 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Ciência Florestal, Vol. 20, No. 2, Apr-Jun, 2010, pp. 269-281 CONTRIBUIÇÃO DA VEGETAÇÃO RASTEIRA NA EVAPOTRANSPIRAÇÃO TOTAL EM DIFERENTES ECOSSISTEMAS DO BIOMA CERRADO, DISTRITO FEDERAL CONTRIBUTION OF UNDERSTORY VEGETATION ON TOTAL EVAPOTRANSPIRATION IN DISTINCT ENVIRONMENTS OF THE CERRADO BIOME, DISTRITO FEDERAL, BRAZIL Otacílio Antunes Santana1 Geraldine Cuniat2 José Imaña-Encinas3
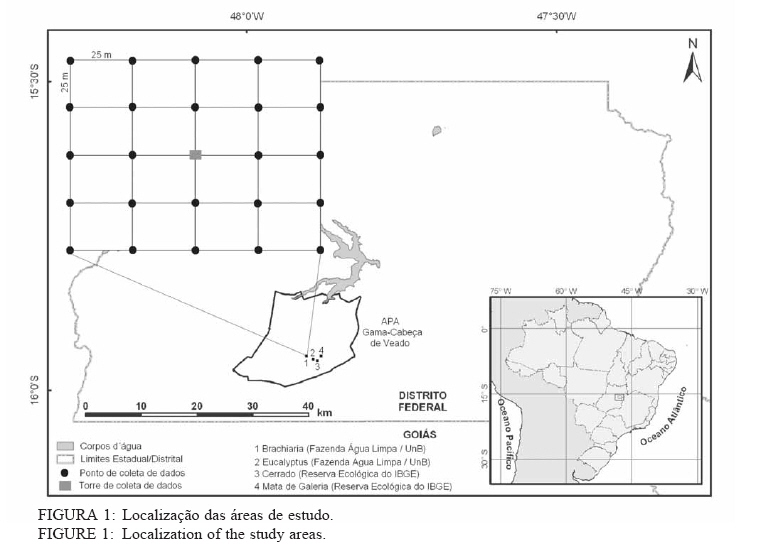
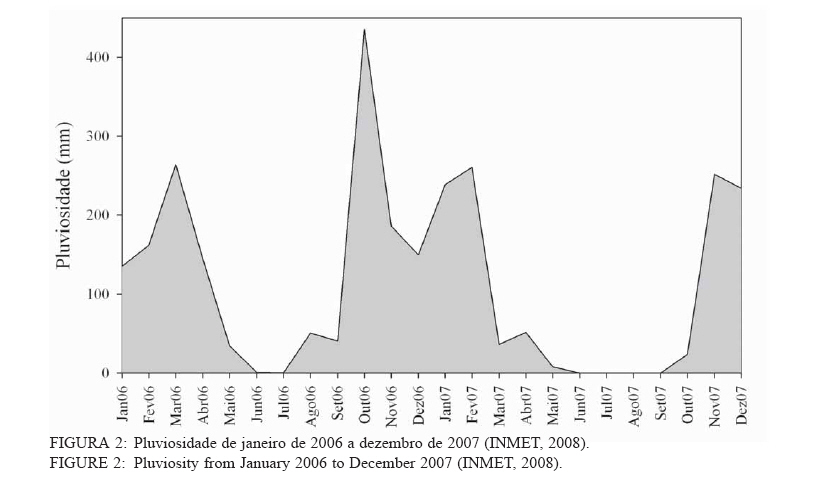
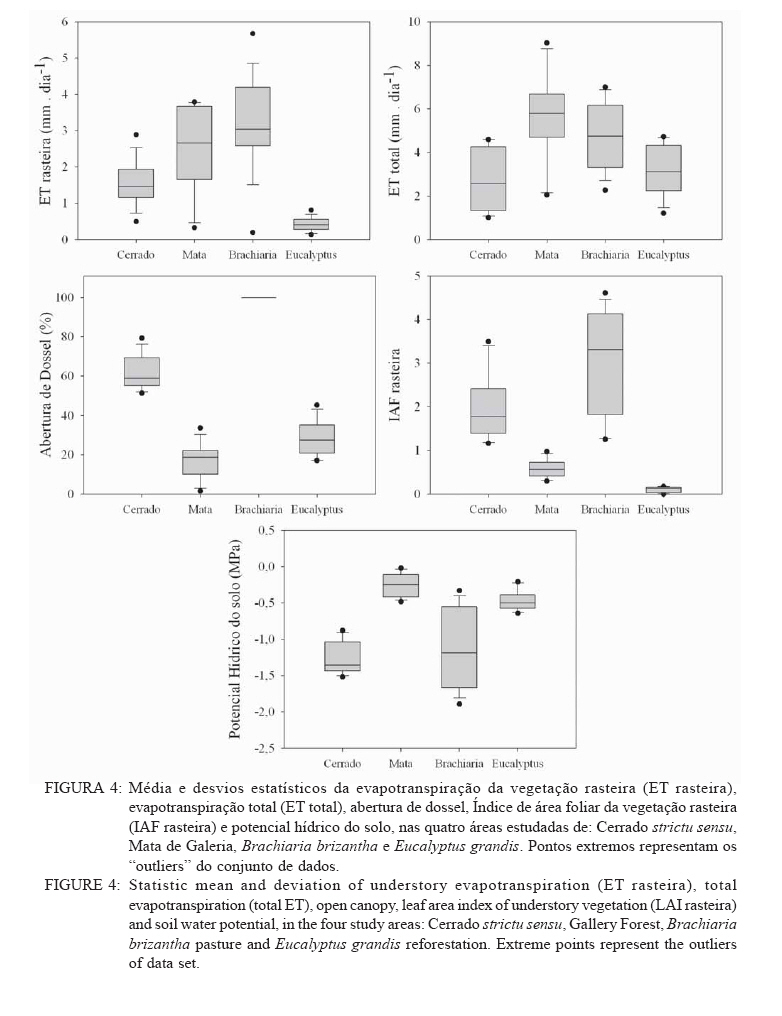
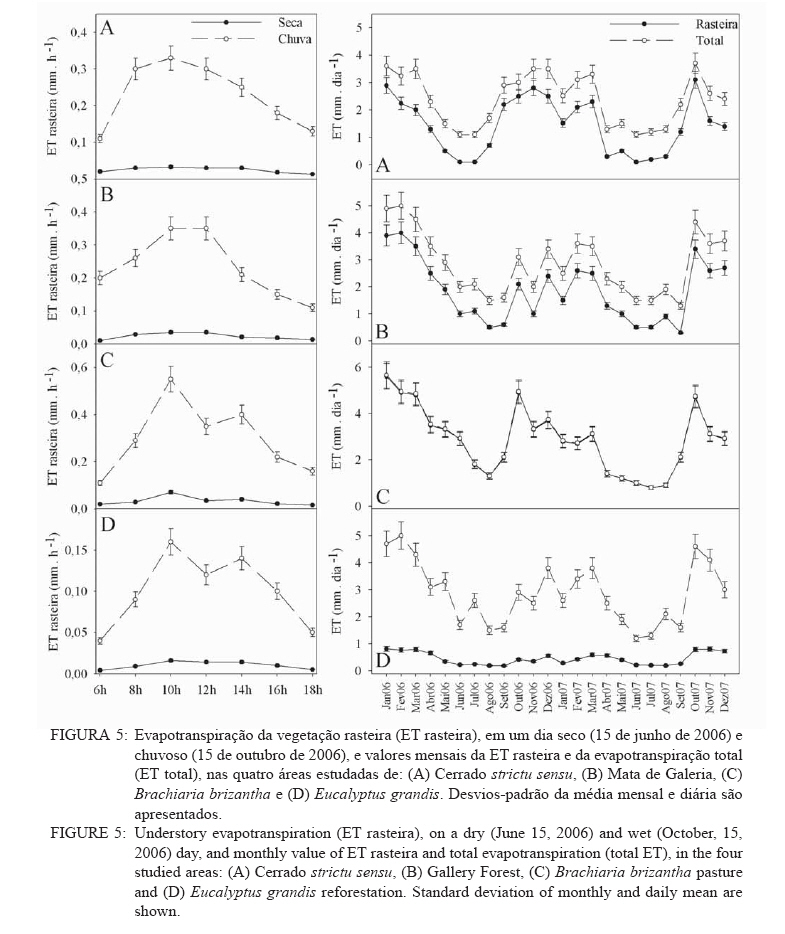
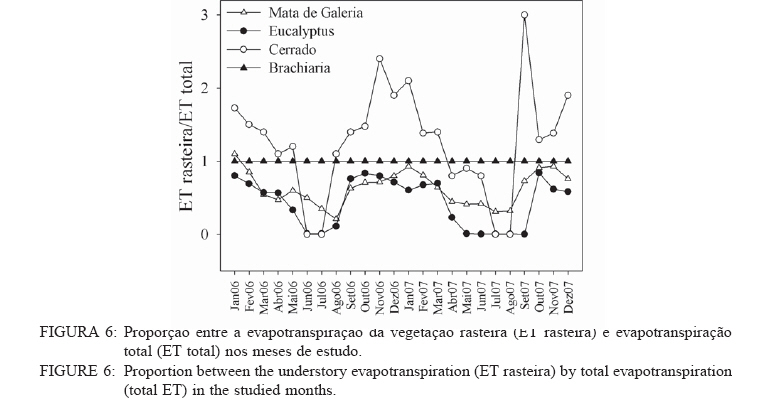
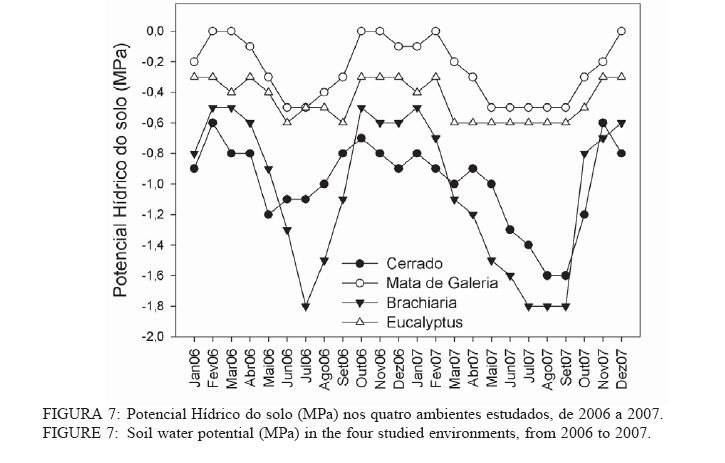
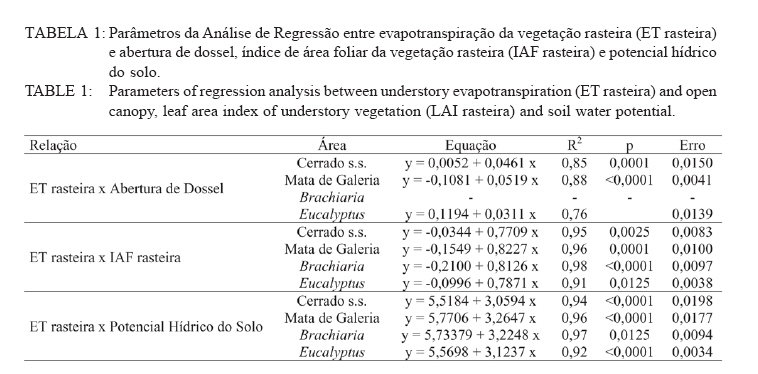
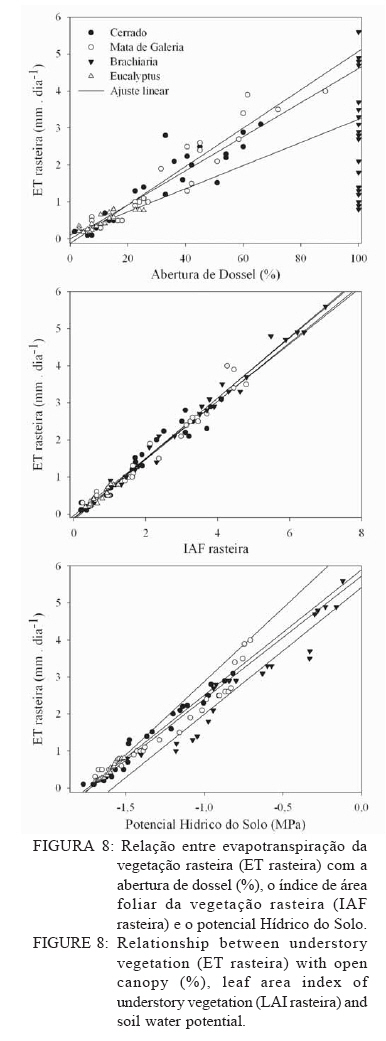
Recebido para publicação em 11/08/2008 e aceito em 26/04/2010. Code Number: cf10022 RESUMOOs objetivos deste trabalho foram: (a) quantificar sazonalmente a evapotranspiração da vegetação rasteira (ET rasteira), (b) quantificar a influencia da ET rasteira para a evapotranspiração total (ET total), e (c) relacionar a ET rasteira com o índice de área foliar da vegetação rasteira (IAF rasteira), abertura de dossel e potencial hídrico do solo; em quatro diferentes paisagens: Cerrado stricu sensu, Mata de Galeria, pastagem e reflorestamento, situados na APA Gama-Cabeça de Veado, Distrito Federal. Câmara de medição de evapotranspiração para vegetação rasteira; analisador de gás infravermelho não dispersivo; analisador de abertura de dossel e índice de área foliar; e psicrômetros de solo, foram utilizados para coleta de dados. A Evapotranspiração da vegetação rasteira (ET rasteira) contribuiu de forma significativa para a Evapotranspiração Total (ET total), sobretudo no período chuvoso, chegando a representar cerca de 60 % na média anual de ET total, em áreas de Cerrado strictu sensu e em Mata de Galeria. Uma significativa e diretamente proporcional relação entre ET rasteira com as variáveis IAF rasteira, abertura total e potencial hídrico do solo foi observada, influenciada pela sazonalidade das chuvas. Palavras-chave: sazonalidade; ciclo hidrológico; IAF; abertura de dossel. ABSTRACTThe objectives this study were to: (a) quantify the seasonality evapotranspiration of understory vegetation (ET rasteira), (b) quantify the influence of ET rasteira on total evapotranspiration (total ET) and (c) verify the relation between ET rasteira and Leaf Area Index of the understory vegetation (LAI rasteira), open canopy and soil water potential, in four distinct landscapes: Cerrado stricu sensu, Gallery Forest, Brachiaria pasture and Eucalyptus reforestation, situated in the APA Gama-Cabeça de Veado, Districto Federal. An open-top chambers for understory vegetation, non-dispersive infrared gas analyzer, plant canopy and leaf area index analyzer; and soil psychrometers were used to collect data. The ET rasteira contributed significantly to total ET, mainly during rainy season, approximately 60 % of the annual mean of total ET, in Cerrado strictu sensu and Gallery Forest areas. Significant and directly proportional relationships between ET rasteira and the understory LAI, open canopy and soil water potential were observed, this influenced by rain seasonality. Keywords: seasonality; hidrologic cicle; LAI; open canopy. INTRODUÇÃOApesar