 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
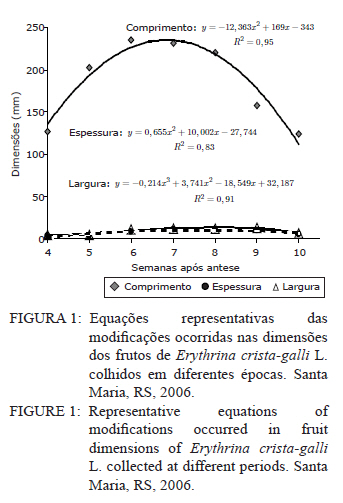
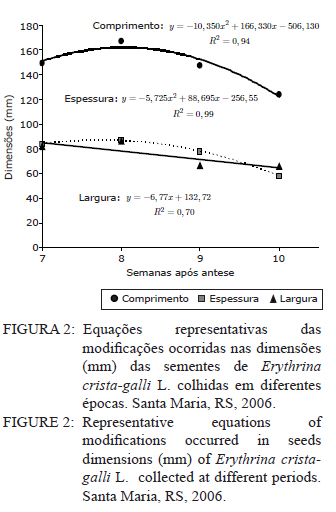
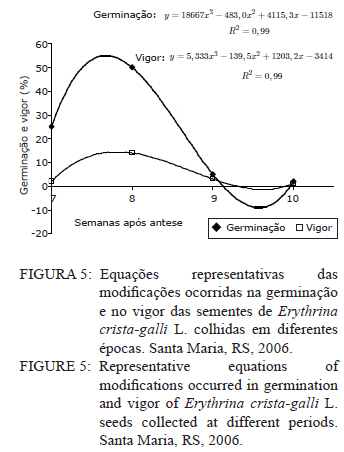
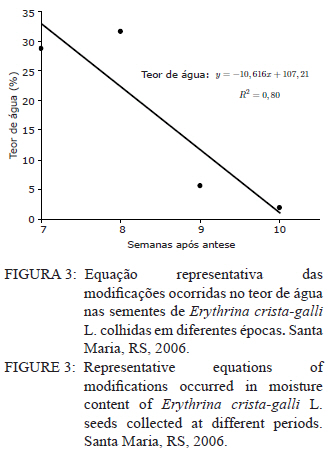
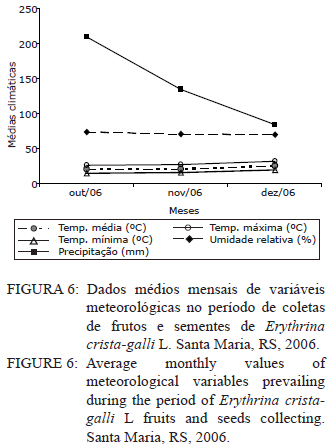
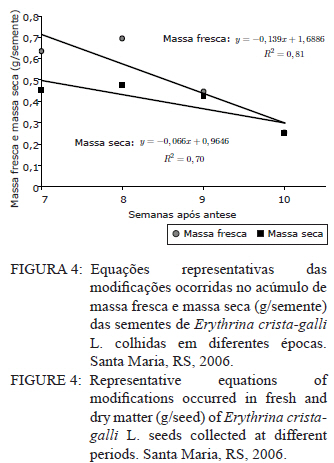
Ciência Florestal, Vol. 21, No. 1, Jan-Mar, 2011, pp. 9-16 Maturação fisiológica de sementes de Erythrina crista-galli L. Physiological maturation of Erythrina crista-galli L. Seeds Marília Lazarotto1 Rafael Beltrame¹ Marlove Fátima Brião Muniz2 Elena Blume² 1. Engenheira(o) Florestal, MSc., doutoranda(o) do Programa de Pós-Graduação em Engenharia Florestal, Centro de Ciências Rurais, Universidade Federal de Santa Maria, Av. Roraima, 1000, CEP 97105-900, Santa Maria (RS). lilalazarotto@yahoo.com.br/browbeltrame@yahoo.com.br. Recebido para publicação em 14/05/2008 e aceito em 14/09/2010 Code Number: cf11002 RESUMO Este trabalho objetivou investigar o processo de maturação fisiológica das sementes de Erythrina crista-galli L. a fim de que se determine o momento ideal de coleta. Para tanto, foram utilizadas seis árvores matrizes na cidade de Santa Maria (RS), de onde foram feitas coletas semanais, começando com o surgimento dos frutos até sua deiscência e posterior maturação das sementes. Os parâmetros indicadores da maturação das sementes observados foram: dimensão de frutos e sementes; massa fresca e massa seca das sementes; teor de água, teste de germinação (expresso em plântulas normais) e vigor (primeira contagem de germinação). A dimensão máxima das sementes ocorreu no período máximo de germinação, com valores de 167,0, 8,72 e 8,60 mm para comprimento, espessura e largura, respectivamente. O teor de água pode ser utilizado como indicativo de maturidade das sementes, assim como massa fresca e massa seca com valores máximos na oitava semana, 31,6 %, 0,693 e 0,474 g/semente, respectivamente. As sementes de Erythrina crista-galli apresentam dormência após a maturação fisiológica, com baixa percentagem de germinação na décima semana após a antese. O ponto de maturidade fisiológica, nas condições de Santa Maria – RS ocorre na oitava semana após a antese, com germinação de 50%, devido a todos os parâmetros observados terem assim indicado. Palavras-chave: corticeira-do-banhado; época de coleta; sementes florestais. ABSTRACT The objective of this research was to study the maturation process of Erythrina crista-galli L. seeds to determinate the ideal collect period. In this regard, six trees located in Santa Maria (RS) were used from which were collected weekly, starting with fruits and followed until seeds maturation. The parameters observed to indicate seed maturation were: fruit and seed dimensions; seed fresh and dry matter; moisture content, germination test (expressed by normal seedlings) and vigor (first count of germination). The seeds dimensions were highest in the same period of maximum germination, with values 167.0, 8.72 and 8.60 mm of length, thickness and width, respectively. The seeds moisture content could be used to indicate the seeds maturation, and also seeds fresh and dry matter with maximum values on the eighth week, 31.6 %, 0.693 and 0.474 g/seed, respectively. The Erythrina crista-galli has dormant seeds after physiological maturation, with low percentage of germination on the tenth week after anthesis. The point of physiological maturation in Santa Maria - RS