 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ciência Florestal, Vol. 20, No. 2, Apr-Jun, 2010, pp. 255-267 CARACTERÍSTICAS FISIOLÓGICAS DE MUDAS DE Eucalyptus spp SUBMETIDAS A ESTRESSE SALINO PHYSIOLOGICAL CHARACTERISTICS OF Eucalyptus spp SEEDLINGS SUBMITTED TO SALINE STRESS Andrea Vita Reis Mendonça1 José Geraldo de Araújo Carneiro2Teresa Aparecida Soares de Freitas3 Deborah Guerra Barroso4
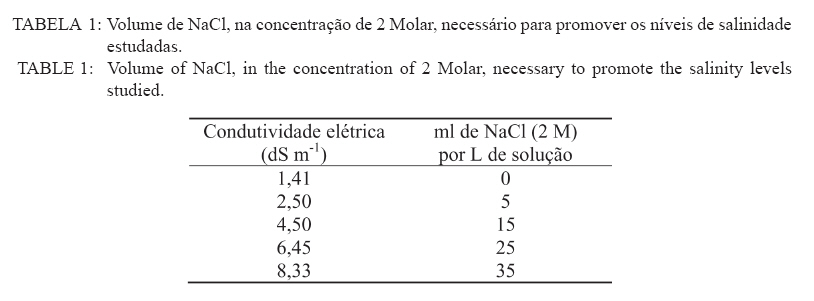
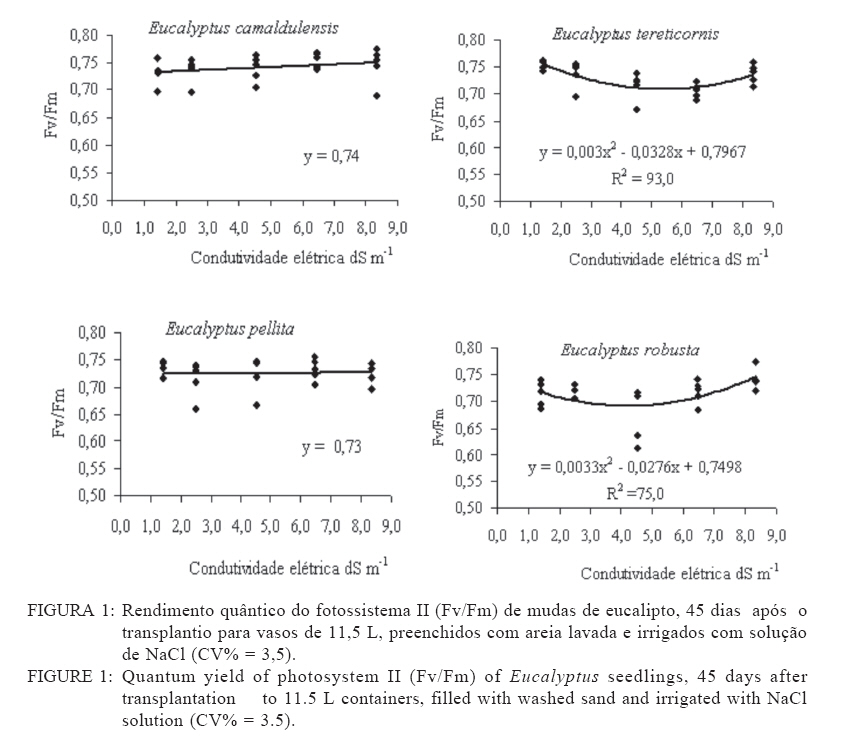
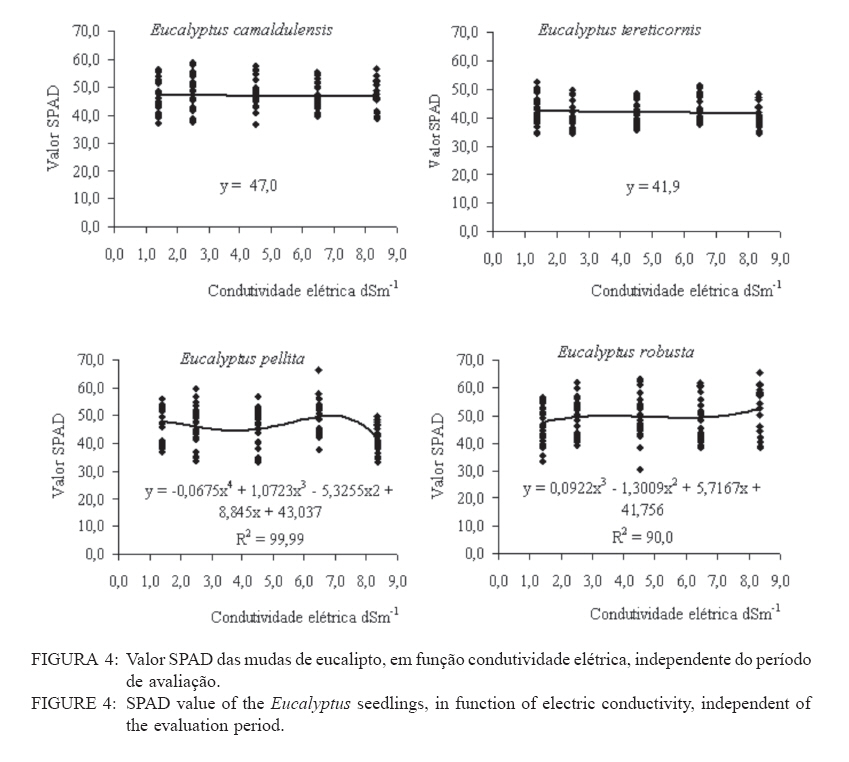
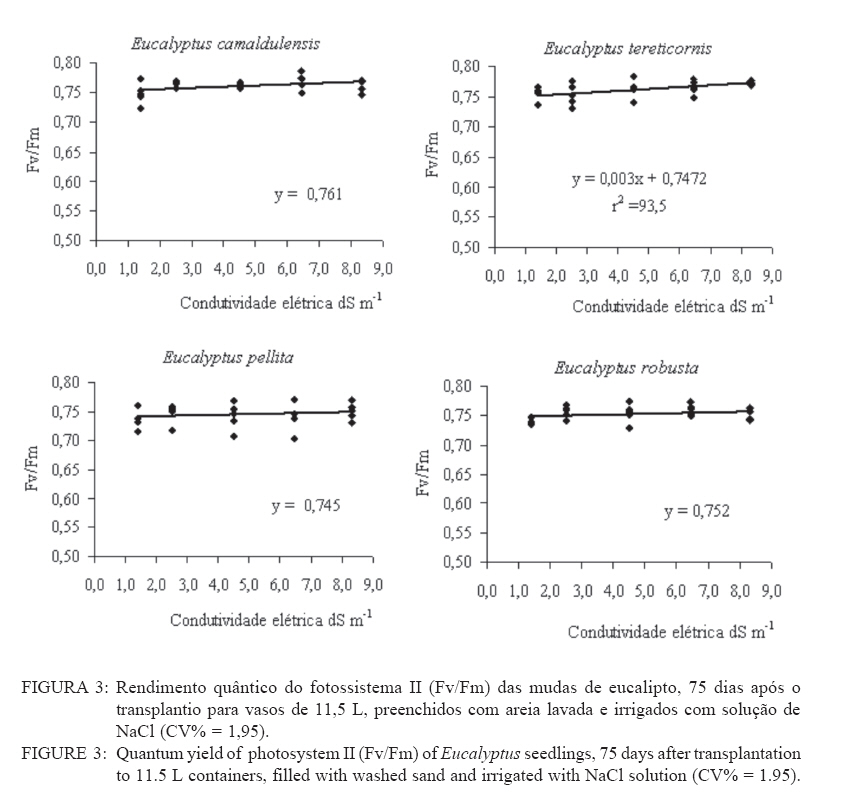
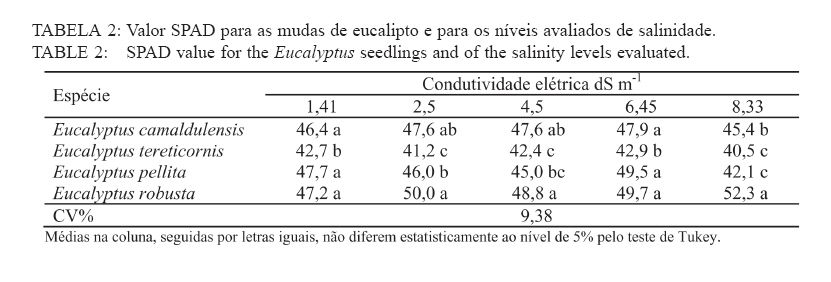
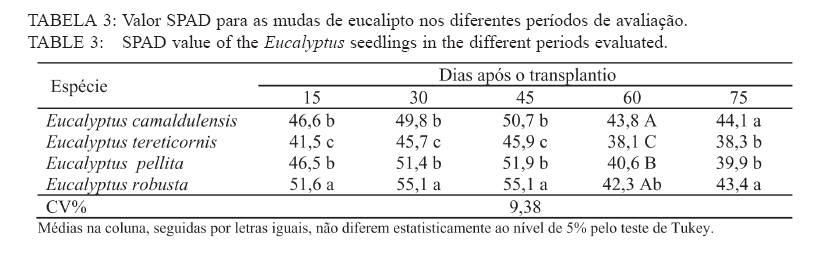
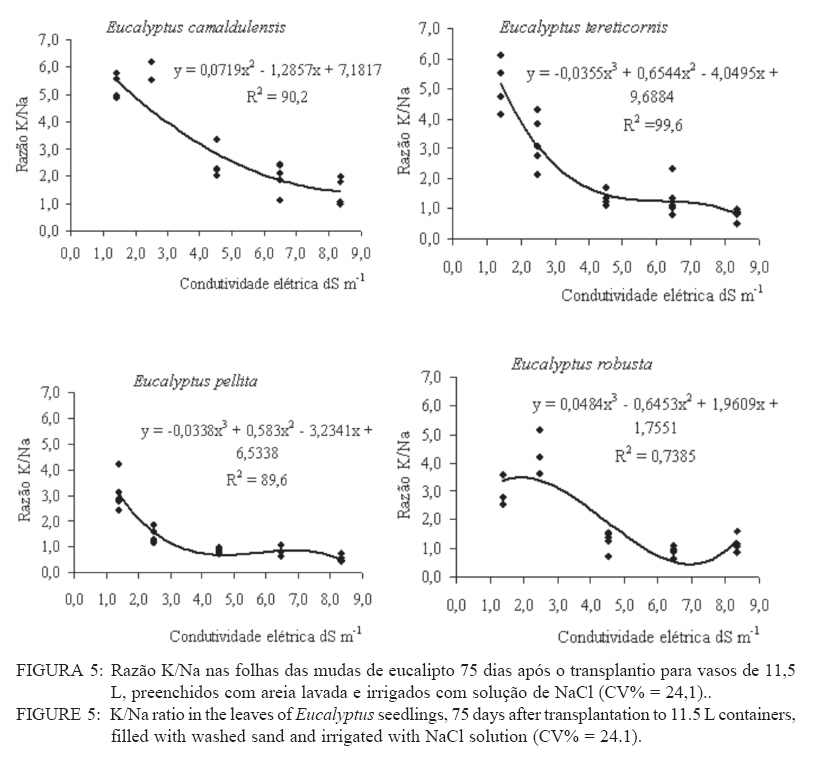
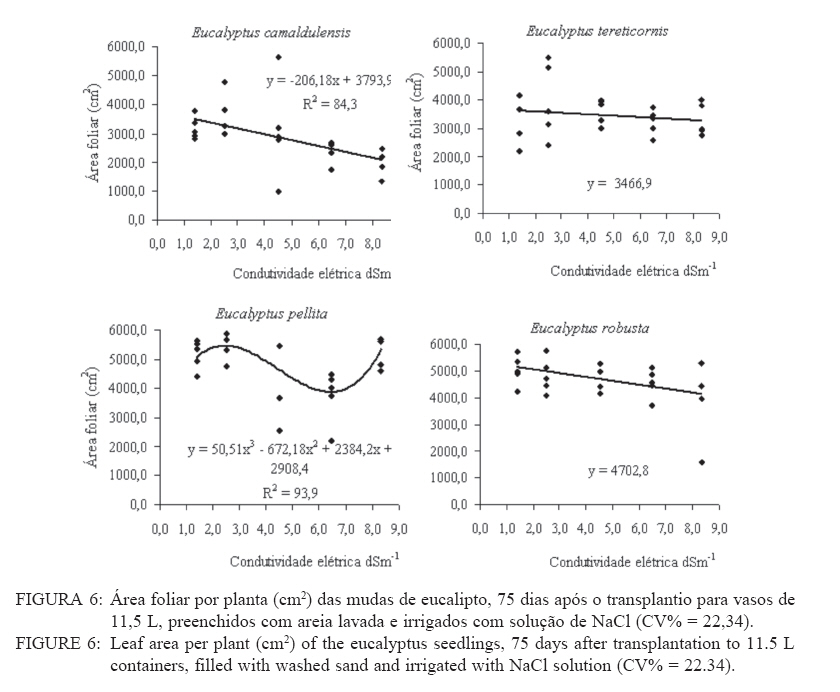
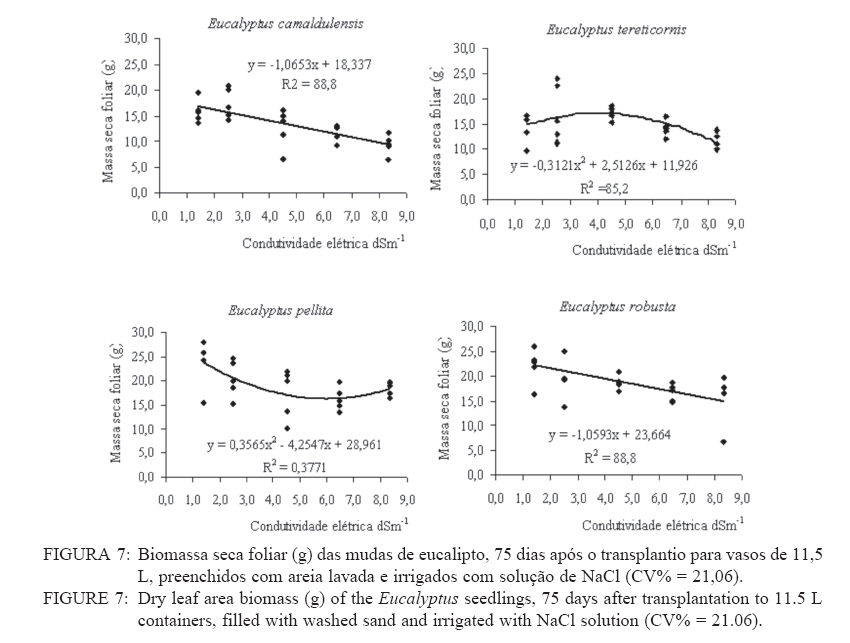
Recebido para publicação em 11/08/2008 e aceito em 12/04/2010. Code Number: cf10021 RESUMO Este trabalho objetivou avaliar como o aumento da concentração de cloreto de sódio (NaCl) influencia o rendimento quântico potencial do fotossistema II (Fv/Fm), índice da intensidade da cor verde nas folhas (valor SPAD), área foliar, biomassa seca foliar e concentração de potássio (K) e sódio (Na) nas folhas de mudas de espécies de Eucalyptus. O experimento foi conduzido em casa de vegetação, em vasos de 11,5 L, preenchidos com areia e irrigados com solução salinizada, em Campos dos Goytacazes, RJ. O delineamento experimental foi inteiramente casualizado em esquema fatorial com quatro espécies (Eucalyptus camaldulensis, Eucalyptus tereticornis, Eucalyptus pellita e Eucalyptus robusta) e cinco níveis de salinidade (1,4; 2,50; 4,50; 6,45 e 8,33 dS m-1), com cinco repetições. Até o nível máximo de salinidade aplicado, as mudas de Eucalyptus camaldulensis, Eucalyptus tereticornis e Eucalyptus robusta responderam positivamente com relação às características valor SPAD, rendimento quântico potencial do fotossistema II, biomassa seca foliar e área foliar. Essas avaliações sugerem tolerância dessas espécies às condições de salinidade impostas. Mudas de Eucalyptus pellita mostraram sensibilidade ao estresse salino. As reduções da biomassa seca e área foliar podem estar relacionadas a estratégias de resistência das plantas ao estresse salino. Palavras-chave: salinidade; fluorescência. ABSTRACTThe objective of this study was to evaluate the effects of increasing sodium chloride (NaCl) concentrations on quantum potential efficiency of the photosystem II (Fv/Fm), index of the intensity of the green color in leaves (SPAD value), dry leaf biomass, leaf area and potassium (K) and sodium (Na) concentrations in Eucalyptus seedlings leaves. The experiment was carried out in a green house, using 11.5 L pots filled with sand and irrigated with nutrient solution, in Campos dos