apresentou densidade básica de 0,57 g/cm3 e porcentagem de carbono fixo de 81,82%, o que revela seu grande potencial para fins energéticos (ARAÚJO, 1999). O cinamomo (Melia azedarach Linn.), natural do sul da Ásia, possui madeira leve, sendo uma espécie indicada para uso em obras expostas, marcenaria, caixotaria, lenha (SANTOS, 1987). Tanto o nim como o cinamomo apresentam ação inseticida, em virtude da presença da azadiractina, um tetranortriterpenóide, que regula o crescimento e a metamorfose dos insetos, causa deterrência alimentar, afeta a biologia, a oviposição e a viabilidade dos ovos dos insetos (SCHMUTTERER, 1990; MORDUE e BLACKWELL, 1993; NEVES e NOGUEIRA, 1996; SALLES e RECH, 1999; TRINDADE et al., 2000). Maroneze e Gallegos (2009) verificaram que o extrato aquoso de folhas de Melia azedarach provocou mortalidade total de lagartas de Spodoptera frugiperda nas concentrações de 1 e 5%, e que na concentração de 0,1%, o extrato afetou negativamente o peso de lagarta se pupas e reduziu acentuadamente a fecundidade dos adultos. Em virtude da falta de informações a respeito dessas espécies quanto à tolerância ao estresse salino, desenvolveu-se este trabalho com o objetivo de avaliar o crescimento e a nutrição mineral de plantas de nim e de cinamomo crescendo em solos salinos visando, dessa forma, a obter informações que possibilitem a utilização dessas espécies para recuperação das áreas com problemas de salinidade do solo, constituindo-se também numa opção para suprir a demanda por madeira, diminuindo a pressão sobre as espécies nativas. MATERIAL E MÉTODOS Localização e descrição do local do experimentoO experimento foi conduzido em telado no Viveiro Florestal pertencente ao Departamento de Engenharia Florestal da Universidade Federal de Campina Grande, Campus de Patos, PB, localizado nas coordenadas geográficas 7o03’34” S e 37o16’19” O, altitude de 250 m. Delineamento experimentalO delineamento experimental utilizado foi inteiramente casualizado, em esquema fatorial 2 x 4, consistindo de duas espécies vegetais (nim e cinamomo) e quatro solos com salinidades diferentes (Condutividade elétrica (CE) 0,49 (solo não salino); 4,15; 6,33 e 10,45 dS m-1), com quatro repetições. Condução do experimentoAs sementes de nim e de cinamomo foram procedentes de árvores matrizes, com 7 anos de idade, crescendo no Viveiro Florestal da Universidade Federal de Campina Grande, em Patos, PB. A semeadura ocorreu diretamente em tubetes com capacidade para 250 cm3, contendo o substrato constituído de uma mistura de subsolo + esterco, na proporção 2:1. Aos 60 dias após a emergência, plantas uniformes quanto à altura foram transferidas individualmente para vasos contendo 3 kg de substrato (solo + esterco, na proporção 2:1). Com exceção do solo não salino, os demais foram coletados em áreas de perímetros irrigados, na profundidade de 0-20 cm, e submetidos a análises químicas (Tabela 1). Os vasos foram irrigados diariamente, de maneira a atingir 50% do volume total de poros (VTP), mantidas por meio de pesagem. Aos 45 dias após a transferência, avaliou-se a altura das plantas. Em seguida, estas foram cortadas e separadas em folhas (lâmina foliar e pecíolo), caule e raízes. Esses componentes foram separadamente acondicionados em sacos de papel e transferidos para estufa a 70oC, até obtenção da massa constante. Após a secagem, esses componentes foram submetidos à pesagem, para a determinação da matéria seca de folhas, caule, raízes, parte aérea (caule + folhas) e matéria seca total das plantas. Em seguida, realizou-se a moagem da parte aérea para a determinação dos teores de nitrogênio, fósforo, potássio, cálcio, magnésio, enxofre e de sódio. No extrato obtido por digestão nítricoperclórica, foram determinados os teores de P por