conditions happens on the eighth week after anthesis, with germination percentage of 50%, which were indicate by all parameters observed. Keywords: Erythrina crista-galli; collect period; forest seeds. INTRODUÇÃO A espécie Erythrina crista-galli L. é uma planta pioneira, decídua, heliófita, de madeira leve e porosa, característica de terrenos úmidos existentes ao longo de rios e estuários, justificando seu nome popular de corticeira-do-banhado (LORENZI, 2002). Ainda, segundo o mesmo autor, a espécie produz anualmente grande quantidade de sementes, porém, estas são frequentemente atacadas por insetos, o que dificulta sua regeneração natural. De acordo com Carpanezzi et al. (2001), quando em populações naturais bem conservadas, somente 6% das flores da corticeira-do-banhado desenvolvem sementes. Isso decorre da autogamia, frequente em muitos sítios de coleta brasileiros, onde houve fragmentação da floresta original. A espécie possui valor ornamental, devido ao belo arranjo de sua copa e floração exuberante. Além disso, Gratieri-Sossella et al. (2008) ressaltam que a espécie é imune ao corte no Rio Grande do Sul pela Lei Estadual 9.519/92 (Art. 33º), que protege figueiras e corticeiras em todos os casos, exigindo imediata reposição da espécie em caso de sua eliminação. Apesar da importância da espécie, estudos sobre a caracterização e conservação de suas sementes ainda são escassos. O estudo da maturação fisiológica objetiva definir o momento ideal de colheita e o estádio de máxima qualidade das sementes, denominado de maturidade fisiológica (POPINIGIS, 1985), que varia em função da espécie e das condições ambientais. Após este momento, a permanência das sementes no campo resulta em sua deterioração progressiva. Segundo Nogueira e Medeiros (2007), para as espécies que apresentam frutos deiscentes, como é o caso da Erythrina crista-galli, há necessidade de acompanhar o estágio de maturação, para que se possa realizar a coleta antes que ocorra a abertura dos mesmos e, consequentemente, a dispersão das sementes. A determinação do ponto de maturidade fisiológica auxilia ainda no conhecimento do comportamento das espécies quanto à sua reprodução, bem como na obtenção de material genético de boa qualidade fisiológica, que é a base fundamental para os programas de melhoramento silviculturais, conservação genética e recuperação de áreas degradadas (FIGLIOLIA e KAGEYAMA, 1994). Esse processo resulta de alterações morfológicas, fisiológicas e funcionais, como aumento de tamanho, variações no teor de água, vigor e acúmulo de massa seca, que se sucedem desde a fertilização do óvulo até o momento em que as sementes estão maduras (CARVALHO e NAKAGAWA, 1983). Embora a Erythrina crista-galli tenha grande produção anual de sementes, muitas delas perdem sua viabilidade, dificultando a regeneração da espécie. Sendo assim, este trabalho teve como objetivo investigar o processo de maturação fisiológica das sementes de Erythrina crista-galli, a fim de determinar o momento ideal de coleta através de alguns parâmetros indicadores. MATERIAIS E MÉTODOS Caracterização da área de coleta As coletas foram realizadas entre outubro e dezembro de 2006, em áreas urbanas da cidade de Santa Maria, Rio Grande do Sul, com coordenadas aproximadas de 29º45’ latitude Sul e 53º43’ longitude Oeste em relação a Greenwich e altitude média de 110 m (BARICHELLO et al., 2000). De acordo com a classificação de Köeppen, Santa Maria