 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
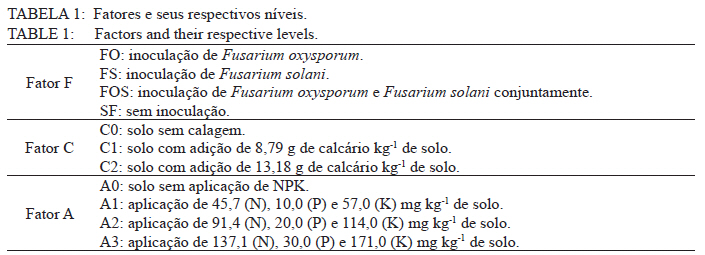
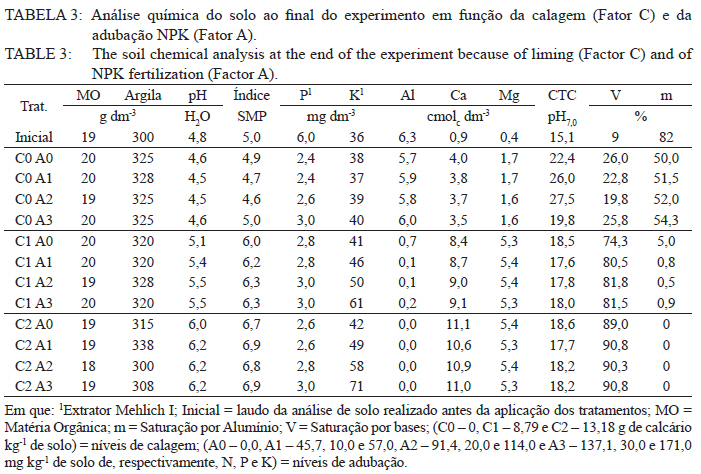
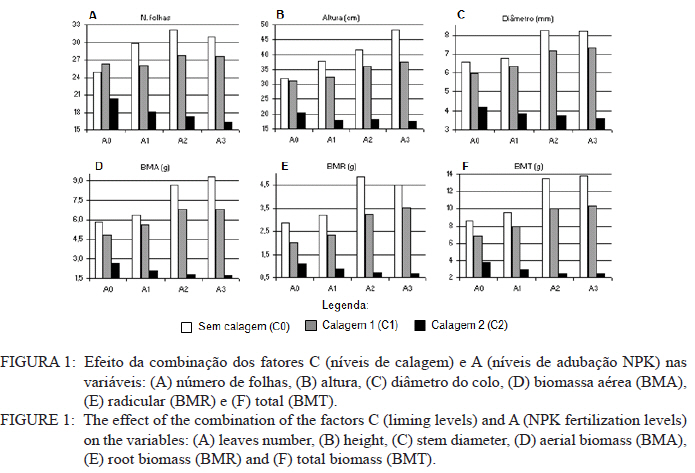
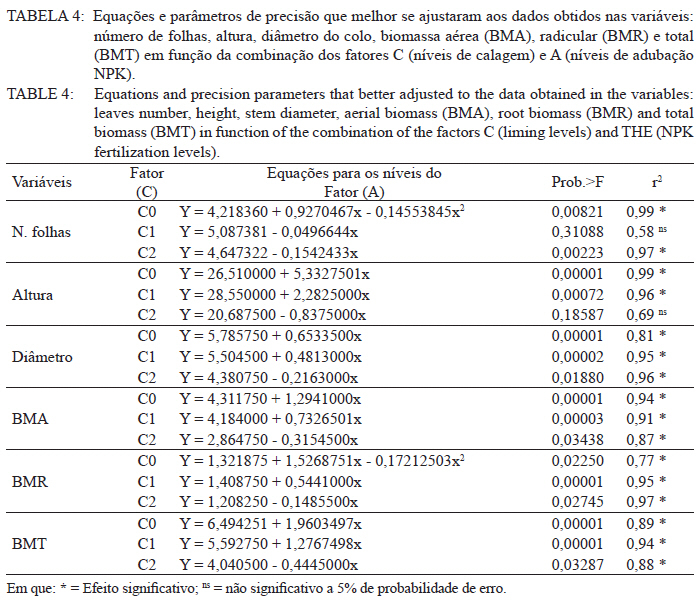
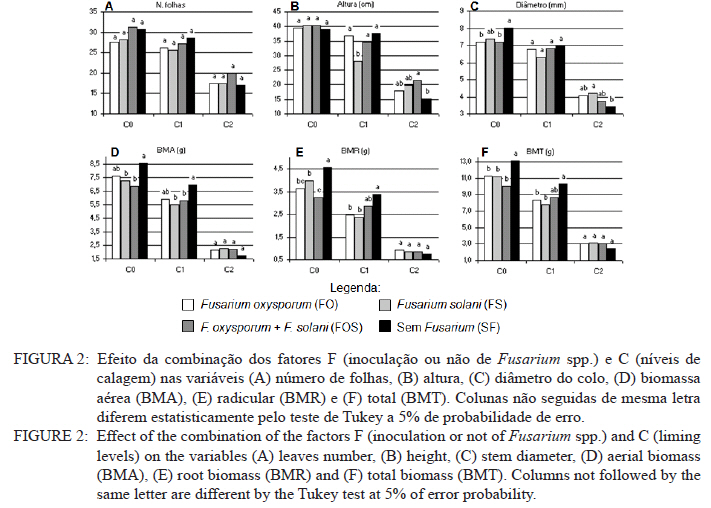
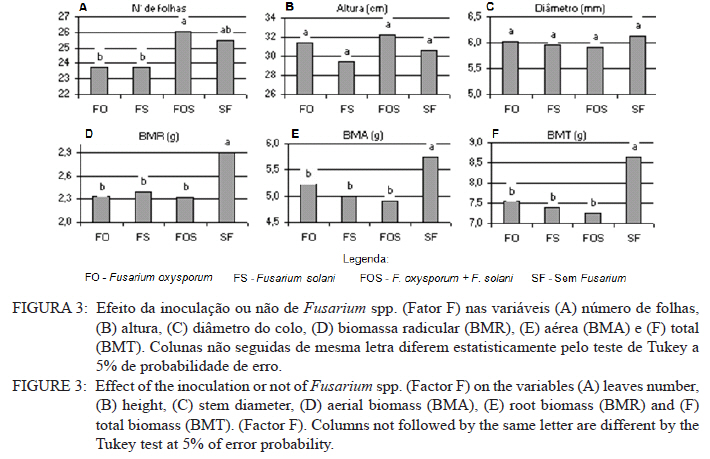
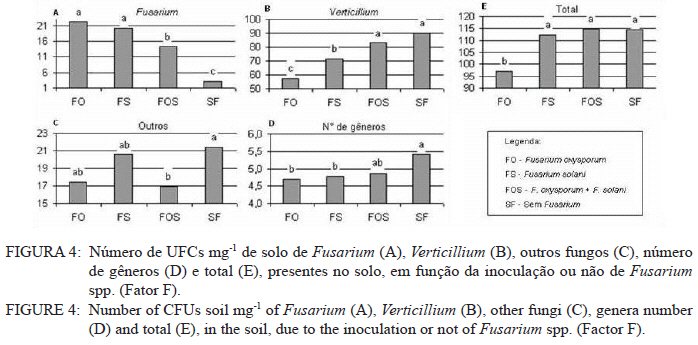
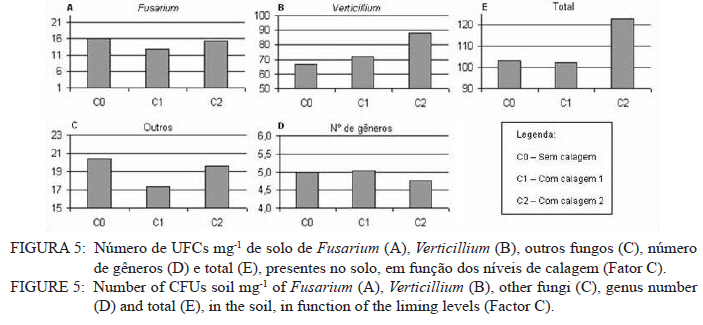
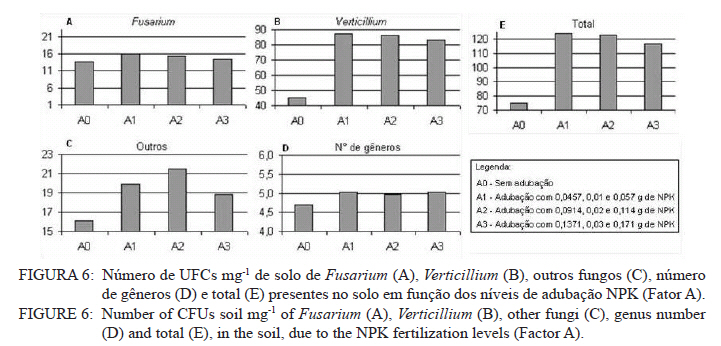
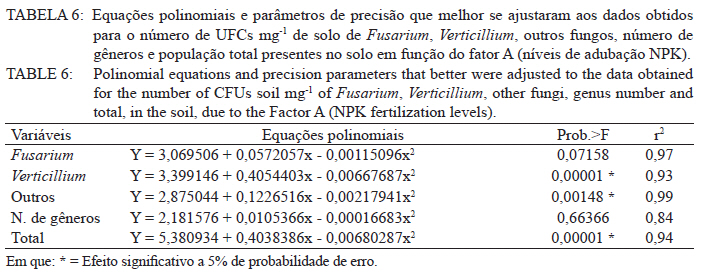
Ciência Florestal, Vol. 21, No. 3, Jul-Sept, 2011, pp. 429-444 Influência da aplicação de NPK e calcário sobre o crescimento da erva-mate, severidade da podridão-de-raízes e população fúngica do solo1 Influence of NPK and lime aplication on erva-mate growth, root-rot severity and soil fungi population Igor Poletto2 Marlove Fátima Brião Muniz3 Denise Ester Ceconi4 Elena Blume5 1. Parte da dissertação de mestrado do primeiro autor apresentada ao Programa de Pós-graduação em Engenharia Florestal, Centro de Ciências Rurais, Universidade Federal de Santa Maria. Recebido para publicação em 5/10/2009 e aceito em 14/09/2010 Code Number: cf11044 RESUMO Este trabalho teve por objetivo avaliar a influência da aplicação de diferentes doses de NPK e calcário sobre o crescimento da Erva-mate, a severidade da Podridão-de-raízes e a população fúngica do solo. Para isso, foi instalado um experimento em casa de vegetação, no Viveiro Florestal da UFSM, utilizando um delineamento experimental inteiramente casualizado, no esquema fatorial 4x3x4 (Fator F: inoculação de Fusarium spp.; Fator C: calagem do solo; Fator A: doses de nitrogênio, fósforo e potássio), totalizando 48 tratamentos. As mudas foram cultivadas em vasos contendo 2 kg de solo, classificado como Argissolo Vermelho-Amarelo. Ao final do experimento foram medidos o diâmetro do colo, a altura a partir do colo, número de folhas, biomassa seca aérea, radicular e biomassa total das mudas. Também foi coletado solo, de cada tratamento, para análise química e contagem da população fúngica. Foi observado que a associação entre adubação NPK e calagem do solo prejudicou o desenvolvimento das mudas de Erva-mate. Pela análise de algumas variáveis sugere-se que a ausência de calagem proporcionou maior resistência às mudas ao ataque de Fusarium spp. ou, a severidade de Fusarium spp. foi diminuída em pH mais baixo. A população fúngica do solo apresentou comportamento variado em função dos tratamentos aplicados. Palavras-chave: Ilex paraguariensis; Fusarium spp.; adubação; proteção florestal. ABSTRACT The present work evaluated the influence of the application of NPK and liming doses in the soil, on the growth of Erva-mate, the severity of rot-root and the fungi population of the soil. To do so, an experiment was installed at the green house, in the Forest Nursery of UFSM, using an experimental design completely randomized factorial 4x3x4 (Factor F: Fusarium spp. inoculation; Factor C: soil limestone; Factor A: NPK doses) , totaling 48 treatments. The seedlings were cultivated in vases containing 2 kg of soil, classified as ‘Red-Yellow Argisoil’ (clay soil). At the end of the experiment was measured the stem diameter, height of the aerial part, leaves number, aerial dry biomass, root dry biomass and total dry biomass of the seedlings. Also, the soil was collected, from each treatment, for the chemical analysis and the counting of the fungi population. It was observed that the association among application of NPK and liming in the soil hampered the development of Erva-mate seedlings. The analysis of some variables suggests that the limestone absence provided greater resistance of seedlings to the attack of Fusarium spp. or the severity of Fusarium spp. was reduced in lower pH. The fungi population of the soil presented varied behavior depending on the applied treatments. Keywords: Ilex paraguariensis; Fusarium spp.; fertilization; forest protection. INTRODUÇÃO A Podridão-de-raízes da Erva-mate (Ilex paraguariensis A. St.-Hil), causada por várias espécies de Fusarium, caracteriza-se por causar destruição do sistema radicular e, por consequência, morte das plantas, em viveiro e em campo. O controle de patógenos de solo, como Fusarium spp., causadores de Murchas-vasculares e Podridão-de-raízes é muito difícil, pois estes se desenvolvem no solo e penetram no hospedeiro via sistema radicular. Além disso, são capazes de sobreviver no solo por longos períodos, o que dificulta sua erradicação (BEDENDO, 1995). Fungicidas químicos não possuem um efeito eficaz em consequência das inúmeras reações químicas que os produtos têm com o solo e também contaminam o meio ambiente e a própria cultura, que muitas vezes é consumida in natura. Neste contexto, há a necessidade de buscar técnicas alternativas de manejo e controle desses patógenos. Na literatura mundial existe um grande número de publicações (HUBER, 1980; MARSCHNER, 1995; ZAMBOLIM et al., 2001; DATNOFF et al., 2007) que tratam da influência dos nutrientes minerais sobre a incidência e a severidade de doenças em culturas agrícolas e florestais. Segundo Huber (1980), todos os elementos minerais essenciais são considerados importantes em relação à incidência ou severidade de doenças. O efeito dos nutrientes em doenças é determinado por: a) efeito da fertilização mineral na severidade da doença; b) comparação das concentrações de elementos nos tecidos de cultivares