colorimetria; Ca e Mg, por espectrofotometria de absorção atômica; K e Na por fotometria de chama de emissão; e, de S, por turbidimetria do sulfato de bário (MALAVOLTA et al., 1997). Os teores de N total foram determinados pelo método semimicro Kjeldahl, sendo a destilação e a titulação realizadas segundo Bremner e Edwards (1965). Partindo desses teores, determinou-se o acúmulo dos nutrientes com base na matéria seca. Os dados foram submetidos à análise de variância e teste de médias (Tukey, 5%) e, quando necessário, à análise de regressão, utilizando-se dos procedimentos disponíveis no programa estatístico SISVAR (FERREIRA, 2000). RESULTADOS E DISCUSSÃO Crescimento e distribuição de matéria seca das plantasDe acordo com a análise estatística, houve interação significativa entre espécies e níveis de salinidade apenas para a altura, enquanto que para a matéria seca o efeito foi isolado, tanto da espécie quanto da salinidade. Na Tabela 2, percebe-se que houve redução na altura das plantas à proporção que a salinidade do solo aumentou e que, comparando-se as espécies dentro de cada nível de salinidade, verifica-se que o nim apresentou as maiores alturas. No entanto, apesar de apresentarem maior desenvolvimento, estas foram mais afetadas pela salinidade, pois quando se compara as plantas que não foram submetidas à salinidade com aquelas submetidas à salinidade mais elevada, nota-se que o nim apresentou redução de 20% na altura, enquanto que no cinamomo essa redução foi de apenas 12,5%. Comparando-se apenas a espécies estudadas, independentemente do tratamento de salinidade, verifica-se que as plantas de nim apresentaram maior matéria seca do caule e das raízes, enquanto que, nas plantas de cinamomo, esses valores foram mais elevados nas folhas, na parte aérea e total (Tabela 3). No que se refere à razão parte aérea:raízes, verificase que as plantas de cinamomo acumularam quase três vezes mais matéria seca na parte aérea do que nas raízes, mostrando que, independente do nível de salinidade empregado, aquelas direcionaram mais assimilados para a parte aérea. Em relação aos efeitos da salinidade, verifica-se redução linear no acúmulo de matéria seca nas plantas com a elevação no nível de salinidade do solo, independentemente da espécie estudada (Figura 1). Comparando-se os valores de matéria seca das plantas do tratamento não salino com aquelas mantidas no solo com salinidade mais elevada, observa-se redução de 39, 35, 38 e 38%, respectivamente na matéria seca de folhas, caule, parte aérea e total. Dessa forma, percebe-se que os efeitos da alta salinidade ocorreram de maneira mais ou menos uniforme, em todas as partes da planta. As plantas de cinamomo podem ter promovido ajustamento osmótico das suas folhas pelo acúmulo de solutos, garantindo assim a manutenção da turgescência celular e o consequente crescimento. Essas informações mostram que os processos afetados pela salinidade, os quais resultam em redução no crescimento das plantas, variam com a espécie e a concentração de sal empregado e que, apesar das evidentes alterações no crescimento e no processo fotossintético, em virtude da salinidade, a natureza exata das mudanças no metabolismo do carbono em virtude do estresse salino não está ainda totalmente elucidada (MUNNS, 1993). Redução na taxa fotossintética pode ocorrer, pelo menos parcialmente, em função de fatos não relacionados ao comportamento osmótico, sugerindo que metabolismo do cloroplasto, e consequentemente processos de partição do carbono, são também afetados pelo estresse salino (EVERARD et al., 1994). Esses autores afirmam ainda que os efeitos dos sais no crescimento das plantas estão mais relacionados com a salinidade total do que com as concentrações individuais de algum constituinte específico. Por