de a água ser a substância mais abundante sobre a costa terrestre, o déficit hídrico é o principal fator limitante da produtividade primária em escala global (JELTSCH et al., 2000). Aproximadamente metade das comunidades vegetais do mundo passa por extensos períodos secos (MEINZER, 2003). Até comunidades ombrófilas, como as florestas tropicais úmidas, podem passar por déficits hídricos moderados durante o dia, e experimentam, ocasionalmente, restrições de água mais severas (MALHI et al., 2002; WOODRUFF et al., 2007). A circulação hídrica proveniente das plantas é influenciada tanto por fatores climáticos (precipitação total e a distribuição sazonal das chuvas) como edáficos (EAMUS et al., 2000). A cobertura vegetal também representa um papel-chave nesse regime hídrico, em especial no que se refere ao fluxo de água pelo sistema solo-atmosfera, que atua interceptando e redistribuindo a precipitação, aumentando a infiltração e levando a uma redução na taxa de evaporação da superfície do solo (HUTLEY et al., 2001). Padrões de uso de água no solo por herbáceas e lenhosas são reconhecidos como sendo os principais determinantes da interação entre esses dois estratos. Os dois componentes vegetacionais formam camadas no solo com independentes balanços hídricos. As herbáceas adquirem água da camada mais superficial do solo, onde suas raízes são dominantes, e as lenhosas têm acesso a camadas mais profundas, mas são competitivamente inferiores nas camadas superficiais do solo (HOFFMANN et al., 2004; CAYLOR et al., 2005; SCHOLZ et al., 2007). Os ecossistemas savânicos se caracterizam por possuir um estrato rasteiro que coexiste com arbustos lenhosos e árvores perenifólias ou decíduas. No cerrado, apesar da sazonalidade nas precipitações, as árvores transpiram entre as estações, pela obtenção de água em grandes profundidades por suas raízes, diferindo das gramíneas e outras plantas herbáceas que possuem raízes superficiais (SCHOLZ et al., 2007). Alguns autores descrevem que existe uma regulação do fluxo de água pelas plantas lenhosas durante a estação seca com uma importação de água do solo, uma redução na área foliar, e uma consequente diminuição na taxa fotossintética e de condutância estomática (EAMUS et al., 2000; BUCCI et al., 2004a; HOFFMANN et al., 2005). Enquanto o componente lenhoso da vegetação do cerrado é capaz de manter o fluxo de água durante a seca, apesar de reduzido, gramíneas e outras herbáceas evitam o estresse hídrico por meio da dessecação da parte aérea e da redução das trocas gasosas, fazendo com que sua transpiração seja mínima ou ausente (SACK e FROLE et al., 2006). No entanto, o cerrado tem sofrido transformações profundas em sua cobertura vegetal e no uso da terra, com a potencialidade de causar modificações na estrutura e funcionamento desse ecossistema (HOBBS et al., 2006). As pastagens monoculturais e reflorestamentos constituem hoje o tipo de cobertura vegetal mais comum em que o cerrado foi desmatado, representando cerca de 60% dos agrossistemas existentes no Brasil Central (KLINK e MACHADO, 2005). A vegetação rasteira, por ser tão sensível a mudança estacional e a presença de água no solo, merece atenção especial por passar no período seco uma redução e desaparecimento de sua parte aérea fotossinteticamente ativa (JELTSCH et al., 