resistentes e suscetíveis; c) correlação entre condições que influenciam a disponibilidade de minerais com a incidência ou severidade de doenças; e d) combinação dos três efeitos anteriores. Para Zambolim et al. (2001), dentro do triângulo que determina a doença (ambiente-patógeno-hospedeiro), os nutrientes podem afetar direta ou indiretamente o hospedeiro, o patógeno e o meio ambiente, predispondo as plantas ao ataque dos patógenos, atuando direta ou indiretamente; induzindo resistência ou tolerância à planta hospedeira; reduzindo ou aumentando a severidade das doenças; e afetando o ambiente que tanto pode favorecer como desfavorecer os patógenos. Neste contexto, o objetivo deste trabalho foi avaliar a influência da aplicação de diferentes doses de NPK e calcário no solo, sobre o crescimento da Erva-mate, a severidade da Podridão-de-raízes e a população fúngica do solo. MATERIAL E MÉTODOS O experimento foi instalado em casa de vegetação pertencente ao Departamento de Ciências Florestais/UFSM, em condições de ambiente não controlado. O delineamento utilizado foi o inteiramente casualizado no esquema fatorial 4x3x4 (Fator F: inoculação de Fusarium spp.; Fator C: calagem do solo; Fator A: doses de nitrogênio (N) fósforo (P)e potássio (K)), totalizando 48 tratamentos, com cinco repetições de uma muda cada. Os níveis dos fatores foram compostos conforme Tabela 1. Fusarium oxysporum (isolado 12.4a) e Fusarium solani (isolado 4.4) utilizados neste estudo pertencem ao Laboratório de Fitopatologia do Departamento de Defesa Fitossanitária/CCR/UFSM. Estes isolados foram escolhidos pela sua patogenicidade e por estarem presentes em todos os ervais estudados por Poletto et al. (2006). O solo utilizado no experimento foi coletado em um horizonte A (10-20 cm) classificado como Argissolo Vermelho-Amarelo (STRECK, 2002), peneirado (malha 0,5 cm) e seco à sombra. Depois de homogeneizado, foi coletada uma amostra e esta analisada quimicamente no Laboratório Central de Análises de Solo da UFSM. De posse dos resultados (Tabela 2), foram calculadas as doses de N, P, K e calcário a serem aplicadas no solo em cada tratamento. Para a calagem, foi utilizado calcário dolomítico com PRNT de 75,1%. A calagem do nível C1 (Fator C) foi realizada segundo a recomendação da Comissão de Química e Fertilidade do Solo (2004) para a espécie, suficiente para elevar o pH a 5,5 e para o nível C2, suficiente para elevar o pH a 6,0. A adubação realizada no nível A2 (Fator A) foi segundo a recomendação da Comissão de Química e Fertilidade do Solo (2004) para a espécie. A adubação realizada no nível A1 foi 50% a menos da recomendação e a do nível A3 foi 50% a mais. As fontes dos nutrientes N, P e K utilizadas foram, respectivamente, NH4NO2, P2O5 e KCl (PA). Para acondicionar o solo, foram utilizados vasos de polietileno preto com capacidade de 2,5 dm3, vedados para evitar a perda de nutrientes e, em cada vaso, foram adicionados 2 kg de solo seco à sombra. No momento do enchimento dos vasos, o calcário (em pó), o P e o K (diluídos em água) foram incorporados ao solo nos respectivos tratamentos. O N foi adicionado diluído em água duas semanas após a repicagem das mudas. Para determinar a quantidade de água a ser reposta em cada vaso, foi determinada a capacidade de campo do solo segundo o método descrito pela Embrapa (1979). Assim, a água dos vasos foi reposta até 80% da capacidade de campo. Para evitar possíveis influências do meio, os vasos foram alternados a cada vinte dias e pesados para manutenção da umidade na capacidade de campo. Em setembro de 2006 as mudas de Erva-mate foram repicadas para os vasos, todas com o mesmo tamanho (quatro folhas e 5 cm de altura, partindo do colo). Os isolados de Fusarium oxysporum ou Fusarium solani utilizados neste estudo foram oriundos de plantas de Erva-mate doentes e, identificados em estudos anteriores por Poletto et al., (2006), conservados, por dois anos, em frascos com sementes de sorgo e solo esterilizados, em temperatura ambiente. Porções destas sementes foram retiradas dos frascos e transferidas para placas de Petri contendo batata-dextrose-ágar (BDA) e mantidas por 10 dias a 24°C com fotoperíodo de 12 h. Porções das colônias crescidas nas placas foram transferidas para erlenmeyers contendo arroz (sem casca, tipo parboilizado) previamente umedecido e autoclavado (30 min a 121°C a 1 atm). Depois de inoculados, os frascos permaneceram na câmara de incubação a 24°C com fotoperíodo de 12 h por 20 dias. Após esse período, o arroz foi colocado em um recipiente com água esterilizada (1000 mL de água para cada 200 g de arroz) e agitado por 10 minutos, a suspensão resultante foi coada e contou-se o número de esporos, com auxílio da câmara de Neubauer em um microscópio óptico. A suspensão apresentou 4,30 x 107 esporos mL-1 para Fusarium oxysporum e 2,99 x 107 esporos mL-1 para Fusarium solani. Cinco meses depois da repicagem das mudas, foram inoculados 20 mL da suspensão de esporos por vaso, incorporados nos dois primeiros centímetros da superfície do solo e, imediatamente, este foi umedecido com água. Para os tratamentos com inoculação das duas espécies conjuntamente, foram aplicados 10 mL de suspensão para cada espécie. Ao final do experimento (treze meses) foram medidos o diâmetro do colo das mudas com o auxílio de um paquímetro digital (precisão 0,01 cm), a altura a partir do colo com