promover várias alterações fisiológicas, a salinidade afeta o crescimento e o desenvolvimento das plantas, em especial das glicófitas (EL-HADDAD e O’LEARY, 1994). A presença de sais na solução do solo reduz o seu potencial osmótico, afetando a capacidade de absorção radicular da planta e consequentemente o seu metabolismo, além do efeito direto dos íons, causando perturbação nutricional da planta, dificultando dessa forma o seu desenvolvimento (VIEIRA, 1989). A salinidade afeta a absorção de água e íons e o metabolismo de carboidratos, o que reduz o crescimento (SERRATO VALENTI et al., 1991) e o acúmulo de matéria seca (FREIRE, 2000; MIRANDA, 2000). Vários fatores contribuem para a redução no crescimento das plantas sob estresse salino, como diminuição na absorção de água, redução na taxa fotossintética e toxicidade de íons, mais especificamente Na+ e Cl-. Em plantas de Atriplex prostrata, Wang et al. (1997) verificaram que o aumento da salinidade do substrato promoveu redução da altura e da matéria seca, e atribuíram esse comportamento à redução na taxa de fotossíntese das plantas em virtude da redução na condutância estomática. No entanto, nem sempre as taxas de fotossíntese são afetadas pelos sais, conforme verificado em Trifolium repeus (ROGERS e NOBLE, 1992). Não apenas a área foliar diminui em função da salinidade, mas também a fixação líquida do CO2 por unidade de área foliar (MARSCHNER, 1995). Acúmulo de macronutrientes e Na+ na parte aérea das plantasVerificou-se interação significativa entre as espécies e os níveis de salinidade estudados, para todos os nutrientes avaliados. Em ambas as espécies, houve redução no acúmulo de N, P, K, Ca, Mg e S, e aumento no acúmulo de Na+, com o aumento da salinidade do solo (Tabela 4). Nas plantas de nim, comparando-se o solo com o maior nível de salinidade com o solo não salino, verifica-se redução no acúmulo de todos os macronutrientes em cerca de 50%. Quanto ao Na+, as plantas que cresceram no solo com o maior nível de salinidade apresentaram acúmulo cerca de seis vezes o valor encontrado naquelas crescidas no solo não salino, mostrando que as plantas não foram eficientes em regular a absorção e translocação desse elemento para a parte aérea. No cinamomo, as reduções no acúmulo dos nutrientes foram menores do que os verificados no nim. Quando se compara o tratamento de alta salinidade com o tratamento não salino, verificamse reduções de 22, 8, 51, 51, 46 e 21%, respectivamente, para N, P, K, Ca, Mg e S. Quanto ao acúmulo de Na+ nesses tratamentos, o valor observado no tratamento de salinidade mais elevado foi 5,4 vezes o valor observado no tratamento não salino. Com exceção do Ca e S, no tratamento CE 6,33 dS m-1, os acúmulos dos nutrientes foram sempre maiores nas plantas de cinamomo. Apesar de terem apresentado menor altura, essas tiveram maior acúmulo de matéria seca na parte aérea, o que pode ser explicado pelo maior acúmulo de nutrientes na parte aérea. Quando se analisa a altura (Tabela 2) e o acúmulo dos macronutrientes (Tabela 4), no cinamomo, verifica-se que não ocorreram alterações significativas nesses valores com o aumento da CE para 4,15 dS m-1, e mesmo com o aumento para 6,33 dS m-1, a altura destas permaneceu estatisticamente igual, apesar das reduções nos acúmulos dos macronutrientes. Isso permite inferir que o cinamomo conseguiu manter seu crescimento em solo com CE 6,33 dS m-1 (Figura 1), mesmo que houvesse prejuízo no acúmulo de macronutrientes e elevação no acúmulo de Na+ na parte aérea, o que pode caracterizar tolerância a esse nível de salinidade do solo. Esse acúmulo de solutos inorgânicos pode ter garantido ajustamento osmótico por parte das plantas, numa tentativa de manter seu crescimento ou, pelo menos, reduzir os efeitos da alta concentração