possui clima do tipo Cfa, caracterizado por clima temperado, com chuvas bem distribuídas, temperaturas de clima subtropical e precipitação total do ano correspondente a 1686,1 mm (MORENO, 1961). Todas as avaliações dos parâmetros foram realizadas no Laboratório de Fitopatologia do Departamento de Defesa Fitossanitária do Centro de Ciências Rurais da Universidade Federal de Santa Maria. Caracterização das árvores matrizes Foram utilizadas seis árvores matrizes, nas quais se realizaram as marcações com barbante de algodão de dez inflorescências por árvore. Assim, fez-se a contagem do número de flores por inflorescência marcada para, posteriormente, ser feita a contagem do número de frutos por inflorescência, do número de sementes que inicialmente se formaram no fruto e das sementes que completaram sua maturação. Coleta dos frutos e sementes Os frutos foram coletados assim que começaram a ser formados, o que ocorreu na quarta semana após a antese. A partir da sétima semana após a antese, iniciou-se a coleta das sementes, pois estas já estavam formadas. O material coletado das seis matrizes foi homogeneizado e imediatamente conduzido ao laboratório para realização das análises. Para essa coleta, não foi realizada marcação prévia de inflorescências, a marcação (descrita no item 2.2) foi feita apenas para caracterização das matrizes. Parâmetros avaliados Dimensões de frutos e sementes. Comprimento, largura e espessura dos frutos e das sementes foram mensurados com o auxílio de um paquímetro digital e de régua graduada, sendo que as medições dos frutos iniciaram-se a partir da quarta semana após a antese, estendendo-se até décima semana. Já as medições das sementes iniciaram na sétima semana após a antese e estenderam-se até a décima semana, ocorrendo nesta a última coleta. Foram utilizados 40 frutos e 40 sementes para cada coleta, divididos em quatro repetições de dez unidades. Massa fresca de sementes. Foram utilizadas quatro repetições de dez sementes, pesadas com balança analítica de precisão 0,01 g. Os resultados foram expressos em g/semente. Massa seca e teor de água de sementes. As mesmas sementes utilizadas para determinação de massa fresca foram colocadas em estufa a 105±3°C por 24 horas (BRASIL, 1992). Posteriormente seu peso foi medido em balança analítica e, assim, foi determinado o teor de água (PU – PS/PS x 100) de sementes com massa fresca e massa seca, sendo que o peso úmido (PU) corresponde à massa fresca e peso seco (PS), à massa seca. Qualidade fisiológica (germinação e vigor). As sementes, depois de colhidas, foram imediatamente submetidas ao teste de germinação, que foi conduzido em caixas plásticas do tipo “gerbox” sobre três folhas de papel-filtro umedecidas com água destilada esterilizada. Foram utilizadas 100 sementes, divididas em quatro repetições de 25, acondicionadas à temperatura de 25°C e fotoperíodo de 12 horas de luz branca/12 horas de escuro. A primeira contagem foi realizada aos sete dias, computando-se a percentagem de plântulas normais, que foi considerada como avaliação de vigor. A germinação foi avaliada aos 14 dias, verificando-se as percentagens de plântulas normais, anormais, sementes não germinadas e mortas. Coleta de dados de variáveis climáticas. A coleta das variáveis climáticas mensais de temperatura média (°C), temperatura máxima média (°C), temperatura mínima média (°C), umidade relativa do ar (%) e precipitação média (mm) foi realizada