 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
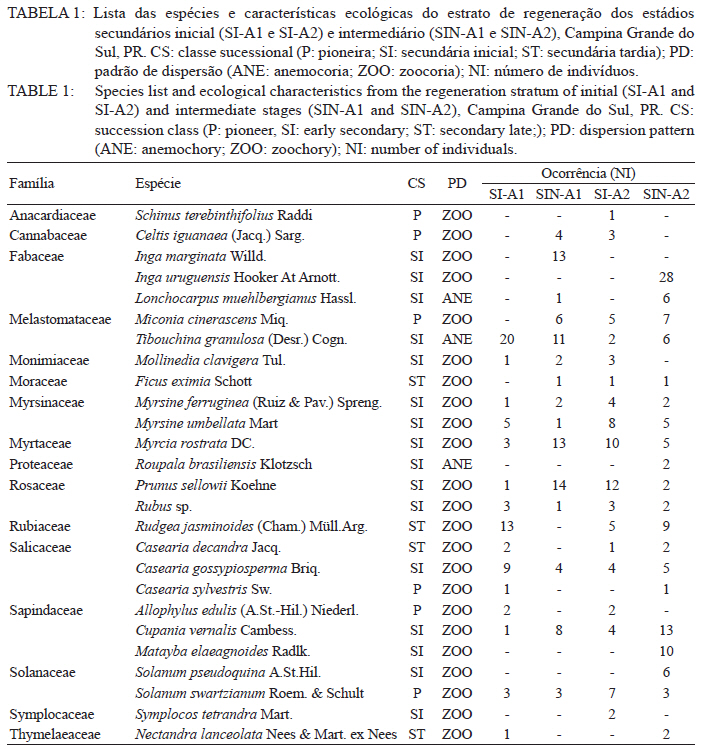
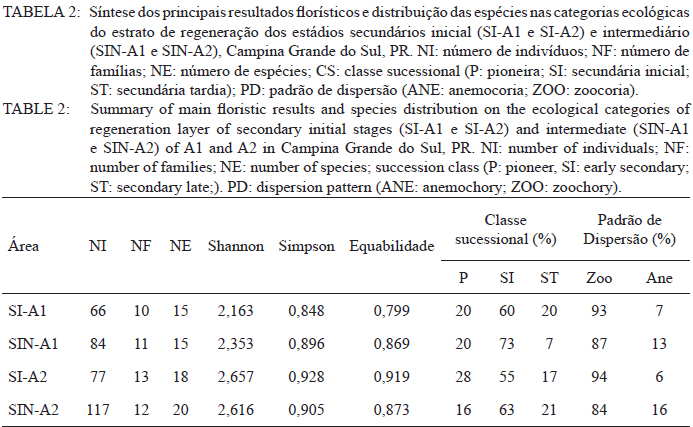
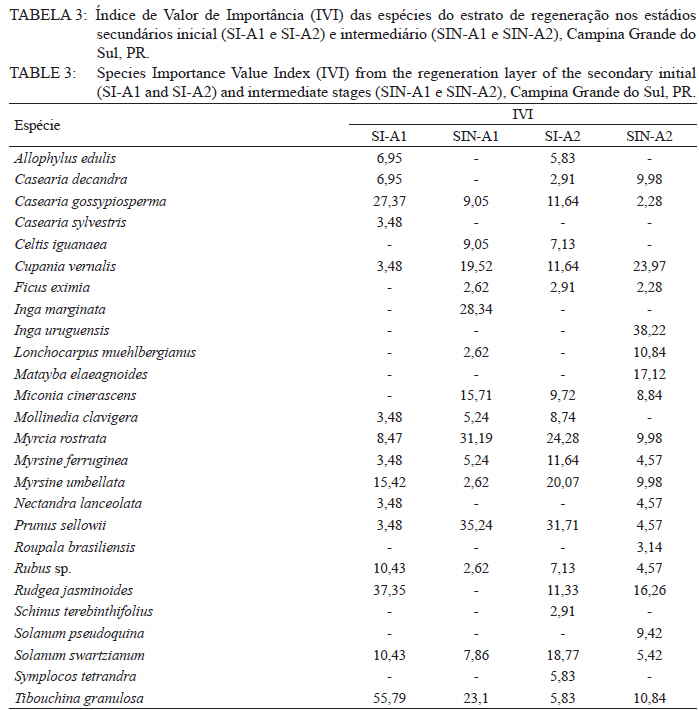
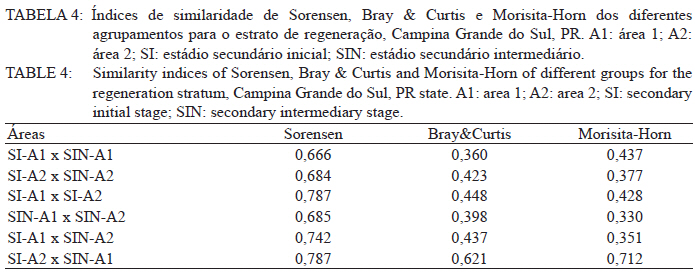
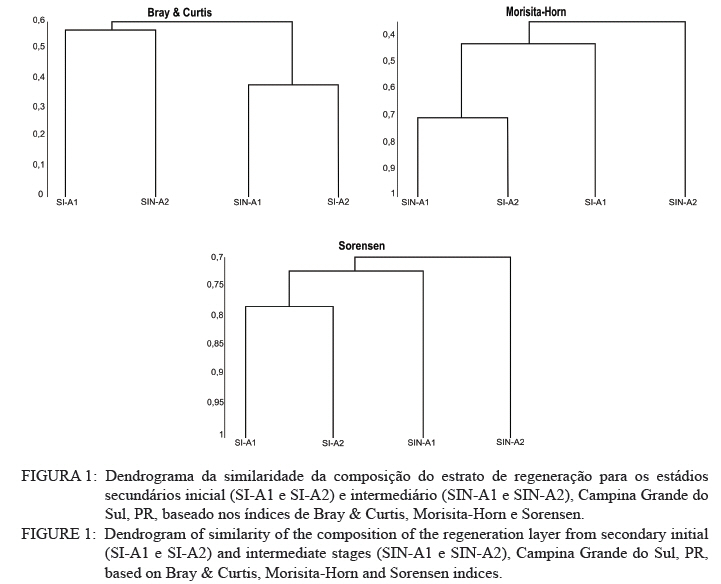
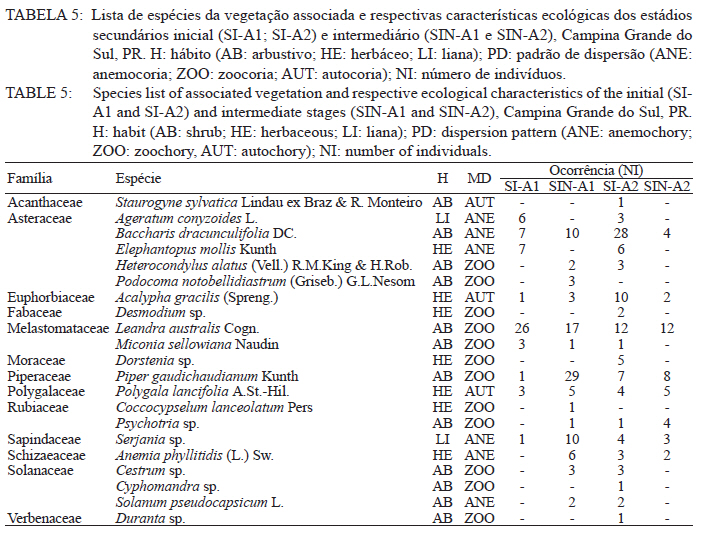
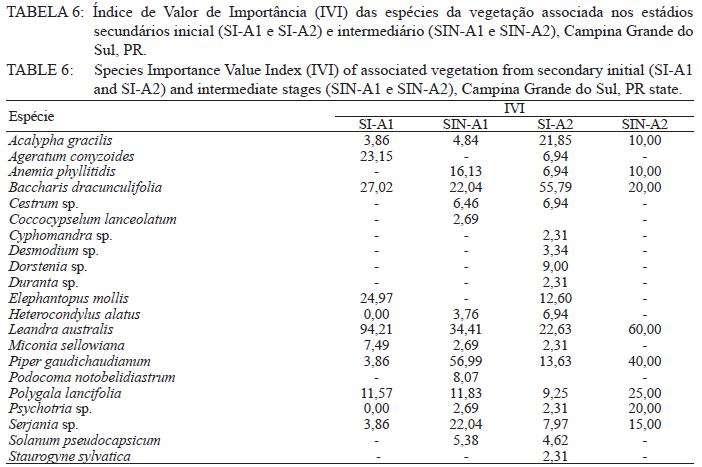
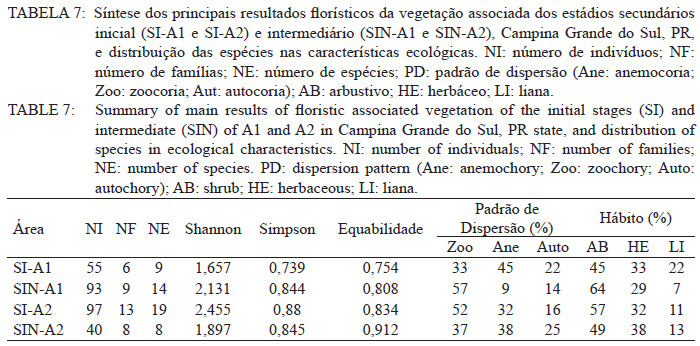
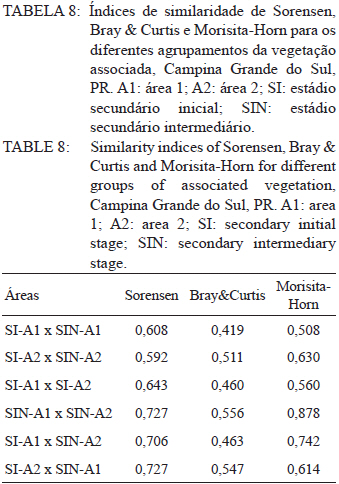
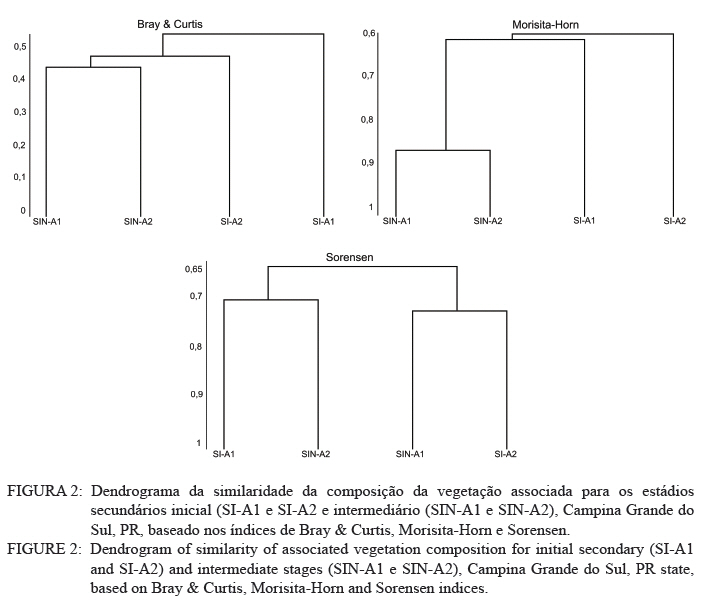
Ciência Florestal, Vol. 21, No. 3, Jul-Sept, 2011, pp. 393-407 Estrutura e composição do estrato de regeneração e vegetação associada de diferentes estádios sucessionais no leste do Paraná1 Structure and composition of the regeneration stratum and associated vegetation of different succession stages in east of Paraná state Marco Antonio Gomes Souto2 Maria Regina Torres Boeger3 1. Parte da Dissertação de mestrado do primeiro autor apresentada ao Programa de Pós-Graduação em Ecologia e Conservação da Universidade Federal do Paraná (PR). Recebido para publicação em 8/07/2009 e aceito em 08/10/2010 Code Number: cf11041 RESUMO Nas últimas décadas, as atividades antrópicas têm gerado o declínio geral da biodiversidade dos diferentes biomas e para compreender os impactos na comunidade vegetal é necessário analisar o potencial regenerativo das florestas. O estudo da composição e estrutura florística da guilda de plântulas em diferentes estádios sucessionais e condições históricas descreve os padrões de substituição das espécies e auxilia no processo de compreensão da dinâmica florestal. O objetivo do trabalho foi comparar florística e estruturalmente o estrato de regeneração e da vegetação associada em dois estádios sucessionais de duas áreas com históricos de uso distintos no Município de Campina Grande do Sul, PR. As plântulas foram coletadas em dois fragmentos de floresta secundária nos estádios secundários inicial e intermediário com históricos de uso de corte raso e seletivo, respectivamente. Cento e sessenta parcelas de 0,16 m² foram demarcadas, onde todos os indivíduos entre 10 - 50 cm de altura foram coletados, identificados em herbário e separados em dois grupos para a análise fitossociológica: plântulas e espécies associadas. As espécies também foram classificadas quanto ao seu hábito, modo de dispersão e classe sucessional, e a similaridade florística entre as áreas foi estabelecida pelos índices de Sorensen, Bray & Curtis e Morisita-Horn. A análise geral indicou uma possível influência do histórico de uso e dos estádios sucessionais no estrato de regeneração e da vegetação associada. A análise dos índices de similaridade mostrou um gradiente de regeneração, sendo que o estádio intermediário da área com corte raso é semelhante ao estádio inicial da área com corte seletivo. Palavras-chave: estádio sucessional; histórico de uso; plântulas; regeneração. ABSTRACT In recent decades, the human activities caused the overall decline in biodiversity and, to understand the impacts on plant communities, it is necessary to analyze the regenerative potential of the forest. Studies on the floristic composition and the structure of the guild of seedlings in different succession stages and historical conditions can describe the patterns of species replacement and help understand the forest dynamics. Thus, the objective of this study was to compare floristic and structural layer of the regeneration and associated vegetation in two succession stages of two areas with different historical uses located in the city of Campina Grande do Sul, in Paraná state. Seedlings were collected in two fragments of the secondary forest at the initial and at the intermediate stages, with historical use of clear and selective cuts, respectively. One-hundred and six plots of 0,16 m² were delimited and all individual plants with up to 10 - 50 cm high were collected, identified, quantified and divided into two groups: seedlings of tree species and species associated for