de sais no solo. O acúmulo de íons inorgânicos com a finalidade de ajustamento osmótico é, segundo Yeo (1983), de grande importância para as plantas superiores conciliarem produtividade com tolerância ao estresse hídrico. Esse ajustamento osmótico pode ser conseguido por meio do acúmulo tanto de solutos orgânicos como inorgânicos, reduzindo o potencial osmótico intracelular e mantendo o potencial de turgor positivo, garantindo a manutenção dos processos de crescimento e alongamento celulares (HSIAO, 1973), a abertura estomática e a fotossíntese (LUDLOW et al., 1985; NEUMANN et al., 1988). No entanto, o aumento das concentrações de íons no simplasto pode eventualmente gerar toxicidade (GREENWAY e MUNNS, 1980). A redução no acúmulo de macronutrientes em função da salinidade do solo está de acordo com alguns relatos encontrados na literatura quanto ao efeito depressivo do sal no acúmulo de nutrientes (BARBOSA et al., 1996; KURBAN et al., 1998; FREIRE et al., 1999; MIRANDA et al., 2002; MIRANDA et al., 2004; AQUINO et al., 2007). Perez-Alfocea et al. (1993) afirmaram que os efeitos da salinidade sobre a concentração de N dependem do nível de salinidade, do tempo de exposição das plantas ao estresse salino, das diferenças genotípicas quanto à tolerância e, de acordo com Botella et al. (1997), também da fonte de N empregada, além do fato de que a presença de NaCl inibe a absorção de NO-3, provavelmente devido ao efeito depressivo do Cl, mas não afeta a absorção de NH+4. Segundo Lynch e Läuchli (1985), o NaCl inibe o movimento radial do Ca2+ em direção ao xilema, podendo também promover desconfiguração dos carregadores para esse nutriente. Concordando com o observado nas plantas de nim e cinamomo, Devitt et al. (1993) verificaram em plantas de Cynodon dactylon que a redução no transporte de potássio para a parte aérea diminuiu com o aumento do sódio. Em plantas de feijão, Gouia et al. (1994) verificaram redução nas concentrações de potássio, cálcio e magnésio na parte aérea quando foram submetidas a 50 mmol L-1 de NaCl. Comportamento semelhante foi verificado por Kurban et al. (1998), em que aumentos nas concentrações de sódio e cloro foram acompanhados por redução nas concentrações de potássio, cálcio, fosfato e nitrato. Hsiao (1973) afirma que quando a turgescência diminui, ocorrem alteração na distribuição espacial das proteínas e a função da membrana é modificada, diminuindo sua permeabilidade à água e íons. Existem diferenças genotípicas entre as espécies vegetais quanto à permeabilidade da membrana, mas as bases bioquímicas e fisiológicas da menor permeabilidade da membrana não estão bem esclarecidas (KURBAN et al., 1998). Premachandra et al. (1991) sugeriram que o ajustamento osmótico por causa do acúmulo de solutos, em especial potássio e cálcio, diminui a permeabilidade da membrana. No entanto, essa afirmação discorda dos resultados obtidos por Kurban et al. (1998), os quais verificaram que a menor permeabilidade da membrana em plantas de Alhagi pseudoalhagi não foi em consequência do acúmulo de potássio e cálcio nos tecidos, pois as concentrações desses solutos inorgânicos diminuíram com o aumento da salinidade. CONCLUSÕESA salinidade reduziu a altura das plantas em ambas as espécies, mas o efeito foi mais pronunciado no nim; Elevação na salinidade dos solos causou aumento no acúmulo de Na+ e redução no acúmulo de macronutrientes na parte aérea em ambas as espécies; O cinamomo foi mais tolerante aos níveis de salinidade dos solos do que o nim. REFERÊNCIAS BIBLIOGRÁFICAS
Copyright 2010 - Ciência Florestal The following images related to this document are available:Photo images[cf10017f1.jpg] [cf10017t4.jpg] [cf10017t2.jpg] [cf10017t1.jpg] [cf10017t3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}