na Estação Metereológica do Departamento de Fitotecnia da Universidade Federal de Santa Maria. Análise estatística O delineamento experimental utilizado foi inteiramente casualizado com quatro repetições, os dados foram submetidos à regressão polinomial, em função das semanas após antese para frutos e sementes, tendo sido testados os modelos linear, quadrático e cúbico, sendo que, para explicar os resultados, foi selecionado o modelo significativo de maior ordem, que promovesse as estimativas para a ocorrência de determinado evento. Para as análises, utilizou-se o Sistema de Análise Estatística - SANEST (ZONTA e MACHADO, 1986). RESULTADOS E DISCUSSÃO As árvores matrizes apresentavam, em média, 64 flores por inflorescência, formando 11 frutos por inflorescência (17,18% das flores). E nesses frutos, formavam-se, aproximadamente, 14 sementes, sendo que, em média dez foram abortadas (71,42%) e apenas quatro (28,57%) completavam sua maturação. Foram verificados efeitos significativos de ordem quadrática para o comprimento, largura e espessura dos frutos (Figura 1). Observou-se que o comprimento, a espessura e a largura foram aumentando até atingir valores máximos entre a sexta e a nona semana após antese. Iossi et al. (2007) verificaram que os frutos de Phoenix roebelenii O’ Brien apresentaram grande aumento no comprimento até o período de maturidade fisiológica e, a partir desse momento, tiveram seu crescimento estabilizado. O mesmo ocorreu em relação ao diâmetro, porém, no caso da corticeira-do-banhado, o valor máximo de diâmetro se deu um pouco depois da maturidade. Para Alves et al. (2005), a dimensão dos frutos de Mimosa caesalpiniifolia Benth. não foi um bom parâmetro para auxiliar na determinação do ponto de maturação fisiológica das sementes, o mesmo ocorrendo para Lopes et al. (2005) com Tibouchina granulosa Cogn. e para Leonhardt et al. (2001) com Citharexylum montevidense (Spreng) Moldenke., já que ambas as espécies citadas atingem valores máximos de dimensão dos frutos muito cedo, confirmando a afirmação de Edwards (1980 apud LEONHARDT et al., 2001) de que frutos de espécies florestais, geralmente atingem um tamanho próximo do máximo, relativamente cedo no período de maturação. Já para Cedrela fissilis Vell. foi possível determinar a maturidade fisiológica das sementes através da análise da coloração dos frutos (CORVELLO et al., 1999). No presente estudo, apenas o comprimento dos frutos pôde ser utilizado como parâmetro complementar para a determinação da maturidade das sementes, já que este teve acentuada queda em sua dimensão após a maturidade máxima, indicando o momento em que as sementes, provavelmente, já entravam em processo de dormência tegumentar. Na Figura 2, observa-se que o comprimento e a espessura das sementes ajustaram-se a modelos quadráticos. Já para largura, o modelo foi linear. As sementes atingiram comprimento, espessura e largura máximos estimados em oito semanas após antese, cujos valores foram de 167,00; 8,72 e 8,60 mm, respectivamente. Nesse caso, o tamanho das sementes foi um bom indicativo de maturidade fisiológica, já que alcançaram tamanho máximo na semana de pico de germinação (Figura 5). Para sementes de sabiá (Mimosa caesalpinifolia Benth.), o tamanho das sementes, em termos de comprimento, largura e espessura, ajustaram-se a modelos quadráticos, sendo que os valores máximos para esses parâmetros ocorreram aos 154 dias após a antese, exatamente