the phytosociological analysis. The species were also classified according to their habits, dispersion mode and succession stages and the floristic similarity between areas was established by Sorensen’s, Bray & Curtis’ and Morisita-Horn’s indices. The analysis suggests a possible influence of historical use and the successional stages of regeneration and the associated vegetation. The analysis of similarity indicates a gradient of regeneration, in which the intermediate stage of the clear cut area is similar to the early stage of the selective cutting area. Keywords: historical use; seedlings; succession stages; regeneration. INTRODUÇÃO Nas últimas décadas, o declínio geral da biodiversidade vem sendo observado especialmente nas florestas tropicais e subtropicais, causado principalmente pelo desmatamento, a fragmentação de habitats e a mudança de uso do solo (Laurance et al., 2000). As consequências da redução da biodiversidade nestes ecossistemas ainda não são bem compreendidas, o que justifica o crescente interesse dos pesquisadores e os estudos dos seus efeitos em curto, médio e em longo prazo (Farwig et al., 2006; Larsen et al., 2005; Laurance et al., 2000). Apesar dos impactos antrópicos imediatos, a diversidade de indivíduos arbóreos adultos pode permanecer elevada devido ao longo ciclo de vida de muitas árvores tropicais. Mas, para manter a riqueza em longo prazo, é necessário que essas espécies vegetais completem com sucesso seu ciclo reprodutivo (Janzen, 1986). Dessa maneira, para compreender os diversos resultados dos impactos antrópicos na comunidade vegetal é necessário, além da análise da fase adulta formadora do dossel, um estudo das fases iniciais do ciclo de vida, formadoras do potencial regenerativo da floresta. O recrutamento e o desenvolvimento das plântulas são eventos reguladores do crescimento e da manutenção das populações vegetais (Melo et al., 2004), sendo que a sobrevivência no período inicial de vida afeta não só a abundância e a distribuição de uma população ou espécie, mas toda a composição e estrutura da comunidade (Denslow, 1991). Desse modo, as plântulas desempenham importante papel no ecossistema florestal, visto que sua distribuição e abundância implicam na disposição dos futuros indivíduos adultos da população e na dinâmica da comunidade vegetal como um todo (Harper, 1977). Como é praticamente impossível acompanhar o desenvolvimento de um indivíduo da fase de plântula até o estágio adulto, o estudo do estrato de regeneração e vegetação associada é um primeiro indicador da capacidade de regeneração da floresta. A composição e densidade do estrato de regeneração dependem, entre outros fatores, do histórico de perturbação e da idade da floresta secundária (Baider et al., 2001). As condições bióticas e abióticas encontradas em florestas secundárias, quando comparadas com áreas de floresta madura, podem influenciar no equilíbrio dos fatores causadores de mortalidade e na abundância de espécies e na diversidade da comunidade (Cersósimo, 1993). Assim sendo, variações na composição e na estrutura da guilda de plântulas entre estádios sucessionais e áreas com diferentes históricos de uso são esperadas. Acredita-se que o conhecimento sobre a composição e a estrutura florística do estrato regenerativo em diferentes estádios sucessionais e condições históricas pode gerar importantes informações sobre a dinâmica florestal (Mendes, 2002), além de constituir um indicador da degradação e da capacidade de regeneração das florestas secundárias (Janzen, 1986). Portanto, o objetivo do presente estudo foi comparar florística e estruturalmente o estrato de regeneração e vegetação associada de estádios sucessionais secundários em dois fragmentos florestais com histórico de uso distinto, localizadas no Município de Campina Grande do Sul, Paraná. MATERIAL E MÉTODO Áreas de estudo O estudo foi desenvolvido em dois fragmentos florestais contíguos (Área 1: 25°07’14,40” S/ 48°50’54,49” O