2000). Observar sua contribuição para um processo do ciclo hidrológico pode justificar uma proposta de controle da mudança no uso da terra (KLINK e MACHADO, 2005; SCHOLZ et al., 2007). Os objetivos deste trabalho em quatro distintos ambientes foram: (a) quantificar sazonalmente a evapotranspiração da vegetação rasteira (ET rasteira), (b) quantificar a influencia da ET rasteira para a evapotranspiração total, e (c) relacionar a ET da vegetação rasteira com seu índice de área foliar vegetação rasteira (IAF rasteira), abertura de dossel e potencial hídrico do solo. MATERIAIS E MÉTODOSO estudo foi realizado na Área de Proteção Ambiental Cabeça-Gama de Veado, dentro da Reserva Ecológica do IBGE (RECOR) e da Fazenda Água Limpa (FAL/UnB), áreas que fazem divisas e que se localizam na região sudoeste do Distrito Federal (Figura 1). Quatro unidades de paisagens foram escolhidas, sendo duas nativas, situadas na RECOR: Cerrado strictu sensu (Cerrado s.s.) e Mata de galeria, e duas com plantios, situados na FAL: uma pastagem com Brachiaria brizantha (Hochst. ex A. Rich.) Stapf (Poaceae) e outra com reflorestamento de Eucalyptus grandis W. Hill ex Maiden (Myrtaceae). A área de Cerrado s.s. estudada possui uma densidade arbórea (> 5 cm de diâmetro a 20 cm do solo) de 685 indivíduos por hectares (ind./ha), sendo as espécies arbóreas com maiores densidades (SANTANA, 2003): Sclerolobium paniculatum Vogel (Fabaceae); Eriotheca pubescens (Mart. & Zucc.) Schott & Endl. (Malvaceae); Qualea dichotoma (Mart.) Warm. (Vochysiaceae); Psdium warmingianum Kiaersk. (Myrtaceae); e Byrsonima verbascifolia (L.) DC. (Malphighiaceae). A vegetação rasteira foi caracterizada pela presença de indivíduos de espécies herbáceas e arbustivas, menores do que 1,3m, e diâmetros à base do tronco (20 cm do solo), para espécies arbustivas, menores que 5 cm (SANTANA, 2003). Na área de Cerrado as espécies da vegetação rasteira que tiveram maiores densidades foram: Echinolaena inflexa (Poir.) Chase (Poaceae); Croton goyazensis Müll. Arg. (Euphorbiaceae); Clitoria guianensis (Aubl.) Benth. (Fabaceae); Ichnanthus camporum Swallen (Poaceae); e Peixotoa goiana C. E. Anderson (Malphighiaceae). Na área de mata de Galeria, foi registrada uma densidade arbórea de 1.573 ind./ha (SANTANA, 2003). As espécies arbóreas que tiveram maior densidade foram: Tapirira guianensis Aubl. (Anacardiaceae); Copaifera langsdorfii Desf. (Fabaceae); Lamanonia ternata Vell. (Cunoniaceae); Anadenanthera colubrina (Vell.) Brenan var. cebil (Griseb.) Altschul (Fabaceae); e Guettarda iburnioides Cham. e Schlecht. (Rubiaceae). As espécies da vegetação rasteira que tiveram maior densidade foram: Scleria scabra Willd. (Cyperaceae); Echinolaena inflexa (Poir.) Chase (Poaceae); Cissampelos ovalifolia DC. (Menispermaceae); Axonopus barbiger (Kunth) Hitchc. (Poaceae); e Elyonorus muticus (Spreng.) O. Kuntze (Poaceae). A densidade arbórea para área estudada de Eucalyptus foi de 1.650 ind./ha, sendo que o plantio desse reflorestamento foi realizado há 12 anos, com espaçamento 3 x 2 m. No momento da coleta dos dados na pastagem de Brachiaria, área para amostragem dos dados, já havia 6 meses de seu plantio. Dentro de cada área de estudo foram estabelecidas quatro parcelas (Figura 1) de 100 x 100 m, sendo em seu centro, instalada uma torre para coleta de dados