quando se inicia o período de maturação fisiológica para essa espécie (ALVES et al., 2005). Diferentemente do ocorrido com sementes de corticeira-do-banhado no presente trabalho, Martins e Silva (1997) relataram que o comprimento, espessura e largura de sementes de Dalbergia nigra (Vell.) Allemao ex Benth. cresceram gradativamente até 16 dias antes da maturidade fisiológica, quando então começaram a decrescer. Em sementes de Phoenix roebelenii O’ Brien (IOSSI et al., 2007), não houve efeito significativo para comprimento, largura e espessura e, consequentemente, esses parâmetros não foram bons indicadores para a maturação. De uma maneira geral, as sementes aumentam de tamanho rapidamente, atingindo o valor máximo num período de tempo curto, em relação à duração total do período de maturação (CARVALHO e NAKAGAWA, 1983). Isso se deve, possivelmente, ao fato de estas terem de manter um alto teor de água durante a fase de maior acúmulo de massa seca. O valor máximo para o teor de água foi de 31,6%, alcançado na oitava semana após a antese, conjuntamente com a maturidade fisiológica e ajustado ao modelo linear (Figura 3). Após esse período, observou-se redução rápida desse parâmetro até a décima semana após a antese. A queda acentuada no teor de água da oitava semana (31,6%) para a décima semana (1,9%) após a antese foi favorecida pela secagem dos frutos, seguida da deiscência, e também pode ser considerada como um indicativo de maturação das sementes. Em sementes de ipê-amarelo (Tabebuia chrysotricha (Mart. Ex A. Dc.) Standl.), Fonseca et al., (2005) constataram que a maturidade fisiológica ocorreu quando o teor de água das sementes era de 61,3 % e, após este período, houve uma queda acentuada deste valor, assemelhando-se ao ocorrido com sementes de Erythrina crista-galli no presente estudo. Para Cedrela fissilis, a maturidade ocorreu quando as sementes possuíam teor de água entre 50 e 60% (CORVELLO et al., 1999). Para Dalbergia nigra (MARTINS e SILVA, 1997), o valor do teor de água das sementes decresceu continuamente de forma lenta até que, 16 dias antes da maturidade fisiológica, houve uma queda pronunciada, diferindo dos dados obtidos neste trabalho, com Erythrina crista-galli, na qual a queda foi acentuada após o ponto máximo de maturação. O valor extremamente baixo do teor de água das sementes de Erythrina crista-galli na décima semana após antese é resultado da entrada da semente em processo de dormência, sendo que, a partir desse momento, as sementes só germinarão se passarem por processo de superação de dormência, como a escarificação mecânica, por exemplo. A alta umidade inicial das sementes e o seu posterior decréscimo estão relacionados com a importância da água nos processos de enchimento e maturação das sementes (MARTINS e SILVA, 1997), uma vez que, após esses processos serem completados, a absorção de água é interrompida. O teor de água das sementes (Figura 3) não parece ter sido influenciado pelas condições climáticas da região para o período em estudo (Figura 6), já que o valor máximo alcançado para esse parâmetro ocorreu justamente no mês de dezembro, em que a precipitação pluvial e a umidade relativa foram inferiores ao registrado nos meses anteriores. Os valores máximos de massa fresca e massa seca ocorreram também na oitava semana após antese, sendo de 0,693 e 0,474 g/semente, respectivamente (Figura 4). Assim, esses parâmetros foram considerados