e 25°07’52” S /48°50’37” O; Área 2 - 25°07’17,89” S/48°50’46” O e 25°07’48,13” S/ 48°50’34,12” O) localizados em Campina Grande do Sul, município a 30 km a leste de Curitiba, no Primeiro Planalto Paranaense (MAACK, 1968). O clima na região é de transição entre Cfa e Cfb, conforme classificação de Köppen, caracterizado como um clima quente temperado subtropical sem período de seca, com temperatura média variando entre 16 - 17°C, sendo que a temperatura média mínima no inverno é de 12°C, com ocorrência de geadas, e a temperatura média máxima é de 22°C no verão (SIMEPAR, 2008). Apresenta um índice pluviométrico de 1.400 - 1.500 mm/ano com umidade relativa do ar permanecendo entre 80% e 85%, sendo os meses com maior precipitação dezembro, janeiro e fevereiro, não havendo período de seca (SIMEPAR, 2008). Os fragmentos florestais encontram-se em região de transição entre Floresta Ombrófila Densa e Floresta Ombrófila Mista, sendo classificada como Floresta Ombrófila Montana (IBGE, 1992). A altitude média das áreas é de 900 m apresentando relevo levemente irregular, com áreas de platôs e aclives, circundadas por cadeia de formações rochosas de sul a leste, representando uma transição de formações geológicas entre a planície litorânea, a Serra do Capivari e o planalto (MACK, 1968). Os solos são do tipo Neossolo Litólico Distro-úmbrico, fragmentário e com textura argilosa e Neossolo Litólico Distrófico, fragmentário e com textura média (EMBRAPA, 2006). No entorno, encontra-se atividades agropecuárias, povoamento de espécies exóticas (Pinus sp.) e remanescentes florestais nativos pertencente às Áreas de Proteção Permanente da Serra do Capivari. A amostragem e coleta de material foram realizadas em dois fragmentos florestais de 20 ha cada um, denominados A1 e A2 com base em seus respectivos históricos de uso, recompostos através de informações fornecidas pelo atual proprietário. Em A1 o histórico de uso remete ao corte raso e frequentes queimadas da vegetação para estabelecimento de pastagem e, após um período de tempo indeterminado, algumas áreas foram abandonadas gradativamente tendo início a regeneração natural. Assim, áreas que foram abandonadas há cerca de cinco anos, atualmente apresentam uma vegetação em estádio secundário inicial de regeneração e áreas abandonadas há 40 anos apresentam vegetação em estádio secundário intermediário de regeneração. Em A2 o histórico de uso refere-se ao corte seletivo de indivíduos arbóreos em diferentes intensidades, sendo que em alguns trechos a vegetação foi quase totalmente retirada, atividade que também foi abandonada gradativamente, possibilitando a regeneração natural das áreas. Igualmente a A1, alguns trechos foram abandonados há cinco anos, possuindo vegetação em estádio secundário inicial e outros trechos foram abandonados há aproximadamente 40 anos, apresentando vegetação em estádio secundário intermediário de regeneração. Portanto, os estádios sucessionais foram definidos de acordo com seu tempo de abandono e características físicas segundo CONAMA (1993). Coleta de material Em cada estádio sucessional e em cada área foram demarcadas 40 parcelas alocadas aleatoriamente com as dimensões de 40 x 40 cm (0,16 m²) em duas etapas (janeiro e julho de 2008), totalizando 160 parcelas e uma área amostral de 25,6 m². Todos os indivíduos vegetais com altura entre 10 - 50 cm (MELO et al., 2004) que se encontravam no interior das parcelas foram coletados e acondicionados em sacos plásticos para posterior identificação. O material vegetal foi herborizado, identificado taxonomicamente. As espécies de angiospermas foram reunidas em famílias de acordo com o Sistema de Classificação APG II (Angiosperm Phylogeny Group, 2003) e classificadas em duas categorias: estrato de regeneração - composto pelas plântulas de espécies arbóreas, e vegetação associada - composta por indivíduos jovens de espécies arbustivas, herbáceas e lianas. Análise dos dados Os dados das coletas