de evapotranspiração total (ET total) subdivididas em quadrados de 25 x 25 m, onde em suas extremidades foram coletados os dados de evapotranspiração da vegetação rasteira (ET rasteira). O clima das áreas de estudo é o Aw segundo a classificação de Köppen-Geiger, caracterizado pela sazonalidade do regime de chuvas: período chuvoso de outubro a abril, e período seco de maio a setembro Figura 2. As áreas estão sob o Latossolo Vermelho (LVd6), distrófico com A moderado, textura média, de relevo plano e suave-ondulado (REATTO et al., 2004). A ET rasteira em cada ponto de coleta de dados foi mensurada por meio da câmara de medidas de ET rasteira (Figura 3A), open-top chambers (HUTLEY et al., 2000). A câmara foi construída por um material transparente que não influenciava a temperatura interna, aberta em cima com a presença de um anemômetro de hélice (model LI 6262, Lincoln, NE, USA), que media a velocidade do vento que estava saindo da câmara. O vapor de água foi quantificado na base e no topo da câmara por um higrômetro (HR 33T, Ontário, Canadá), em uma média de fluxo de 8 litros min-1, coletada por uma bomba de aquário de 1,5V convencional, e sistematizada por um controlador de massa de fluxo (model FC, 280 Tylan General, Torrence, CA, USA). Foram registradas as médias a cada 1 minuto em um sistema de aquisição de dados (21X Campbell Scientific, UT, USA). A câmara ocupava e mensurava uma área de 1 m de circunferência de vegetação rasteira, englobando gramíneas, arbustos e plântulas de até 1 m de altura. Foi feita uma calibração em laboratório com um vapor de água conhecido, e obtiveram-se erros menores de 5%. ET rasteira (mm) foi calculada como segue (HUTLEY et al., 2000): ET rasteira = V·(ñsaída-ñentrada)/ A , sendo V = fluxo volumétrico do vapor de água que saia da câmara (m3·s-1) taxa; rsai e rent são a densidade de vapor em (g·m-3) de saída e entrada, respectivamente; e A é a área basal da câmara (m2). A ET rasteira foi mensurada em cada um dos pontos, e em cada área de coleta de duas em duas horas, das 6 às 18h, cinco vezes por mês, de janeiro de 2006 a dezembro de 2007, todos em dias sem chuvas independente do regime destas. A ET total, ou seja, somatório da evaporação (Capex V2X, Charles Austin, Survey, UK). Todos do solo e da transpiração da vegetação rasteira e os equipamentos foram instalados em uma torre arbórea, foi quantificada partindo de um analisador acima do dossel no centro de cada área de estudo. de gás não dispersivo infravermelho (Figura 3B), de Foi registrada, a cada 30 min, a covariância dos dados rápida resposta (6262, LiCOr Inc., Lincoln, NB, de ET total em um computador portátil. Esses dados USA) e coletado por uma fina mangueira BEV-A-também foram coletados diariamente de janeiro de Line (4 mm de diâmetro) com um filtro aerosol de 2006 a dezembro de 2007. abertura de poro de 1 µm (ACRO 50 PTFE, German, O potencial hídrico do solo foi mensurado Ann Arbor, MI, USA), em um fluxo de 15 min-1 com psicrômetros de solo (PST-55, Wescor, Logan, UT) a cada 30 minutos, e todos os dados ficavam registrados em um sistema de aquisição de dados (CR-7, Campbell Scientific, Logan, UT), localizados a uma profundidade de 1 m (Figura 3C). Os psicrômetros foram calibrados individualmente com soluções salinas de conhecidas osmolaridades seguindo o procedimento