uma régua (precisão 0,01 cm), e realizada a contagem do número de folhas. A parte aérea das mudas foi separada da radicular, embalada em sacos de papel, colocada para secar em estufa de circulação de ar a 70°C até atingir peso constante e, posteriormente, ser pesada em balança analítica de precisão 0,01 g para determinação da biomassa seca aérea (BMA). As raízes foram lavadas em água corrente e embaladas em sacos de papel, secas e pesadas para determinação da biomassa seca radicular (BMR). A biomassa seca total (BMT) foi obtida pela soma das BMA e BMR. Antes da secagem, foi retirada uma amostra do caule e da raiz de cada muda, colocada em câmara úmida em caixas “gerbox” e mantida na câmara de incubação a 24°C com fotoperíodo de 12 h por 6 dias. Após esse período, foi observado o desenvolvimento ou não de Fusarium spp. nas amostras, com auxílio de microscópio estereoscópico. Depois de analisadas, as amostras foram secas, pesadas e o valor obtido somado ao restante do peso da muda. Coletou-se uma amostra de solo de cada uma das 5 repetições que constituíam os tratamentos e, após homogeneização destas, foi retirada uma amostra composta (200 g) para a realização de análise química no Laboratório Central de Análises de Solo da UFSM. Porções de 50 g foram retiradas, da mesma maneira, para a determinação e contagem da população fúngica do solo, e embaladas individualmente em sacos plásticos (dimensões 10 x 15 cm), que foram vedados e armazenados em geladeira a 4°C imediatamente após a coleta até sua análise. Para a determinação da população fúngica, 10 g de solo de cada tratamento foram diluídos em 90 mL de água destilada e esterilizada, juntamente com 2 gotas do emulsificante Tween 80, agitado por 2 min e, partindo dessa suspensão, foram feitas diluições sequenciais de 10-2, 10-3 e 10-4. Em cada diluição foi retirada uma alíquota de suspensão e inoculada em três placas de Petri (0,5 mL placa-1) contendo meio BDA, suplementado com 4,0 mg de estreptomicina (concentração de 70%)/100 mL de meio. As placas inoculadas foram mantidas em câmara de crescimento por 6 dias a 24ºC, com fotoperíodo de 12 h e, após esse período, foram quantificados e identificados os fungos presentes, em nível de gênero, pelas observações das placas diretamente no microscópio estereoscópico e com confecção de lâminas para observação em microscópio óptico, com base na bibliografia especializada (BARNETT e HUNTER, 1999). O número de unidades formadoras de colônia (UFCs) de cada gênero fúngico presente foi determinado através das equações citadas por CARTER (1993). Os dados obtidos nos parâmetros de crescimento foram submetidos à análise da variância para verificação da significância e, em caso positivo, foi aplicada análise de regressão ou comparação de médias com teste de Tukey a 5% de probabilidade de erro. Os dados obtidos da contagem do número de folhas e da população fúngica do solo foram transformados para Raiz (x + k) com k = 0,5 para aproximação da curva normal. Para essas análises, foi utilizado o software estatístico SANEST (ZONTA et al., 1984). RESULTADOS E DISCUSSÃO Efeito dos tratamentos na disponibilidade de nutrientes Na Tabela 3 são apresentados os valores obtidos na análise química do solo, para cada tratamento, ao final do experimento. As características químicas do solo foram interpretadas de acordo com o Manual de Adubação e Calagem para os Estados do RS e SC (COMISSÃO DE QUÍMICA E FERTILIDADE DO SOLO, 2004). Após o crescimento das mudas da Erva-mate, o solo apresentou as características apresentadas na Tabela 3. A matéria orgânica obtida foi considerada “baixa”, antes e após a aplicação dos tratamentos e condução do experimento, com teores em torno de 2%. Segundo Silva et al. (2004), juntamente com os componentes inorgânicos da fase sólida (fração mineral), a matéria orgânica exerce um papel fundamental na química do solo, apresenta uma alta capacidade de interagir com outros componentes, alterando assim propriedades químicas, físicas e biológicas, as quais afetam o crescimento e o desenvolvimento das plantas. O nutriente P, antes da aplicação dos tratamentos, era considerado “baixo” (6,0 mg dm-3) e, ao final do experimento, os teores encontrados foram considerados “muito baixos” (em média 2,8 mg dm-3) em todos os tratamentos, inclusive naqueles nos quais se adicionou o nutriente. Sugere-se como causa o fato de que o solo natural possui baixos teores de P, proporcionando uma maior quantidade de sítios de adsorção nos colóides do solo e, as quantidades do nutriente adicionadas, em função dos tratamentos, foram adsorvidas por estes colóides, ficando o P com baixos teores na solução do solo. Outra explicação possível é a absorção pelas plantas do P que permaneceu disponível. O principal fator que afeta o movimento e disponibilidade do P é o pH e, em solos ácidos, esse elemento fica limitado. Recomenda-se, nesses casos, a utilização de calagem que proporciona o aumento da disponibilidade de P, em consequência da eliminação do alumínio trocável que causa toxidez para as plantas, do aumento da atividade microbiana que provoca maior mineralização do P orgânico e ao aumento da concentração dos íons OH- que podem deslocar o fósforo adsorvido na superfície dos minerais para a solução do solo. Além disso, quando o pH do solo está alto, observa-se uma redução na disponibilidade do nutriente, ou seja, o P tem sua