foram organizados em quatro grupos, de acordo com a área: estádio secundário inicial (SI-A1) e intermediário (SIN-A1) de A1; estádio secundário inicial (SI-A2) e intermediário (SIN-A2) de A2. A análise da composição florística e diversidade, assim como da distribuição das espécies em grupos sucessionais, modos de dispersão e hábito foi realizada separadamente por estrato de regeneração e vegetação associada. Para cada grupo foram calculados os valores de densidade, frequência e Índice de Valor de Importância, pelo programa Fitopacshell1.6.4 (SHEPHERD, 2006) e descritos por Mueller-Dombois e Ellenberg (1974), além dos índices de Diversidade de Shannon (H’), de Simpson (D) e a equabilidade (MAGURRAN, 1988). Com relação ao hábito, as espécies da vegetação associada foram classificadas em arbustiva, herbácea e liana de acordo com Oliveira et al. (2001). Para a classificação em grupos sucessionais das espécies do estrato de regeneração foram utilizados os critérios propostos por Budowski (1970), considerando-se as categorias: pioneiras (P), secundárias iniciais (SI) e secundárias tardias (ST). A síndrome de dispersão dos propágulos baseou-se na classificação de Van der Pijl (1982): zoocórica (dispersão realizada por animais), anemocórica (dispersão realizada pelo vento) e autocórica (dispersão realizada pela própria planta). A similaridade florística foi calculada entre os diferentes grupos (SI-A1xSIN-A1, SI-A2xSIN-A2, SI-A1xSI-A2, SIN-A1xSIN-A2), pelos índices de Sorensen (MUELLER-DUMBOIS e ELLENBERG, 1974), Morisita-Horn e de Bray & Curtis (MAGURRAN, 1988). Nesse último, por ser uma medida de distância, a similaridade foi obtida pela diferença entre 1 e o valor calculado pelo índice. Os valores dos índices calculados variam de 0 a 1, assumindo-se como valor crítico de similaridade 0,5 (50%), segundo Oliveira et al. (2001). Com os resultados das análises de similaridade foram elaborados dendrogramas para visualizar as similaridades e possíveis agrupamentos utilizando-se o método do pareamento por média matemática não ponderada (UPGMA), por meio do programa Fitopacshell 1.6.4. RESULTADOS E DISCUSSÃO Estrato de regeneração Composição florística e estrutura No total, 344 indivíduos pertencentes a 26 espécies, distribuídas entre 16 famílias foram coletados. As famílias que apresentaram um destacado número de espécies e indivíduos foram Melastomataceae, Rosaceae, Fabaceae, Salicaceae e Sapindaceae que, somadas, representam 50% das espécies e aproximadamente 62% dos indivíduos. A lista de espécies e suas respectivas características ecológicas encontram-se na Tabela 1. Apesar do número relativamente baixo de indivíduos e espécies coletados, as curvas cumulativas de espécies de cada estádio sucessional em cada área estabilizaram-se satisfatoriamente em torno da parcela 30, o que foi verificado através da interpretação da curva de espécie/área (MUELLER-DOMBOIS e ELLENBERG, 1974). A maioria dessas famílias é destacada na composição florística da Floresta Atlântica do sul-sudeste brasileiro (OLIVEIRA-FILHO e FONTES, 2000; SCUDELLER et al., 2001; TABARELLI e PERES, 2002). Entretanto, algumas famílias comuns e importantes na composição da comunidade vegetal observadas em levantamentos florísticos de plântulas e do componente arbustivo-arbóreo das florestas tropicais como Myrtaceae, Rubiaceae e Thymelaeaceae (OLIVEIRA et al., 2001; IVANAUSKAS et al., 2002; NAVE, 2005) foram consideradas raras neste estudo, isto é, foram encontradas em número reduzido e/ou em estádios sucessionais restritos, destacando-se Myrtaceae e Thymelaeaceae, com apenas uma espécie encontrada. Essas duas famílias podem ser indicadoras do estádio de regeneração ou de degradação ambiental (TABARELLI e MANTOVANI, 1999), sendo importantes na disponibilidade de recursos para grandes e médios frugívoros, além do destaque na composição florística do sub-bosque e do dossel de florestas subtropicais. A análise das áreas A1 e A2 e seus respectivos