Goytacazes (RJ). The experiment was set up according to a completely randomized factorial design, with four species (Eucalyptus camaldulensis, Eucalyptus tereticornis, Eucalyptus pellita and Eucalyptus robusta) and five saline levels (1.41; 2.50; 4.50; 6.45 and 8.33 dS m-1), with five replicates. Up to the maximum applied saline level, seedlings of Eucalyptus camaldulensis, Eucalyptus tereticornis and Eucalyptus robusta showed positive responses regarding SPAD values and quantum potential efficiency of the photosystem II, dry leaf biomass and leaf area. These evaluations suggest tolerance of these species to saline conditions. Seedlings of Eucalyptus pellita showed sensitivity to saline stress. The reductions of dry leaf biomass and leaf area may be related to seedling resistance to saline conditions. Keywords: saline conditions; chlorophyll fluorescence. INTRODUÇÃOO sucesso da utilização de espécies vegetais para produção de produtos madeireiros e não madeireiros ou para plantios de recuperação de áreas degradadas em solos salinos, depende da tolerância das mesmas à salinidade. Características, diretamente relacionadas à fotossíntese, fluorescência da clorofila e teor de clorofila têm sido utilizadas para identificar diferenças em respostas fisiológicas de espécies ou cultivares sob estresse salino por diversos autores (JAMIL et al., 2007; JIANG et al., 2006; LEE et al., 2004; LU et al., 2003; LU et al., 2002; WANG et al., 2002; MISRA et al., 2001; RAWAT e BANERJEE, 1998). Os índices relacionados à fluorescência da clorofila fornecem uma indicação da capacidade fotossintética da planta, o que é uma informação importante em estudos de estresse. De acordo com Ball et al. (1994), o funcionamento do fotossistema II (PSII) é uma característica sensível para identificar estresse em plantas. Os fatores de estresse também estão relacionados à degradação da clorofila (HENDRY e PRICE, 1993), segundo Jamil et al. (2007), quando plantas crescem sob condições de salinidade, sua atividade fotossintética diminui levando a um crescimento reduzido, menor área foliar e menor conteúdo de clorofila. Considerando que o teor de clorofila é uma característica fortemente correlacionada com valor SPAD (índice da intensidade da cor verde), é que a determinação desse valor é um procedimento rápido e não destrutivo, ele tem potencial para ser utilizado como indicador de estresse. Algumas espécies do gênero Eucalyptus têm sido consideradas tolerantes à salinidade. Estudos realizados por Su et al. (2005) e Marcar (1993) comprovam a tolerância de Eucalyptus camaldulensis à salinidade. O Eucalyptus tereticornis também tem sido apontado como tolerante (TOMAR et al., 2003 e MARCAR, 1993) e mostrou-se eficiente em melhorar as condições