adequados para indicar a maturidade fisiológica das sementes, já que os valores máximos ocorreram na semana de máxima germinação (Figura 5) e de máximo desenvolvimento das sementes (Figura 2). Em sementes de Eucalyptus grandis Hill ex Maiden (AGUIAR et al., 1988), Cedrela fissilis Vell. (CORVELLO et al., 1999) e Dalbergia nigra (Vell.) Allemao ex Benth. (MARTINS e SILVA, 1997), o acúmulo de massa fresca e massa seca também foram parâmetros indicadores do ponto de maturação fisiológica. O máximo peso de massa seca tem sido mencionado como o ponto em que a semente atinge a maturidade fisiológica, porém, essa característica não deve ser utilizada como única indicadora (LEONHARDT et al., 2001). Em trabalho realizado com Enterolobium contortisiliquum (Vell.) Morong., por Borges et al. (1980), constatou-se que o parâmetro massa seca não pode ser utilizado com eficácia para determinar o índice de maturidade, já que, para essa espécie, a variação do peso de massa seca foi muito pequena entre os diferentes estádios de maturação. Segundo Carvalho e Nakagawa (1983), o acúmulo de massa seca em uma semente em formação se faz, inicialmente, de maneira lenta, para, em seguida, começar uma fase de rápido e constante acúmulo, até que um máximo é atingido e mantido por algum tempo, podendo, no final, sofrer um pequeno decréscimo, como resultado de perdas pela respiração. Observou-se que, na sétima semana após a antese, a germinação foi de 25%, porém, constatou-se grande porcentagem de plântulas anormais, indicando que as sementes não estavam suficientemente desenvolvidas para germinarem e produzirem todas as estruturas essenciais da plântula normal (Figura 5). A máxima porcentagem de germinação (50%) ocorreu na oitava semana após a antese. Referindo-se aos dados da primeira contagem de germinação (vigor), a primeira contagem foi máxima (14%) também na oitava semana, sendo esta admitida como o período de maturação fisiológica. Tanto os dados de vigor quanto os de germinação ajustaram-se ao modelo cúbico (Figura 5). Após a oitava semana, a germinação decresceu significativamente, chegando a valores de 2%, na décima semana. Assim como ocorre com Erythrina crista-galli, as sementes de Tibouchina granulosa Cogn. apresentam dormência após a maturação fisiológica, com baixa percentagem de germinação (LOPES et al., 2005). A maturação de sementes ocorre em períodos variados, após a antese, sendo essa variação atribuída a características de cada espécie. Para Phoenix roebelenii, IOSSI et al. (2007) constataram que a maturação fisiológica das sementes ocorreu aos 138 dias após antese; para Miconia cinnamomifolia ocorreu aos 110 dias da antese (LOPES e SOARES, 2006); Tibouchina granulosa Cogn., entre 84 e 105 dias da floração (LOPES et al., 2005); e as sementes de Mimosa caesalpiniifolia Benth. alcançam a maturidade entre 154 e 168 dias após antese (ALVES et al., 2005). Outros autores apresentam a maturação com semanas após a antese, como Leonhardt et al. (2001), que verificaram a maturação de Citharexylum montevidense (Spreng.) Moldenke entre 29 e 33 semanas após a antese; Corvello et al. (1999), com maturação de Cedrela fissilis, entre 29 e 31 semanas; e Ragagnin et al. (1994), com sementes de Podocarpus lambertii Kl., aos 131 dias da floração. Já Souza e Lima (1985), observaram germinação máxima de sementes de Anadenanthera macrocarpa (Benth.) Brenan aos 220 dias após a frutificação. A maturidade fisiológica