estádios sucessionais indicou diferenças relevantes entre os descritores ecológicos utilizados e as espécies em destaque. Os estádios secundários inicial e intermediário de A2 apresentaram maiores índices de Shannon, Simpson e equabilidade quando comparados, respectivamente, com os mesmos estádios secundários de A1 (Tabela 2), além do maior número de famílias e espécies, o que indica uma melhor estruturação do estrato de regeneração de A2, em termos florísticos, decorrente do histórico de uso menos intenso (OLIVEIRA et al., 2001). Apesar disso, esses valores estão muito aquém do encontrado para vegetação arbórea adulta na mesma fitorregião em outros trabalhos, que levantaram cerca de 80 espécies e índice de diversidade de Shannon-Weaver (H’) ao redor de 3,50 (SILVA, 1994; REGINATO e GOLDENBERG, 2007). Na análise comparativa entre os estádios sucessionais de A1, o estádio intermediário apresentou maiores valores para os índices de diversidade e número de indivíduos, mas o número de espécies e famílias é semelhante quando comparado com o estádio inicial. Possivelmente, os fatores físicos limitantes, como a maior incidência de luz e baixa umidade (MARTINS e RODRIGUES, 2002) do estádio inicial e/ou ocorrência de fatores bióticos como a competição, herbivoria ou predação (BENÍTEZ-MALVIDO e MARTÍNEZ-RAMOS, 2003) influenciaram no recrutamento de um menor número de espécies. Em A2, o estádio inicial apresentou os maiores valores para os índices de diversidade e número de famílias, quando comparado com o estádio intermediário, porém com um número menor de indivíduos e espécies. Isso pode ser decorrente da estrutura florestal presente no estádio intermediário de A2, com uma estratificação vertical definida e dossel compacto, que acarretou uma redução natural do número de espécies com capacidade de estabelecimento, tendência observada durante a regeneração natural de áreas do sul e sudeste brasileiro (TABARELLI et al., 1994; PESSOA et al., 1997). A composição florística do estrato de regeneração apresentou diferenças relativas ao maior valor de importância (VI) das espécies entre os estádios sucessionais e áreas (Tabela 3). Em SI-A1 as espécies com maior VI foram Tibouchina granulosa, Rudgea jasminoides e Casearia gossypiosperma, espécies de ampla distribuição no domínio da Floresta Atlântica (SCUDELLER et al., 2001), sendo que a ocorrência das duas últimas não era esperada numa área no início de regeneração natural, por se tratarem de espécies secundárias iniciais e tardias e por estas áreas possuírem condições abióticas mais adversas ao estabelecimento de espécies arbóreas com características de estádios mais avançados de regeneração natural (KELLMAN, 1980). Alguns fatores podem ter ocasionado essa alta ocorrência, dentre eles o efeito de filtro das árvores do dossel que poderia parcialmente determinar a composição e estrutura da comunidade de plantas sob a projeção da sua copa (GANDOLFI et al., 2007) ou a alta sensibilidade de algumas espécies à heterogeneidade luminosa do sub-bosque (MONTGOMERY e CHAZDON, 2002), que influenciariam o recrutamento de espécies vegetais. As espécies com maior VI de SIN-A1 foram Inga marginata, Prunus sellowii e Myrcia rostrata, sendo que as duas últimas também se destacam em SI-A2, indicando uma certa similaridade entre os estádios. Em SI-A2, além das espécies já mencionadas, destacaram-se Myrsine umbellata e Solanum swartzianum, sendo esta última uma espécie pioneira de ocorrência esperada em estádios iniciais de regeneração. Em SIN-A2, as espécies com maior VI foram Inga uruguensis, Cupania vernalis, Rudgea jasminoides e Matayba elaeagnoides, sendo as duas últimas espécies características de florestas primárias ou de estádio avançado de regeneração florestal (LORENZI, 2000), podendo indicar um maior desenvolvimento da área, no estádio intermediário. Apesar das diferenças de composição, a maioria das espécies encontradas nas duas áreas apresenta características