de Bucci et al. (2004b). O índice de área foliar da vegetação rasteira (IAF rasteira) e a abertura de dossel (%) foram quantificados através de um analisador de dossel LAI-2000 (LiCOR, Plant Canopy Analyzer; Figura 3D). Esses dados foram coletados exatamente no mesmo ponto de coleta da ET rasteira, sendo IAF rasteira a 20 cm e abertura de dossel a 1,5 m do solo. A razão entre ET rasteira/ET total foi efetuada para mostrar a contribuição da ET rasteira para a ET total durante todo o ano, conforme proposto por Hutley et al. (2000). Testes estatísticos não paramétricos foram efetuados para obter a diferença estatística entre dois grupos de amostras, Teste Mann-Whitney, e para mais de dois grupos de amostras, Teste Kruskal-Wallis, ambos para obtenção do nível de significância, valor de p. O teste Mann-Whitney foi aplicado aos dados entre o período de seca e chuva, e entre a ET rasteira e ET total, e o Teste Kruskal-Wallis entre as variáveis para as quatro áreas estudadas. Análise de regressão foi aplicada para obtenção dos valores do coeficiente de determinação (R2), do erro, da equação e do nível de significância do ajuste (valor de p), entre as relações da Evapotranspiração da vegetação rasteira (ET rasteira) com a abertura de dossel (%), o índice de área foliar da vegetação rasteira (IAF rasteira) e o Potencial Hídrico do Solo, nas quatro áreas de estudo. RESULTADOS E DISCUSSÃOOs maiores valores médios anuais da ET rasteira foram encontrados na área de pastagem de Brachiaria (3,23 mm·dia-1), área que também apresentou maiores desvios-padrão (1,51 mm·dia-1), seguido pela área de Mata de Galeria (2,84 mm·dia1), de Cerrado s.s. (1,52 mm·dia-1) e de Eucalyptus (0,46 mm·dia-1). Diferentemente da ET total, onde os maiores valores médios ocorreram na área de Mata (5,85 mm·dia-1), seguidos pelas áreas de Brachiaria, Eucalyptus e Cerrado s.s. (Figura 4). A sequência decrescente dos valores médios de ET rasteira das áreas estudadas (Brachiaria>Mata de Galeria>Cerrado>Eucalyptus) foi devida primeiramente pela densidade da vegetação rasteira. A maior densidade de indivíduos das espécies que formam a camada rasteira, espécies herbáceas e arbustivas, resulta em uma maior a área foliar por área, contribuindo para a transpiração vegetal (HUTLEY et al., 2001). A Mata de Galeria apresenta menor densidade de indivíduos da vegetação rasteira do que o Cerrado (SANTANA, 2003), porém teve uma maior média de ET rasteira em virtude da presença de águas livres em superfícies após precipitações, na serrapilheira e nos galhos (JELTSCH et al., 2000). Os valores de ET rasteira foram semelhantes aos dados encontrados na literatura, coletados com semelhantes metodologias: 4,01 mm·dia-1 (SANTOS et al., 2004); 2,11 mm·dia1 (BUCCI et al., 2005); 1,61mm·dia-1 (SANTANA, 2003); 0,53 mm·dia-1 (HUTLEY et al., 2000), para as áreas de Brachiaria, Mata de Galeria, Cerrado e Eucalyptus, respectivamente. A sequência decrescente dos valores médios de ET total das áreas estudadas (Mata de Galeria>Brachiaria>Eucalyptus>Cerrado) foi em razão sobretudo da densidade arbórea somados a densidade da vegetação rasteira, porém na área de Mata de Galeria, o curso d´água próximo a vegetação contribuiu com sua evaporação, para que esse ecossistema fosse o de maior valor médio de ET total (SCHOLZ et al., 2007). Os