físicas e químicas de um solo sódico na Índia (MISHRA et al., 2003). Em relação a mudas de Eucalyptus robusta na literatura não há um consenso com relação ao seu grau de tolerância à salinidade. Sun e Dickinson (1993) consideram essa espécie altamente tolerante, Marcar (1993), como moderadamente tolerante, e Greenwood et al. (1994) de baixa tolerância. Para mudas de Eucalyptus pellita há pouca informação quanto ao seu grau de tolerância ao estresse salino, mas Sun e Dickinson (1993) verificam que essa espécie se mostrou moderadamente tolerante à salinidade. Embora, estudos tenham evidenciado a tolerância à salinidade de Eucalyptus camaldulensis, Eucalyptus tereticornis, Eucalyptus robusta e Eucalyptus pellita, poucos estudos avaliaram respostas fisiológicas dessas espécies a diferentes níveis de salinidade e fases de vida da planta. A resistência a fatores de estresse, a exemplo da salinidade, deve ser avaliada nas diferentes fases de vida da planta, uma vez que plantas tolerantes, a determinado fator de estresse, na fase adulta, não são necessariamente tolerantes na fase de muda. Este trabalho objetivou avaliar o efeito da salinidade sobre características fisiológicas: rendimento quântico potencial do fotossistema II, índice da intensidade da cor verde nas folhas (valor SPAD), área foliar e biomassa seca foliar de quatro espécies de Eucalyptus, na fase de mudas. MATERIAL E MÉTODOSO experimento foi conduzido no período de maio a novembro de 2004, em casa de vegetação da Universidade Estadual do Norte Fluminense Darcy Ribeiro (UENF), localizada no município de Campos dos Goytacazes, RJ. O delineamento experimental foi inteiramente casualizado em esquema fatorial com quatro espécies (Eucalyptus camaldulensis, Eucalyptus tereticornis, Eucalyptus pellita e Eucalyptus robusta) e cinco níveis de salinidade (1,41; 2,50; 4,50; 6,45 e 8,33 dS m-1), com cinco repetições. Cada repetição foi composta por uma muda. As mudas foram produzidas via sementes em tubetes de 50 cm3, preenchidos com substrato Plantimax®, enriquecidos com osmocote 14-14-14 (6 g kg-1). As mudas foram transplantadas para vasos com capacidade de 11,5 L, preenchidos com areia de rio (substrato). Antes do transplantio, os vasos com substrato foram lavados com 100 L de água corrente, fornecida pela empresa de tratamento de água da localidade e, subsequentemente, com 10 L de água desmineralizada. Após secagem do substrato, as mudas foram transplantadas dos tubetes para os vasos. No momento do transplantio, foi adicionado 1,8 L de solução nutritiva a cada vaso (BOLLES JONES, 1954), quantidade suficiente para atingir, aproximadamente, 60% da capacidade de campo. Para estimar a capacidade de campo cinco amostras do substrato foram secas em estufa à 1050C, até