de Tabebuia chrysotricha (FONSECA et al., 2005) ocorreu antes da dispersão, em início de deiscência, na oitava semana, 56 dias após a antese, a exemplo do que ocorreu com Erythrina crista-galli neste estudo. Ávila et al. (2009) verificaram que o momento adequado para a coleta de sementes de Eugenia uniflora L. ocorreu entre 56 e 63 dias após a antese, e esta foi alcançada quando a altura, diâmetro e peso de frutos e sementes atingiram seus valores máximos e quando o teor de água e massa seca das sementes tenderam à estabilização, também concordando com resultados encontrados para Erythrina crista-galli. Segundo Muniz et al. (2003), em estudo da germinação de Erythrina crista-galli L., após adquirirem dormência, a germinação de sementes que passaram por escarificação mecânica foi de 34,67% e de sementes embebidas em água foi de 12%. Ambos os valores foram inferiores ao registrado no presente trabalho, no qual a germinação máxima alcançada ocorreu antes da entrada do período de dormência, quando as sementes ainda possuíam um elevado teor de água e o tegumento não se apresentava endurecido. Segundo Tokuhisa et al. (2008), a dormência da semente é um importante estádio do ciclo de vida das plantas, sendo caracterizada pela ausência temporária da capacidade de germinação, permitindo que as espécies vegetais sobrevivam às adversidades, principalmente aquelas que dificultem ou impeçam o crescimento vegetativo da planta. Os dados médios mensais de temperaturas máximas, mínimas, médias, precipitação pluvial e umidade relativa do ar ocorridas no período de outubro a dezembro de 2006 estão apresentados na Figura 6. Esse período refere-se ao período de coletas de frutos e sementes de Erythrina crista-galli. As variáveis climáticas podem influenciar em alguns parâmetros observados neste estudo e, consequentemente, na maturação das sementes. Observa-se que, no mês de outubro, ocorreram os maiores valores para precipitação pluvial (208,9 mm), sendo que, nos meses seguintes, a precipitação caiu muito, com 134,6 mm para o mês de novembro e 84,2 mm para o mês de dezembro. No primeiro mês de observação, os frutos estavam sendo formados, já que ainda encontravam-se flores nas árvores matrizes. A umidade relativa do ar chega ao máximo no mês de outubro (74%), coincidindo com o período de máxima qualidade fisiológica das sementes, porém, nos próximos meses não difere muito, com 71% para novembro e 70% para dezembro. Não existem grandes variações na temperatura média para os três meses apresentados, contudo, dezembro foi um mês de temperaturas mínimas e máximas ligeiramente superiores. CONCLUSÕES Para Erythrina crista-galli L., o ponto de maturidade fisiológica, nas condições de Santa Maria (RS), ocorre na oitava semana após a antese, sendo este o momento ideal de coleta das sementes. A maturação fisiológica das sementes é alcançada quando o comprimento dos frutos, largura comprimento, espessura, teor de água, massa fresca e massa das sementes atingem seus valores máximos. As sementes de Erythrina crista-galli L. apresentam dormência após a maturação fisiológica, com germinação muito baixa na décima semana após a antese. REFERÊNCIAS BIBLIOGRÁFICAS
Copyright © 2011 - Ciência Florestal The following images related to this document are available:Photo images[cf11002f4.jpg] [cf11002f2.jpg] [cf11002f1.jpg] [cf11002f3.jpg] [cf11002f6.jpg] [cf11002f5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}