ambientes de Cerrado s.s. e Eucalyptus por não possuírem um estrato rasteiro tão intenso como a da Brachiaria e nem curso d´água como a Mata de Galeria, possuem menores valores de ET total. A área com Eucalyptus possui uma ET total maior do que no Cerrado, por sua maior densidade arbórea de 1.650 indivíduos por hectares (ind./ha) e 685 ind./ha respectivamente. Menor radiação solar incidente na vegetação rasteira em áreas de Mata de Galeria não permitiram altos valores de ET rasteira em relação a outros ambientes no período chuvoso, em consequência da mínima transpiração vegetal por causa da menor incidência solar nessa vegetação (BUCCI et al., 2004b). Os desvios nos valores de ET nas áreas foram em razão da sazonalidade do regime de pluviosidade, sendo que, nos períodos secos, tanto a ET rasteira, como a ET total reduzem bruscamente seus valores (EAMUS et al., 2000; EAMUS et al., 2001), como observado pelos valores extremos na Figura 4. Essa sazonalidade ocorre também diariamente, mostrando o fator da radiação solar durante o dia, influenciou nos valores de ET rasteira, e que essa diferença é visível tanto nas estações secas como nas úmidas valores na área com Brachiaria, fator determinante (Figura 5), também descrito por Scholz et al. (2007). para o aumento da transpiração vegetal com o A densidade da vegetação rasteira foi aumento de tecido foliar por unidade de área (Figura transcrita pelo valor de IAF rasteira, sendo os maiores 4). Segundo Santos et al. (2004), para essa pastagem, maiores formações de orvalho, no período chuvoso, e uma alta incidência de radiação solar, pela ausência de dossel do estrato arbóreo, ajudaram no aumento da ET rasteira. Efetuados os testes não paramétricos, foram observadas as diferenças significativas (p < 0,001) entre os valores de ET rasteira e ET total, e entre o período seco e chuvoso para essas duas ET. Isso confirmou os valores elevados dos desvios-padrão nas variáveis analisadas, demonstrando a influencia da sazonalidade pluviométrica, conforme descrito por Hutley et al. (2000 e 2001). Significativas (p < 0,001) também foram as diferenças entre os valores, nas quatro fisionomias estudadas, em cada variável mensurada: ET rasteira, ET total, abertura de dossel, IAF rasteira e potencial hídrico do solo, demonstrando que cada ecossistema possue uma contribuição diferenciada o balanço hídrico do sistema, dentro do ciclo hidrológico, corroborando com Bucci et al. (2005). No período estudado, para área de Cerrado e Mata de Galeria, a ET rasteira contribuiu com cerca de 60% do valor do ET total, e apenas 16% para a área de Eucalyptus (Figura 6). Para Brachiaria a ET rasteira representou integralmente a ET total, sendo que dados semelhantes nessa área, com as distintas metodologias de coleta das ETs, mostraram a eficiência e a calibração dos dados. Eamus et al. (2000 e 2001) e Hutley et al. (2000 e 2001) em savana australiana, e com metodologias semelhantes, mostraram a importância da vegetação rasteira para o ciclo hidrológico, sendo que a ET rasteira representou a maior parte da ET total anualmente, o que os dados deste trabalho corroboram. O IAF rasteira, entre as fisionomias vegetais estudadas, foi influenciado pelo regime sazonal das chuvas, nas áreas de Cerrado, conforme descrito por Simoni et al. (2003) e Scholz et