comuns, como ampla distribuição, frutificação intensa e produção de sementes viáveis, além da dispersão realizada por aves (LORENZI, 2000; 2002). Dessa maneira, o pouco tempo de abandono e a regeneração natural das áreas estudadas podem explicar a ausência de espécies consideradas raras e o domínio de espécies comuns, situação que pode se alterar com o avanço da sucessão, já que em florestas tropicais secundárias a riqueza florística, com a entrada de espécies endêmicas, é a última característica ecológica a ser recuperada (LIEBSCH et al., 2008). De acordo com Chiarello (1999), a porcentagem de espécies zoocóricas na Floresta Atlântica alcança entre 70 - 90%, sendo que em florestas secundárias essa porcentagem seria atingida após 80 anos de regeneração natural (LIEBSCH et al., 2008). A relação direta entre o amadurecimento da floresta e a proporção de espécies zoocóricas, observada por outros autores (TABARELLI et al., 1994; TABARELLI e PERES, 2002; NUNES et al., 2003), não foi encontrada no presente estudo, sendo a zoocoria amplamente dominante nos estádios sucessionais de ambas as áreas, numa proporção média de 90%. A recuperação acelerada desse fator pode ser decorrente dos principais grupos de dispersores encontrados em áreas em estádios sucessionais iniciais (NUNES et al., 2003) e dos frugívoros generalistas que otimizam a dispersão das espécies vegetais mais comuns (CHIARELLO, 1999; DE WALT et al., 2003; NUNES et al., 2003). Portanto, há a possibilidade de uma alta atividade dos dispersores locais, em função do estado avançado de conservação das áreas do entorno contíguas pertencentes a Serra do Capivari (PR) que podem servir de refúgio desses dispersores. Uma vez que a flora característica das áreas no início da sucessão secundária é representada principalmente por espécies pioneiras e secundárias iniciais (TABARELLI e MANTOVANI, 1999), era esperado que esses grupos sucessionais fossem os mais encontrados nas áreas analisadas (Tabela 2). Entretanto, pequenas diferenças foram observadas na análise dos estádios sucessionais, separadamente. Em SI-A1, as espécies secundárias tardias corresponderam a 20% do total de espécies, com valor semelhante ao encontrado em SIN-A2 e superior ao encontrado em SIN-A1 (7%). Esse valor não era esperado para uma área no início de regeneração e histórico de uso com alto potencial de degradação (TABARELLI et al., 1994). No estádio inicial de A2, destacaram-se as espécies pioneiras e secundárias iniciais, enquanto que no estádio intermediário as espécies secundárias iniciais e tardias tiveram destaque, fato encontrado por outros autores (TABARELLI e MANTOVANI, 1999). Esses dados podem indicar que o processo de regeneração, no que se refere à distribuição das espécies entre os grupos sucessionais, está mais estruturado em A2 devido ao menor impacto das atividades antrópicas passadas, propiciando condições favoráveis às espécies secundárias em detrimento das pioneiras. Embora SIN-A2 apresente um menor número de espécies pioneiras, este é ainda considerado alto dentro do padrão esperado para as espécies pioneiras de florestas mais conservadas (10%; TABARELLI e MANTOVANI, 1999). A proporção de espécies pioneiras encontradas pode ser consequência do conjunto dos fatores ambientais como maior intensidade luminosa e maior temperatura proporcionada pela heterogeneidade da cobertura do dossel, favorecendo o estabelecimento dessas espécies (BENÍTEZ-MALVIDO, 1998; TABARELLI e MANTOVANI, 1999). Similaridade florística A análise da similaridade florística entre os estádios sucessionais realizada pelo índice de Sorensen indicou alta similaridade entre todas as situações (Tabela 4) e a interpretação do dendrograma (Figura 1) mostra que os estádios iniciais são mais semelhantes entre si e os estádios intermediários formam grupos à parte. Já os índices de Bray & Curtis e Morisita-Horn apresentaram resultados diferentes de Sorensen, mas semelhantes entre si, indicando semelhança