máxima disponibilidade dentro de uma faixa intermediária de pH, considerada ideal para o elemento (ANGHINONI e BISSANI, 2004). O nutriente K, na análise de solo inicial, era considerado “baixo” (36 mg dm-3), ao término do experimento continuou sendo “baixo” (em média 44 mg dm-3), inclusive para o solo dos tratamentos que receberam adição do nutriente, exceto para C1A3 e C2A3, em que os teores encontrados foram considerados “médios” (respectivamente 61 e 71 mg dm-3). De acordo com a Comissão de Química e Fertilidade do Solo (2004), a disponibilidade do K é interpretada de acordo com a Capacidade de Troca de Cátions a pH 7,0 (CTC pH7,0), também chamada de CTC potencial, que é alta no solo, antes e após a aplicação dos tratamentos. Solos com maior CTC retêm mais K na fase sólida, diminuindo as perdas por lixiviação, apresentam também maior capacidade de manter alto o K na solução e, consequentemente, o gradiente de concentração (MEURER e INDA Jr., 2004). Interpretando os resultados obtidos na análise de solo e considerando que, mesmo tendo sido aplicado K, seu teor continuou “baixo” na maioria dos tratamentos, sugere-se como causa a absorção pelas mudas de Erva-mate, pois, nos tratamentos nos quais se aplicou NPK, as plantas cresceram e se desenvolveram melhor e, consequentemente, absorveram maior quantidade de nutrientes, fazendo com que os teores no solo, ao final do experimento, continuassem baixos. Nos tratamentos C1A3 e C2A3, que ao final do experimento apresentaram teores médios de K, a maior adição do nutriente resultou nesse teor final. Os teores de cálcio no solo original foram “baixos” (0,9 cmolc dm-3), e depois da aplicação dos tratamentos passaram a ser “médios” (em média 3,8 cmolc dm-3) para aqueles sem calagem (C0) e “altos” (em média 9,9 cmolc dm-3) para os demais tratamentos (com calagem). Os teores iniciais de magnésio também foram “baixos” (0,4 cmolc dm-3) e após a aplicação dos tratamentos passaram a ser “altos” (em média 4,1 cmolc dm-3). O aumento dos teores de Ca e Mg, mesmo nos tratamentos sem calagem, provavelmente, deve-se a água da irrigação que foi de poço artesiano, normalmente rica nesses nutrientes. Como o experimento teve duração de 13 meses, a quantidade de água adicionada aos vasos foi suficiente para aumentar estes valores. Os valores da saturação de bases (V%) foram “muito baixos” no solo original (9%) e nos tratamentos sem calagem (em média 23,6%), “médio” para o tratamento C1A0 (74,3%) e “altos” para os demais tratamentos (em média 83,4%). Como a V% é obtida, basicamente pela soma dos elementos Ca, Mg e K, seu comportamento foi semelhante ao dos teores desses elementos. O pH em água foi “muito baixo” antes do experimento e para todos os tratamentos C0 (sem calagem) (em média 4,6), “baixo” para C1A0 e C1A1 (em média 5,2), “médio” para C1A2, C1A3 e C2A0 (em média 5,7), e “alto” para os demais tratamentos com C2 (em média 6,2). O aumento do pH do solo em razão da calagem é o efeito esperado por tal prática. A calagem, além de fornecer cálcio e magnésio, aumenta o pH do solo, reduzindo a acidez deste e diminuindo a disponibilidade do alumínio (Tabela 3). À medida que aumenta o pH do solo e os teores de Ca e Mg, diminuem os teores de Al e a saturação por Al (m%). Os valores da saturação por Al no solo, antes da aplicação dos tratamentos e nos tratamentos sem calagem (C0), são “altos” (em média 58%), “baixo” no tratamento C1A0 (5%) e “muito baixo” para os demais tratamentos (em média 0,3%). A saturação por Al nula é um dos principais fatores para se obter boas produtividades, pois as plantas não sofrerão restrições de crescimento por influência da toxidez desse elemento. Os efeitos de toxidez de Al nas plantas podem ser observados principalmente no sistema radicular, reduzindo o desenvolvimento das raízes, causando engrossamento e necrose, que acabam por diminuir a absorção e translocação de P pela planta. Esses distúrbios no sistema radicular das plantas prejudicam o desenvolvimento da parte aérea, que podem apresentar sintomas de deficiência de P (TEDESCO e BISSANI, 2004). Efeito dos tratamentos nos parâmetros de crescimento das mudas Os valores de significância obtidos na análise da variância para os parâmetros de crescimento das mudas mostraram que houve interação significativa somente entre os fatores F x C e C x A. Os fatores isolados F, C e A também apresentaram valores significativos para a maioria das variáveis. Na Figura 1, são apresentados os valores dos parâmetros de crescimento obtidos nas mudas de Erva-mate submetidas à combinação dos fatores C (níveis de calagem) e A (adubação NPK). Para as variáveis altura, diâmetro, biomassa aérea (BMA) e biomassa total (BMT) (Figura 1B, C, D e F), obtiveram-se respostas lineares positivas (Tabela 4) nos níveis C0 e C1 de calagem e linear negativa para C2 em função das doses de adubação. Para os parâmetros de precisão, as equações apresentaram bom ajuste aos dados obtidos, com exceção da variável altura que não foi significativa no maior nível de calagem (C2). Para o número de folhas (Figura 1A), obteve-se resposta quadrática para o nível C0, linear negativa para C1 (não significativa) e linear negativa para C2 (Tabela 4), em função das doses de adubação. A BMR (Figura 1E) teve efeito semelhante, apresentando resposta quadrática para o nível C0, linear positiva para C1 e linear negativa para C2 (Tabela 