al. (2007), sendo que características ecofisiológicas da vegetação de Cerrados mantêm indivíduos e espécies vivas durante a estação de seca, apesar de sua perda foliar. As espécies herbáceas desse Bioma regulam a perda hídrica pelos estômatos, entram em senescência e perdem sua parte aérea no período seco, ressurgindo posteriormente no período chuvoso, todo o processo fotossintético e de evapotranspiração vegetal (BUCCI et al., 2005). Isso também pode explicar a sazonalidade da razão entre ET rasteira e ET total, mosatrando que, durante o período seco, a ET rasteira contribui menos com a ET total do que no período chuvoso (Figura 6). Essa proporção foi constante, sempre 1, para área da Brachiaria em razão da falta de cobertura vegetal arbórea, a ET rasteira representa a ET total. Para as áreas de Eucalyptus e Mata de Galeria, a ET rasteira contribuiu menos para a ET total, observando também uma sazonalidade, sendo que, na área de Mata de Galeria não houve período de contribuição zero em virtude da disponibilidade hídrica próxima à vegetação, mantendo parte de sua vegetação rasteira realizando fotossíntese durante o período de seca, conforme observado por Santana (2003). A ET rasteira da área de Cerrado foi a que mais contribuiu para a ET total, sobretudo no período chuvoso, período em que a vegetação rasteira resurge com maiores IAF, comportamento descrito por Hoffmann et al. (2005). Os valores médios de IAF rasteira foram semelhantes aos encontrados na literatura (HUTLEY et al., 2000; SIMONI et al., 2003; SANTOS et al., 2004; HOFFMANN et al., 2005), utilizada a mesma metodologia, em todas as fisionomias vegetais estudadas, sendo 1,8 para Cerrado s.s.; 0,5 para Mata de Galeria; 3,5 Brachiaria e 0,1 para Eucalyptus. Para abertura de dossel, os valores médios amostrados para as áreas foram de 60, 20, 0%, e 25% para Cerrado s.s., Mata de Galeria, Brachiaria e Eucalyptus respectivamente também semelhantes aos valores encontrados na literatura, coletados com a mesma metodologia (HUTLEY et al., 2000; SIMONI et al., 2003; SANTOS et al., 2004; HOFFMANN et al., 2005), variando pelas estações do ano. Segundo Haol et al. (2008), outro fator que influenciou a transpiração vegetal e a evaporação do solo foi o potencial hídrico do solo, em seus estudos na região de Cerrado. Em períodos secos, há uma diminuição nesse potencial, reduzindo a água disponível no solo, especialmente nas camadas superficiais, influenciando as fisionomias vegetais com maiores densidades de vegetação rasteira em relação ao estrato arbóreo, e consequentemente, reduzindo o valor de ET rasteira nesse período. O potencial hídrico do solo registrado nas áreas de estudo (Figura 7) também mostrou um padrão sazonal, influenciado pelo regime das chuvas e uma cobertura arbórea, que diminuíam a incidência solar na superfície do solo, fazendo que houvesse uma menor desidratação do solo. Esse padrão foi observado nos resultados, onde áreas com maiores densidades arbóreas, Mata de Galeria e Eucalyptus, tiveram maiores valores médios de potencial hídrico do solo, -0,2 e -0,5 respectivamente, ou seja, quando mais próximo de zero foi o valor do potencial hídrico, mais hidratado estava o solo e menos água foi requerido do sistema (SCHOLZ et al., 2007; HAOL et al., 2008). Nas áreas de Cerrado e em áreas de Brachiaria, a pouca