peso constante. Em seguida quantificou-se o volume de água necessário para saturar as mostras. Durante a condução do experimento foi realizada irrigação diária, às 8, 12 e 17 horas, com água desmineralizada, mantendo, aproximadamente, 60% da capacidade de campo. Para monitorar a capacidade de campo, um vaso de cada tratamento era pesado, verificando, assim, a quantidade de água necessária para irrigação. À solução nutritiva foi adicionado NaCl, em diferentes quantidades para obtenção de distintos níveis de salinidade (Tabela 1). A medida da fluorescência da clorofila a foi feita na primeira folha expandida do terceiro ramo partindo do ápice das plantas realizada por meio de fluorímetro portátil (MINIPAM, Effeltrich, Germany). Os discos foliares foram protegidos da luz, por meio de pinças de adaptação ao escuro, durante 30 minutos, antes da leitura. Após a adaptação ao escuro, foram medidos os valores de fluorescência inicial (Fo) e máxima (Fm) e com essas variáveis foi obtido o rendimento quântico potencial do fotossistema II (Fv/Fm), em que Fv é a fluorescência variável (Fv = Fm – Fo). Essa avaliação foi realizada aos 15, 30, 45, 60 e 75 dias após a aplicação dos tratamentos. O valor SPAD foi mensurado na primeira folha expandida do terceiro ramo partindo do ápice das plantas, por meio do medidor portátil de clorofila, SPAD-502, sendo obtido um valor médio proveniente da medição de três pontos na folha. Essa avaliação foi realizada aos 15, 30, 45, 60 e 75 dias após a aplicação dos tratamentos. Aos 75 dias, as plantas foram retiradas dos vasos, a parte aérea da planta foi separada em folhas e caule. A área foliar foi mensurada com um planímetro eletrônico de bancada (LI-3000, LI-COR Inc.) e posteriormente, as folhas foram secas por 48 h a 72ºC. O teor de Na e K nas folhas foi determinado (MALAVOLTA et al., 1997; JONES JÚNIOR et al., 1991). Os dados foram submetidos à análise de variância (a = 0,05), posteriormente ao teste Tukey de médias (tratamentos de natureza qualitativa) e regressão seqüencial (tratamentos de natureza quantitativa). Para a característica SPAD, avaliada no tempo, foi realizada uma análise de variância para parcelas divididas no tempo, considerando que o tempo foi um tratamento não passível de aleatorização (BANZATTO e KRONKA, 1992). Para a característica Fv/Fm também se realizou, a princípio, análise de variância para parcelas divididas no tempo. Entretanto, ao proceder os testes de normalidade e homocedasticidade, detectou-se não normalidade de resíduos e heterocedasticidade. Mesmo utilizando técnicas de transformação de dados, conforme Snedecon e Cocharan (1989) os pressupostos não foram atendidos. A opção foi realizar uma análise para cada período de avaliação, garantindo assim