4). Em todas as variáveis observadas (Figura 1), verificou-se significativa diminuição dos valores com o aumento da calagem. Na média entre variáveis, houve uma diminuição dos valores do nível C0 para C1 de 17% e do nível C0 para C2 de 74%. A maior redução nos valores foi observada para a variável BMR (78%). Esses valores mostram que a calagem do solo foi prejudicial ao desenvolvimento da Erva-mate, sobretudo ao sistema radicular. A Erva-mate habita naturalmente regiões de solos ácidos e apresentam um adequado desenvolvimento em solos com pH na faixa de 4,5 -5,5, sendo a calagem do solo para cultivo da Erva-mate recomendada apenas quando os teores de Ca e Mg estiverem muito baixos (Ca ≤ 2,0 cmolc dm-3 e Mg ≤ 0,5 cmolc dm-3). No entanto, a prática da calagem é comum na cultura da Erva-mate para ajustar o pH à exigência da espécie agrícola, que é geralmente cultivada consorciada. Neste trabalho, optou-se realizar a calagem acima da recomendação para verificar qual seria o efeito sobre o seu crescimento e a sua predisposição ao ataque de Fusarium sp. Analisando conjuntamente as variáveis representadas na Figura 1 e na Tabela 4, os maiores valores foram encontrados no nível sem calagem (C0), em todas as doses de adubação, e efeito contrário para o C2 que, quanto maior a adubação, menor foram os valores observados. Ceconi et al. (2007) estudando diferentes doses de fósforo na produção de mudas de Erva-mate em um Argissolo Vermelho Amarelo, observaram que a máxima eficiência técnica para as variáveis observadas ficou entre 362,5 e 466,25 mg P kg-1 de solo. Resultados semelhantes foram encontrados por Schumacher et al. (2004) na produção de mudas de Angico-vermelho (Parapiptadenia rigida (Benth.) Brenan) e Ceconi et al. (2006) em mudas de Açoita-cavalo (Luehea divaricada Mart.). Fernandes et al. (2000) testaram doses de 0, 150, 300 e 600 mg P kg-1 de solo, classificado como Latossolo, para a produção de mudas de Aroeirinha (Schinus terenbinthifolius Raddi), Paineira (Chorisia speciosa St. Hill.) e Jambolão (Syzygium jambolanum Lam.) e encontraram respostas lineares positivas e quadráticas nas variáveis medidas. Comparando os resultados deste trabalho, com os dos autores acima, pode-se afirmar que a dose de máxima eficiência técnica em P, neste experimento, seria acima da dose máxima testada (30 mg P kg-1 de solo). Nicoloso et al. (2001), estudando a exigência nutricional de mudas de Grápia (Apuleia leiocarpa Vog. Macbride) em um Argissolo Vermelho Distrófico arênico, concluíram que a máxima eficiência técnica estimada da adubação fosfatada e potássica, na produção de massa seca da planta, é acima de 80 mg kg-1 de solo e, para a adubação nitrogenada, é de 70 mg kg-1 de solo e que, o efeito benéfico da adição de N, está condicionado à aplicação conjunta de P. Pandolfo et al. (2003) avaliando a produção de massa verde de Erva-mate a campo, em um Latossolo Vermelho Aluminoférrico, durante cinco anos, sob adubação com doses de 0, 25, 50, 75, 100, 125 g de N por planta por ano, verificaram o efeito quadrático na produção acumulada nos 5 anos, com máxima eficiência técnica de 80,5 g de N por planta por ano, produzindo 36,5 kg de massa verde por planta. Efeito positivo na aplicação de N também foi observado por Marques et al. (2006). Os autores observaram efeito significativo na aplicação de N na produção de mudas de Dalbergia nigra (Vell.) Fr. All. Ex Benth., em um Argissolo Vermelho Amarelo. Eles recomendam que a dose de 180 mg N kg-1 de solo é a melhor para a produção de mudas da espécie. Lourenço et al. (1998) estudaram a aplicação de diferentes doses de NPK (20-5-20), na produtividade de um erval, no município de Áurea, RS. Os tratamentos consistiram na aplicação de 0, 170, 340 e 510 g planta-1. Nas medições efetuadas após dois anos de aplicação, os autores observaram resposta quadrática para as variáveis altura e biomassa, sendo os maiores valores obtidos nas doses de, respectivamente, 170 e 340 g planta-1. Reissmann e Carneiro (2004), testando cinco níveis de calagem do solo, durante 8 anos seguidos, observaram que as plantas de Erva-mate não apresentaram diferenças significativas entre tratamentos, nas variáveis altura, diâmetro da copa e produção de biomassa, ficando o tratamento testemunha - sem calagem (11,6% de saturação de bases), com os maiores valores. Portanto, analisando os trabalhos encontrados na literatura sobre adubação com NPK em espécies florestais, juntamente com os resultados encontrados neste estudo, pode-se afirmar que as doses desses nutrientes aplicadas à Erva-mate, neste experimento, não foram suficientes para prejudicar seu crescimento, exceto quando a adubação foi realizada conjuntamente com a calagem do solo. Na Figura 2, estão representados os valores obtidos nos parâmetros de crescimento das mudas de Erva-mate submetidas à combinação dos fatores F (inoculação de Fusarium spp.) e C (níveis de calagem). Para a variável número de folhas (Figura 2A), não houve diferença significativa entre os níveis dos fatores. Observando-se separadamente as variáveis número de folhas e altura (Figura 2A e B) pode-se inferir que: a ausência de calagem (C0) proporcionou maior resistência às mudas ao ataque de Fusarium spp., igualando-se a testemunha (SF); a severidade de Fusarium spp. foi diminuída em pH mais baixo; ou estas variáveis não refletem os efeitos causados pelo ataque de Fusarium