ou ausência densidade arbórea, faz com que o solo receba uma maior incidência solar, sobretudo nas épocas secas, em que a vegetação rasteira sofre uma dissecação, obtendo esses ambientes valores médios menores de potencial hídrico do solo (BUCCI et al., 2004a; SANTOS et al., 2004), -1,25 e -1,4 respectivamente. As relações entre ET rasteira com abertura total, IAF rasteira e potencial hídrico do solo, foram significativas (R2 > 0,76; p < 0,01; Erro < 0,02), em todos os ambientes estudados, sendo diretamente proporcional essas relações, conforme apresentados os parâmetros na Tabela 1, e a dispersão dos dados na Figura 8. Segundo Hutley et al. (2000 e 2001), Scholz et al. (2007) e Haol et al. (2008) essas relações significativas indicam a importância da manutenção da camada vegetal que se deve ter, pois uma maior área foliar disponível no ambiente, sobretudo da vegetação rasteira que contribue mais para a ET total, maior será a transpiração vegetal, participando assim de forma significativa na ciclagem hídrica. Goldstein et al. (2008) descreveram que essas relações, mostraram a importância da cobertura arbórea, representada pela abertura de dossel, para proteção contra a incidência de raios solares diretamente ao solo, mantendo o potencial hídrico do solo aproximadamente constante em épocas distintas do ano, reduzindo a temperatura superficial do solo e a perda hídrica para a atmosfera, como observado com os dados deste trabalho. CONCLUSÕES Os valores de evapotranspiração da vegetação rasteira (ET rasteira) foram próximos aos descritos na literatura, utilizadas as mesmas metodologias, com a sequência decrescente dos valores médios nos ambientes estudados Brachiaria>Mata de Galeria > Cerrado > Eucalyptus, em razão sobretudo da densidade da vegetação rasteira e da presença de águas superficiais. Desviospadrão de até 50% dos valores médios de ET rasteira foram encontrados nas áreas durante o período estudado, devido a sazonalidades registradas, tanto durante os períodos de chuva e seca ocorridas, quanto durante o dia. A ET rasteira contribuiu com cerca de 60% do valor do ET total para as áreas Cerrado e Mata de Galeria, e apenas 16% para a área de Eucalyptus. Para Brachiaria a ET rasteira representou integralmente a ET total. Isto implica a importância da vegetação rasteira para o ciclo hidrológico. As relações entre ET rasteira com abertura total, IAF rasteira e potencial hídrico do solo foram significativas em todos os ambientes estudados, sendo diretamente proporcionais essas relações, afirmando a importância da cobertura vegetal rasteira e arbórea para contribuição na transpiração vegetal e na proteção da incidência solar direta na superfície do solo. AGRADECIMENTOSÀ Maria Aparecida da Silva, pela ajuda nos trabalhos de campo e na identificação das espécies arbóreas e herbáceas. Ao CNPq/Conselho Nacional de Desenvolvimento Científico e Tecnológico e à CAPES/Coordenação de Aperfeiçoamento de Pessoal de Nível Superior, pelo apoio financeiro. REFERÊNCIAS BIBLIOGRÁFICAS
Copyright 2010 - Ciência Florestal The following images related to this document are available:Photo images[cf10022t1.jpg] [cf10022f2.jpg] [cf10022f5.jpg] [cf10022f1.jpg] [cf10022f8.jpg] [cf10022f7.jpg] [cf10022f6.jpg] [cf10022f3.jpg] [cf10022f4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}