o atendimento dos pressupostos da análise de variância. RESULTADOS E DISCUSSÃOPara os períodos de 15 e 30 dias não houve efeito significativo da salinidade sobre o Fv/Fm para nenhuma das espécies testadas. Já aos 45 dias (Figura 1) e 60 dias (Figura 2) houve efeito significativo da salinidade sobre Fv/Fm para o Eucalyptus tereticornis e Eucalyptus robusta. Observou-se um efeito quadrático para mudas de Eucalyptus terticornis e Eucalyptus robusta aos 45 dias, havendo redução do Fv/Fm até a condutividade elétrica de 5,45 dS m-1 e 4,18 dS m-1 respectivamente. Após atingir os pontos de mínimo, constatou-se aumento de Fv/Fm com a elevação do nível de salinidade (Figura 1). Para o período de 60 dias, o efeito foi linear para essas duas espécies, com aumento de Fv/Fm, em resposta ao aumento da salinidade (Figura 2). Geralmente em plantas submetidas a estresse salino, observa-se redução no rendimento quântico do fotossistema II (Fv/Fm), conforme observado em plantas de Paspalum vaginatum (LEE et al., 2004) e Brassica juncea (MISRA et al., 2001). Também é esperado que a salinidade reduza o teor de clorofila nos tecidos fotossintetizantes, em virtude da degradação desse pigmento. Segundo Svitsev et al. (1973) esse efeito pode estar relacionado ao aumento da atividade da clorofilase, enzima envolvida na remoção da calda fitol durante a degradação da clorofila. Entretanto, não foi verificada neste estudo redução no rendimento quântico do fotossistema II (Fv/Fm) e do valor SPAD, exceto para mudas de Eucalyptus pellita que, ao contrário das demais espécies apresenta redução do valor SPAD, partindo de 6,6 dS m-1 de condutividade elétrica (Figura 4). No período de 75 dias, apenas as mudas de Eucalyptus tereticornis responderam ao tratamento salino, sendo detectado um aumento de Fv/Fm com a elevação da condutividade elétrica da solução aplicada (Figura 3). O aumento da salinização não afetou o valor SPAD das mudas de Eucalyptus camaldulensis e Eucalyptus tereticornis, enquanto as de Eucalyptus pellita e Eucalyptus robusta apresentaram relações lineares de quarto e terceiro graus respectivamente (Figura 4). As mudas de Eucalyptus robusta tiveram aumento do valor SPAD estimulado partindo da condutividade elétrica de 5,9 dS m-1. O valor SPAD do Eucalyptus pellita apresentou um ponto de máximo na condutividade elétrica de 6,6 dS m-1 tanto acima como abaixo dessa condutividade, o valor SPAD tendeu a ser reduzido. Semelhantemente ao observado para mudas de Eucalyptus pellita, a redução do teor de clorofila, mediante o aumento da concentração de NaCl, foi relatado para mudas de Eucalyptus camaldulensis e de Dalbergia sissoo, partindo da condutividade elétrica de 5 dS m-1 (RAWAT e BANERJEE, 1998) e para mudas de Sesbania rostrata