spp. Para as outras variáveis houve, na maioria das vezes, superioridade da testemunha (SF) em relação aos níveis com presença de Fusarium. No nível de calagem C1 (Figura 2), observou-se superioridade nos valores obtidos para a testemunha (SF) em relação aos demais, mesmo que para algumas variáveis, a análise estatística estabeleceu igualdade entre os níveis. O Fusarium solani (nível FS) apresentou os menores valores em todas as variáveis e, com isso, pode-se afirmar que esse patógeno é mais severo nesta calagem ou então, que as mudas tornaram-se mais suscetíveis, provavelmente, por alterações fisiológicas e bioquímicas. Analisando a resposta dos níveis com e sem inoculação de Fusarium, em relação ao nível C2 de calagem (Figura 2), a testemunha (SF) apresentou os menores valores das variáveis analisadas em comparação aos níveis inoculados (FO, FS e FOS), embora a análise estatística não tenha estabelecido diferenças para algumas variáveis. Isso, provavelmente, pode ser atribuído à diminuição da área de absorção de raízes, causada pelo ataque de Fusarium spp., impedindo a absorção e ficando menos expostas ao excesso de nutrientes. Embora o efeito da inoculação de Fusarium spp. tenha proporcionado a manifestação de sintomas severos (POLETTO et al., 2010), o efeito sobre os parâmetros de crescimento não foi tão pronunciado. Neste caso, os treze meses de condução deste experimento podem ter sido insuficientes para a manifestação mais severa do patógeno em função dos tratamentos aplicados. Os trabalhos publicados nos principais periódicos, nacionais e internacionais, envolvendo patógenos radiculares ou da parte aérea, na maioria, não mencionam valores de variáveis como altura e produção de biomassa e, sim, notas quantificando a incidência e a severidade dos sintomas ocasionados pelos patógenos, dificultando a comparação dos resultados obtidos no presente trabalho com outros estudos sobre o mesmo assunto. Na Figura 3, encontram-se os gráficos dos valores obtidos nos parâmetros de crescimento para o fator F (inoculação de Fusarium spp.) e a comparação de médias pelo teste de Tukey. Notou-se que para a variável número de folhas (Figura 3A) não houve diferença significativa entre os níveis FO (Fusarium oxysporum), FS (Fusarium solani) e SF (sem Fusarium), com média de 24 folhas. O nível FOS (Fusarium oxysporum + Fusarium solani) não diferiu do SF, mas diferiu do FO e FS. A altura e o diâmetro (Figura 3A e B) não tiveram diferenças significativas, permanecendo com médias de, respectivamente, 30,9 cm e 6,0 mm. As variáveis BMR, BMA e BMT (Figura 3D, E e F) foram fortemente influenciadas pela presença de Fusarium. Os níveis FO, FS e FOS foram iguais estatisticamente, diferindo da testemunha (SF) que permaneceu com os maiores valores. Em média, os valores de BMT dos níveis inoculados foram 14% menores que os não inoculados. Nota-se que a inoculação de Fusarium spp. proporcionou significativa diminuição do sistema radicular, provavelmente, por impedimento no crescimento ou por necrose, o que provocou a diminuição da BMA (Figura 3E), pela menor área de absorção de nutrientes e/ou por obstrução dos vasos condutores de seiva. A redução do sistema radicular de mudas, segundo Carneiro (1995), pode ocorrer por diversos motivos como: a composição física e química do substrato, a umidade, o sombreamento e, sobretudo por ataque de patógenos radiculares. A reduzida proporção de raízes, em relação à parte aérea compromete, de várias formas, a qualidade das mudas, interferindo diretamente na sobrevivência e desenvolvimento a campo. A significativa redução na produção de biomassa aérea e radicular, nos níveis inoculados com Fusarium spp., mostra o potencial desse patógeno em causar danos à Erva-mate. Efeito dos tratamentos na população fúngica do solo A população fúngica do solo foi composta pelos gêneros Fusarium, Verticillium, Penicillium, Aspergillus, Mucor, Paecilomyces, Trichoderma, Nigrospora, Cladosporium, Rhizopus, Rhizoctonia e Dichobotrys. A população de Fusarium e Verticillium foi analisada estatisticamente, pois esses fungos apresentaram-se em maior número e em todos os tratamentos. As demais foram somadas compondo a variável “outros”, a variável “Nº de gêneros” foi composta pela contagem do número de gêneros presentes em cada tratamento e a variável “total” foi a soma do número de UFCs de todos os fungos presentes. Com exceção do Fusarium e Rhizoctonia, causadores de Podridão-de-raízes, os demais fungos presentes no solo não são considerados patógenos de podridões radiculares da Erva-mate, porém, podem associar-se à planta de maneira oportunista, como organismos secundários na decomposição dos tecidos mortos (BARNETT e HUNTER, 1999; AUER e GRIGOLETTI Jr., 2002; POLETTO et al., 2007). Na análise de variância para população fúngica do solo, houve efeito significativo para todas as variáveis no fator F e na interação entre F*C. A população de Fusarium e o número de gêneros fúngicos foram influenciados significativamente somente pelo fator F e na interação entre F*C. As populações de Verticillium e total foram influenciadas significativamente, por todos os fatores isolados e interações duplas. Para a interação tripla, somente a variável Verticillium apresentou significância, por este motivo, não foi abordada neste estudo. Embora verificadas diferenças estatísticas para a maioria das variáveis, pelo