apenas entre SI-A2 e SIN-A1 (Figura 1). A interpretação do dendrograma elaborado através dos dois índices sugere um gradiente entre os estádios, sendo SI-A1 e SIN-A2 completamente diferentes, indicando que o estádio intermediário de uma área com histórico de uso impactante é semelhante a uma área em estádio inicial que possui histórico de uso de menor impacto. Dessa maneira, o gradiente de similaridade sugerido é: SI-A1, SI-A2, SIN-A1 e SIN-A2. Vegetação associada Composição florística e estrutura Na vegetação associada, foram coletados 285 indivíduos de 21 espécies pertencentes a 13 famílias. A lista com as espécies e suas respectivas características ecológicas encontra-se na Tabela 5 . Apesar do reduzido número de indivíduos e espécies coletados, as curvas cumulativas de espécies de cada estádio sucessional em cada área estabilizaram-se satisfatoriamente em torno da parcela 32, o que foi verificado através da interpretação da curva de espécie/área (MUELLER-DOMBOIS e ELLENBERG, 1974). A composição florística da vegetação associada nas áreas analisadas mostrou-se mais homogênea do que o estrato de regeneração, sendo que as famílias Asteraceae, Melastomataceae e Piperaceae foram as mais destacadas, apresentando maiores valores de importância, na formação do sub-bosque florestal (OLIVEIRA-FILHO e FONTES, 2000; TABARELLI e PERES, 2002). Apesar da contribuição da vegetação associada na regeneração natural e constituição da estrutura florestal ser incipiente, indivíduos arbustivos, herbáceos e lianas podem interferir diretamente no estabelecimento e sobrevivência de plântulas de espécies arbóreas (CERSÓSIMO, 1993). Comparando-se os valores de importância da vegetação associada, nota-se que SI-A1 e SIN-A2 possuem valores aproximados, enquanto SIN-A1 e SI-A2 também possuem certa proximidade entre si (Tabela 6). Provavelmente, em SI-A1 e SIN-A2 os valores foram menores devido a diferentes fatores, pois em SI-A1, o ambiente pode oferecer condições ambientais mais restritivas, impedindo que um maior número de espécies se estabeleça (KELLMAN, 1980), enquanto que em SIN-A2, as espécies arbóreas de ciclo de vida longo podem estar substituindo espécies arbustivas pioneiras (TABARELLI e MANTOVANI, 1999), embora a tendência seja o incremento do número de espécies arbóreo-arbustivas no transcorrer da regeneração natural (PESSOA et al., 1997). Essa substituição ocorre em função das mudanças das condições abióticas, conferindo maior habilidade competitiva às espécies arbóreas, em detrimento das herbáceas e arbustivas (NYKVIST, 1996). Em SIN-A1 e SI-A2, as condições ambientais parecem ter maior amplitude, possibilitando a convivência de um maior número de espécies. Dados semelhantes foram encontrados por Tabarelli e Mantovani (1999), onde uma floresta com 40 anos de regeneração natural apresentou riqueza e diversidade de espécies superiores às observadas em floresta madura e em áreas com menor tempo de regeneração natural. A homogeneidade florística também se refletiu na lista de espécies de cada área, com destaque para Leandra australis, presente nas quatro áreas, Baccharis dracunculifolia, presente em todas as áreas exceto em SIN-A2 e Piper gaudichaudianum, presente nos estádios intermediários. Baider et al. (2001) verificaram que Baccharis dracunculifolia é característica de floresta secundária com histórico de uso de agricultura, enquanto Leandra australis é mais frequente no sub-bosque de florestas maduras. Assim, entende-se que essas áreas estão em transição em virtude do destaque das duas espécies com características inversas. Na análise da distribuição das espécies quanto à dispersão encontrou-se tendência semelhante àquela encontrada para os fatores fitossociológicos, com a formação de dois grupos (Tabela 7). Em SI-A1 e SIN-A2 apresentou uma distribuição mais homogênea, com um maior número de espécies anemocóricas, enquanto SIN-A1 e SI-A2 apresentou o domínio das espécies zoocóricas. Devido às