e Phaseolus vulgaris, na concentração de NaCl a partir de 50mM (JUNGKLANG et al., 2003). As mudas de Eucalyptus camaldulensis mantiveram a eficiência quântica do fotossistema II e o valor SPAD inalterado com a elevação da concentração de sal. Os valores SPAD também se mantiveram inalterados em Eucalyptus tereticornis, mas a eficiência quântica do fotossistema II apresentou tendência de aumento com o aumento da salinidade, especialmente nas duas últimas avaliações, aos 60 e 75 dias. Para as mudas de Eucalyptus robusta a eficiência quântica do fotossistema II e o valor SPAD também responderam positivamente ao aumento do nível de salinidade (Figuras 1, 2, 3 e 4). Os resultados nos quais a eficiência quântica do fotossistema II manteve-se inalterada com o aumento da salinidade são compatíveis com os de outros estudos, com diferentes espécies e níveis de salinidade. Nestes, o índice Fv/ Fm também não foi alterado mediante o aumento da concentração de sal para mudas de Sueda salsa (LU et al., 2002), Sesbania rostrata (JUNGKLANG et al., 2003) e de Gossypium hirsutum L. (BRUGNOLI e BJORKMAN, 1992 e BRUGNOLI e LAUTERI, 1991). O menor valor da característica valor SPAD, em todas as concentrações de NaCl testadas e em todos os períodos de avaliação, foi observado para mudas de Eucalyptus tereticornis, em relação às demais espécies (Tabela 2 e 3). Wang et al. (2002), ao trabalhar com Pennisetum purpureum e seis níveis de concentração de sal na solução de irrigação (1,5; 5; 10; 15; 20 e 25 dS.m-1) mostraram valores decrescentes da característica valor SPAD até 15 dS m-1 de condutividade elétrica. Constataram, partindo desse ponto, elevação do valor SPAD com o aumento da concentração de sal na solução, semelhante ao comportamento das mudas de Eucalyptus robusta avaliadas neste estudo. Tais resultados podem estar associados a um estímulo do aumento do teor de clorofila em resposta à salinidade, tal suposição carece de investigação para sua fundamentação. Constatou-se redução da razão K/Na com a elevação do nível de salinidade nas folhas das mudas das quatro espécies em estudo (Figura 5). Para as quatro espécies estudadas, houve redução do teor de K e aumento do teor de Na em resposta ao aumento da concentração de NaCl, havendo redução na razão K/Na. Esse comportamento geralmente é encontrado, independentemente da espécie (JUNCKLANG et al., 2003 e FERREIRA et al., 2001). Isso se deve ao fato do Na ser competidor do K, pois o mecanismo de absorção desses elementos é semelhante (WATAB et al., 1991). Conforme se pode observar na Figura 5 e na Tabela 4, o Eucalyptus pellita apresenta a menor razão K/Na o que pode estar relacionada a menor resistência dessa espécie à salinidade, uma vez que uns dos mecanismos das plantas