teste de Tukey, não foi possível obter uma explicação lógica para o comportamento destas, em função da interação dos fatores F e C (inoculação ou não de Fusarium x níveis de calagem) e a interação F e A (inoculação ou não de Fusarium x níveis de adubação). Na interação entre os fatores C (níveis de calagem) e A (níveis de adubação) somente as variáveis Verticilium, outros e total apresentaram significância para a interação. Como a variável Fusarium, objeto principal de estudo deste trabalho, não apresentou significância, esta interação não se torna tão interessante, então, optou-se em não apresentar os dados. Na Figura 4, observou-se os gráficos do número de UFCs mg-1 de solo da população fúngica em função do fator F (inoculação ou não de Fusarium spp.), juntamente com o teste de Tukey. A população de Fusarium foi maior nos níveis FO e FS (21 UFCs, em média), porém, quando Fusarium oxysporum e Fusarium solani foram inoculados conjuntamente (FOS) houve uma significativa redução (33%) no número de UFCs, mesmo que a severidade de alguns sintomas nas mudas de Erva-mate foi maior neste nível (POLETTO et al., 2010). Uma hipótese, para isso, é que as duas espécies podem competir entre si por espaço, causando uma menor produção de esporos. No nível SF, foram encontradas cerca de 3 UFCs mg-1 de solo, uma quantidade significativamente menor que o encontrado nos demais. Esse número se refere, provavelmente, à população natural de Fusarium presente no solo utilizado. O menor número de UFCs presente no nível FS em relação ao FO, embora não significativo, pode ser atribuído ao menor número de esporos inoculados no solo nesse nível durante a instalação do experimento (4,30 x 107 esporos mL-1 para Fusarium oxysporum e 2,99 x 107 esporos mL-1 para Fusarium solani), ou a característica natural da espécie em produzir menos. Diferentemente de Fusarium, a população de Verticillium (Figura 4B) foi maior nos níveis SF e FOS (respectivamente 90 e 83 UFCs) e menor no FO (57 UFCs). Parece haver uma relação inversa entre as duas populações, quanto menor a população de Fusarium maior a população de Verticillium. Nesse caso, pode ter ocorrido uma competição por espaço e nutrientes do solo por esses dois fungos, uma população ocupando o espaço da outra, ou então, uma prejudicando o desenvolvimento da outra, suprimindo-a. Segundo Wollum (1998), no solo existe uma grande competição entre organismos por nutrientes, espaço e umidade. A população dos demais fungos (Figura 4C) foi maior nos níveis SF e FS e menor nos demais níveis. O N° de gêneros (Figura 4D) foi significativamente maior no nível SF, em comparação com os demais. A população total de fungos foi significativamente igual entre os níveis FS, FOS e SF, com média de 114 UFCs mg-1 de solo. Já o nível FO apresentou 97 UFCs mg-1 de solo, número significativamente menor que os demais. A calagem (Fator C) não influenciou significativamente a população de Verticillium (Figura 5B), embora apresentasse uma tendência de resposta quadrática (Tabela 5) ao aumento de pH do solo. Como a população de Verticillium é maior que as demais, acabou influenciando a população total (Figura 5E) que apresentou comportamento quadrático (Tabela 5) ao aumento de pH. Monteiro et al. (2004) observaram em meio de cultura, que o crescimento e a esporulação de Verticillium lecanii não foram influenciados significativamente pelos diversos níveis de pH testados (5, 6, 7, 8 e 9), com exceção do pH 4 que reduziu seu crescimento. A população de Fusarium (Figura 5A) foi influenciada significativamente pelo pH do solo, apresentando uma tendência de resposta quadrática nos valores obtidos (Tabela 5). O número de UFCs nos níveis C0 e C2 foram semelhantes (respectivamente 16 e 15 UFCs mg-1 de solo), porém, houve uma redução de aproximadamente 20% desses para o C1, podendo indicar que este nível de calagem, recomendado pela Comissão de Química e Fertilidade do Solo (2004) para a Erva-mate, exerce uma influencia positiva para a diminuição da população de Fusarium. Efeito semelhante também foi observado na soma da população dos demais fungos (Figura 5C), respondendo de forma quadrática (Tabela 5), obtendo os menores valores no nível C1. O número de gêneros (Figura 5D) não foi influenciado significativamente pela calagem. Segundo Brandão (1992), os fungos podem ser encontrados em solos com pH de 3,0 a 9,0, porém, o valor ótimo varia de espécie para espécie. A inibição do crescimento microbiano, em valores de pH considerados desfavoráveis, resulta não só do efeito direto da elevada concentração de H+ ou de OH-, mas também da influência direta do pH na penetração de compostos tóxicos, presentes no meio, nas células microbianas. A ação do pH sobre os microrganismos do solo depende de sua tolerância a esse fator (TSAI et al., 1992). Para Maffia e Mizubuti (2005), dentre as características químicas do solo, o pH e os nutrientes são os que mais afetam a atividade microbiana e os microrganismos respondem diferentemente a amplitudes do pH e fungos, em geral, crescem melhor em pH mais ácido. Portanto, a menor população de Fusarium (20%) no nível C1 de calagem (5,4), neste estudo, em relação aos demais níveis, pode sinalizar que houve um desfavorecimento no desenvolvimento desse fungo. A resposta de fungos à calagem do solo também foi observada por Rodrigues et al. (2002). Os autores estudaram a influência de