escassas informações sobre classificação das espécies quanto à dispersão, não se pode afirmar que essas diferenças sejam derivadas da influência do estádio sucessional ou do histórico de uso da área. Na distribuição dos hábitos, as espécies arbustivas se destacaram com cerca de 55% do total das espécies da vegetação associada, seguida de espécies herbáceas (33%) e lianas (12%), conforme a Tabela 7. Esses valores podem indicar a reestruturação florestal das áreas, já que os valores de espécies formadoras do sub-bosque florestal se aproximam do esperado para as áreas preservadas ou em estádio avançado de regeneração florestal, que é de 50% (SILVA, 1994). Entretanto, apenas os estádios intermediários de A1 e A2 apresentam sinais de estratificação vertical, uma das principais características de estádios mais avançados da sucessão florestal (TABARELLI et al., 1994). Similaridade florística A análise da similaridade florística da vegetação associada pelos índices de Sorensen e Morisita-Horn indicou uma alta similaridade em todos os agrupamentos enquanto o índice de Bray & Curtis indicou pequena dissimilaridade para alguns grupos (Tabela 8) com valores próximos do limite de similaridade. Assim, a afirmação categórica sobre a diferença entre os estádios não é possível, com base nessa análise. A alta similaridade encontrada pode ser devido às características bióticas e abióticas que possibilitaram o estabelecimento das mesmas espécies (KELLMAN, 1980), e não foi encontrada por Liebsch e Acra (2004) em formação vegetal e estádios sucessionais semelhantes. Os dendrogramas baseados nos índices de Bray & Curtis e de Morisita-Horn indicam maior similaridade entre os estádios intermediários, enquanto que o dendrograma do índice de Sorensen não indicou uma associação esperada, seja pelo estádio sucessional ou histórico de uso (Figura 2). Fatores que promovem baixa similaridade entre parcelas de amostragem são diferenças de solo, alto índice de exploração em determinada área e diferenças de sítio (OLIVEIRA e ROTTA, 1982). Apesar de o segundo fator ter destaque, o mesmo parece não ter interferido na composição florística desse estrato da vegetação, sendo que a similaridade do solo e relevo parece ser mais decisiva na definição na homogeneidade da vegetação associada, assim como a luminosidade, que também deve ser considerada, pois é fator preponderante no recrutamento de espécies de sub-bosque (TABARELLI e MANTOVANI, 1999). CONCLUSÕES Os resultados indicaram a influência do histórico de uso e dos estádios sucessionais na composição florística e estrutural das áreas de forma diferenciada para as plântulas de espécies arbóreas e para a vegetação associada. Para as espécies arbóreas, A2 apresentou maiores valores para os descritores ecológicos e maior número de espécies e famílias, indicando um maior desenvolvimento do estrato de regeneração do que A1, possivelmente devido ao seu histórico de uso menos intenso. Apesar da alta similaridade florística encontrada entre todos os estádios sucessionais analisados, ficou caracterizado um gradiente de similaridade e possivelmente de sucessão, sugerindo um efeito do histórico de uso nas áreas analisadas em conjunto com o tempo de abandono. Para a vegetação associada, a composição florística mostrou-se muito homogênea entre todos os estádios e áreas analisadas, indicando que essa característica pode ser pouco influenciada pelo histórico de uso após alguns anos de abandono e demonstrando que essas espécies podem ser menos exigentes quanto ao local de recrutamento, respondendo diretamente a similaridades edáficas das áreas analisadas. REFERÊNCIAS BIBLIOGRÁFICAS
Copyright © 2011 - Ciência Florestal The following images related to this document are available:Photo images[cf11041t2.jpg] [cf11041t4.jpg] [cf11041t7.jpg] [cf11041t6.jpg] [cf11041f1.jpg] [cf11041t5.jpg] [cf11041f2.jpg] [cf11041t3.jpg] [cf11041t8.jpg] [cf11041t1.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}