halotolerantes é a restrição da entrada do Na na raiz e ou do transporte para as folhas (O‘LEARY, 1995). O aumento da concentração de Na em detrimento da concentração de K pode resultar em aumento do potencial de soluto no vacúolo que gera maior turgescência e, conseqüentemente, maior expansão foliar. Nas mudas de Eucalyptus pellita ocorreu uma substituição mais intensa do K pelo Na (Figura 5), o que pode ter promovido maior expansão foliar nos maiores níveis de salinidade. Milford et al. (1977) também observaram um aumento na área foliar em plantas de beterraba com o aumento da proporção de substituição do K pelo Na. Segundo Marschner (1995), a maior taxa de crescimento foliar proporcionado pela maior substituição do K pelo Na não pode ser atribuído à maior eficiência fotoquímica e, sim, ao menor potencial osmótico proporcionado pelo K. Com relação ao efeito da salinidade, as mudas de Eucalyptus camaldulensis e Eucalyptus pellita responderam ao aumento da condutividade elétrica, as outras não foram influenciadas por esse fator. As de Eucalyptus camaldulensis apresentaram redução linear da área foliar em resposta ao aumento da concentração de NaCl. A área foliar das mudas de Eucalyptus pellita apresentaram um comportamento cúbico, com máxima e mínima área foliar nas condutividades elétricas de 2,45 e 6,42 dS m-1 respectivamente (Figura 6). Evidenciando que as espécies respondem de maneira diferenciada ao estresse salino, as mudas de Eucalyptus camaldulensis apresentaram decréscimo de área foliar, com a elevação do nível de salinidade. Tendência semelhante foi observada em mudas de Annona muricata (CAVALCANTE et al., 2001), Bactris gasipaes (FERNANDES et al., 2003) e de Salvadora oleoides (RAMOLIYA e PANDEY, 2002). A redução da área foliar pode ser considerada negativa, por diminuir a área de interceptação de luz e, conseqüentemente, prejudicar a fotossíntese. Entretanto, uma menor área foliar pode implicar em menor transpiração, o que aumenta a eficiência no uso da água e, em condições salinas, minimiza o acúmulo de sais na planta e o aumento da concentração de sais na rizosfera. Portanto, a redução da área foliar pode contribuir para o melhor desempenho da planta em ambiente salino. A biomassa seca foliar média foi significativamente maior para as mudas de Eucalyptus pellita e Eucalyptus robusta, independentemente do nível de condutividade elétrica. As de Eucalyptus camaldulensis e Eucalyptus robusta apresentaram uma redução linear da biomassa seca foliar com o aumento do nível de salinidade, enquanto que as mudas de Eucalyptus tereticornis e o Eucalyptus pellita apresentaram uma relação quadrática (Figura 7). As mudas de Eucalyptus tereticornis apresentaram valores crescentes de biomassa seca foliar