doses de calcário na severidade da podridão-radicular do feijoeiro causada por Rhizoctonia solani e observaram resposta linear positiva no índice de doença (ID) em função da calagem. Houve uma variação no ID de 57% para 87% quando o pH foi aumentado de 4,9 para 5,8. Os níveis de adubação NPK (Fator A) não influenciaram significativamente a população de Fusarium no solo (Figura 6A) embora tenha apresentando uma tendência de resposta quadrática (Tabela 6). A disponibilidade de nutrientes, assim como a capacidade de sua utilização pelos diferentes organismos do solo, tem correlação direta com o desenvolvimento de algumas doenças do sistema radicular. Patógenos que crescem numa faixa ampla de fonte de carbono e utilizam compostos nitrogenados orgânicos e inorgânicos podem ter vantagens adaptativas em diferentes tipos de solo (PUNJA, 1985 apud MAFFIA e MIZUBUTI, 2005). A população de Verticillium (Figura 6B) respondeu de forma quadrática à adubação (Tabela 6), com um aumento de 48,5% do nível A0 para o A1 e diminuindo cerca de 5%, partindo desse. Com esses resultados, pode-se afirmar que o maior desenvolvimento deste gênero fúngico está condicionado a uma maior disponibilidade de nutrientes no solo. A maior população de Verticillium perante as demais acabou influenciando os resultados da soma total (Figura 6E), que apresentou o mesmo comportamento. A população dos demais fungos (Figura 6C) apresentou resposta quadrática (Tabela 6) às doses de adubação, sendo o maior valor (21,5 UFCs mg-1 de solo) observado no nível A2. O número de gêneros (Figura 6D) não foi significativo estatisticamente. O reduzido número de UFCs total, observado no nível sem adubação (A0), perante os demais (Figura 6E), indica que a população fúngica do solo é fortemente comprometida pela baixa disponibilidade de nutrientes, existindo uma quantidade que, para mais ou para menos, é ideal para seu desenvolvimento, já que, nas maiores doses de NPK (A2 e A3), se observa uma tendência de diminuição do número de UFCs no solo. A quantificação do inóculo é fundamental para a maioria dos trabalhos que envolvem patógenos radiculares, sobretudo, em estudos epidemiológicos. As possibilidades de infecção de plantas por patógenos do sistema radicular relacionam-se à quantidade de inóculo presente. Essa relação é mais importante para patógenos do sistema radicular que para os da parte aérea, em vista da baixa capacidade de redistribuição dos primeiros, os quais normalmente causam doenças monocíclicas (MAFFIA e MIZUBUTI, 2005; MICHEREFF et al., 2005). A população de Fusarium estabelecida ao final do experimento nos tratamentos inoculados, em média, foi de 18,76 UFCs mg-1 de solo, enquanto que nos tratamentos não inoculados, a média foi de 3 UFCs mg-1 de solo. Menezes (2007) analisando a população fúngica presente no solo cultivado com crisântemo, em casa de vegetação, encontrou 14,6 UFCs mg-1 de solo de Fusarium, 17,2% do total. Essa população foi suficiente para causar sérios danos à cultura de crisântemo, ocasionada pela doença de murcha-vascular. Portanto, a quantidade de inóculo no solo, do presente estudo, foi suficiente para o estabelecimento da doença e, que a quantidade e o método de inoculação foram adequados para o estabelecimento do Fusarium. A maior população presente foi do gênero Verticillium, em média, 69% do total, 75,5 UFCs mg-1 de solo. Uma hipótese para justificar a grande quantidade desse fungo, seria a de que ele é um habitante natural e abundante no solo utilizado neste experimento. As populações dos demais fungos somaram, em média, 19,1 UFCs mg-1 de solo, perfazendo 17,5% do total presente, que foi de 109,43 UFCs mg-1 de solo. A população fúngica encontrada neste estudo foi acima da encontrada por Menezes (2007) em solo com cultivo de crisântemo, em casa de vegetação, que foi de 84,7 UFCs mg-1 de solo. O resultado encontrado neste estudo foi, provavelmente, influenciado pela população de Verticillium. Nas amostras de raízes e parte aérea, colocadas em câmara úmida, observou-se que em 100% dos tratamentos inoculados ocorreu crescimento de Fusarium, partindo dos tecidos internos das mudas, após seis dias de incubação. Isso mostra que embora a Erva-mate tenha apresentado, em alguns tratamentos, resistência ao patógeno, não conseguiu impedir a penetração e a colonização. Nos tratamentos sem inoculação, não houve desenvolvimento de Fusarium. CONCLUSÕES A associação entre adubação NPK e calagem do solo prejudica o desenvolvimento das mudas de Erva-mate. O melhor desenvolvimento das mudas é observado nos tratamentos com adubação NPK sem calagem do solo. Pela análise de algumas variáveis sugere-se que a ausência de calagem proporciona maior resistência às mudas ao ataque de Fusarium spp. ou, a severidade de Fusarium spp. é diminuída em pH mais baixo. A inoculação de Fusarium spp. proporciona significativa redução da biomassa aérea e de raízes das mudas. A população fúngica do solo apresenta comportamento variado em função dos tratamentos aplicados. REFERÊNCIAS BIBLIOGRÁFICAS
Copyright © 2011 - Ciência Florestal The following images related to this document are available:Photo images[cf11044t5.jpg] [cf11044t1.jpg] [cf11044t2.jpg] [cf11044f6.jpg] [cf11044t4.jpg] [cf11044f3.jpg] [cf11044f1.jpg] [cf11044t6.jpg] [cf11044t3.jpg] [cf11044f4.jpg] [cf11044f2.jpg] [cf11044f5.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}