até atingir um máximo de 4,03 dS m-1 mas, partindo desse ponto, constataram decréscimo dos valores. Nas mudas de Eucalyptus pellita, ao contrário, houve redução de biomassa seca foliar até 5,97 dSm-1, e partindo desse ponto de mínimo, observaram-se valores crescentes. O aumento da biomassa seca foliar propiciada pelo aumento da salinidade, observada nas mudas de Eucalyptus pellita, a partir de 5,97 dS m-1 de condutividade elétrica, pode estar relacionado ao acúmulo de sacarose na folha. Corroborando os resultados observados pela espécie citada, Poljakoff-Mayber e Lerner (1994) apresentam exemplos em que se observaram, em diferentes culturas, decréscimos da exportação de fotoassimilados das folhas para outros órgãos da planta, em resposta ao estresse salino, resultando no acúmulo de amido nas folhas, o que pode causar decréscimo na fotossíntese. O aumento da concentração de sal não afetou a área foliar das mudas de Eucalyptus tereticornis e Eucalyptus robusta, mas causou redução desta característica para as de Eucalyptus camaldulensis. Constatou-se que o aumento da salinidade proporcionou redução da biomassa seca foliar das mudas de Eucalyptus camaldulensis, Eucalyptus tereticornis e Eucalyptus robusta. Mudas de Eucalyptus pellita, ao contrário das outras espécies estudadas, apresentaram tendência de aumento de área foliar e biomassa foliar em resposta ao aumento da concentração de NaCl. É importante ressaltar que o nível de salinidade em que se inicia a redução do valor SPAD coincide, aproximadamente, com o nível onde aumenta a área foliar. Isso pode ser uma evidência de que, ao aumentar a área foliar, o teor de clorofila por cm2 de folha diminui. Esses resultados levaram a supor que, em condições de elevada salinidade, a maior área e biomassa seca foliar estão relacionadas ao menor rendimento quântico do fotossistema II (Fv/Fm) e menor concentração de clorofila nas folhas. CONCLUSÕESAté o nível máximo de salinidade aplicado, 8,33 dS m-1 de condutividade elétrica, as mudas de Eucalyptus camaldulensis, Eucalyptus tereticornis e Eucalyptus robusta responderam positivamente com relação as características valor SPAD e rendimento quântico potencial do fotossistema II, o que pode ser um indicativo de tolerância destas espécies à salinidade. As reduções da biomassa seca e área foliar estão relacionadas a estratégias de resistência das plantas ao estresse salino. REFERÊNCIAS BIBLIOGRÁFICAS
Copyright 2010 - Ciência Florestal The following images related to this document are available:Photo images[cf10021f3.jpg] [cf10021t3.jpg] [cf10021f7.jpg] [cf10021f6.jpg] [cf10021t4.jpg] [cf10021f1.jpg] [cf10021f5.jpg] [cf10021t2.jpg] [cf10021f4.jpg] [cf10021f2.jpg] [cf10021t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}