 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
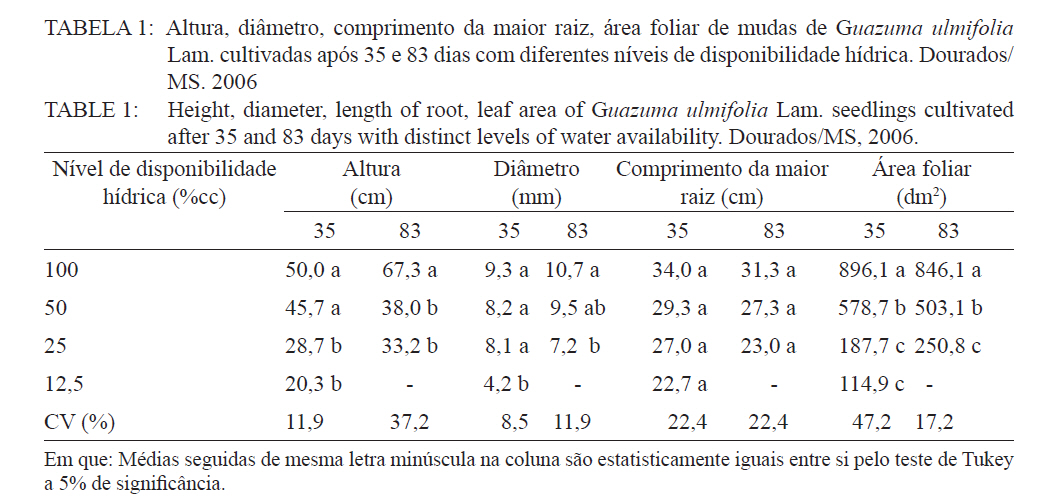
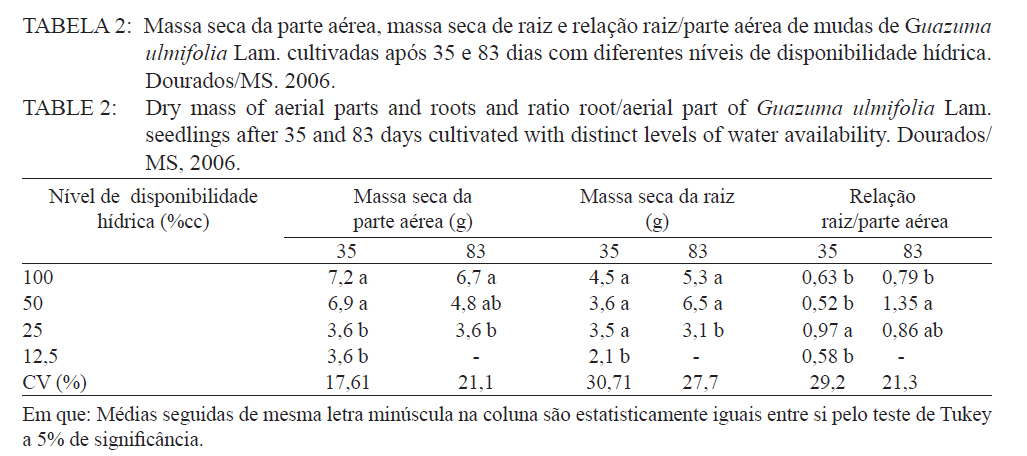
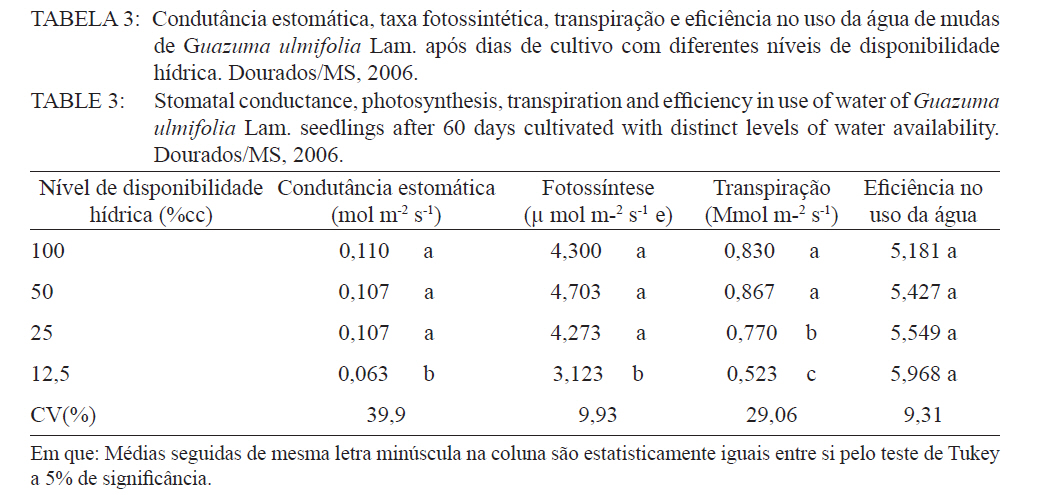
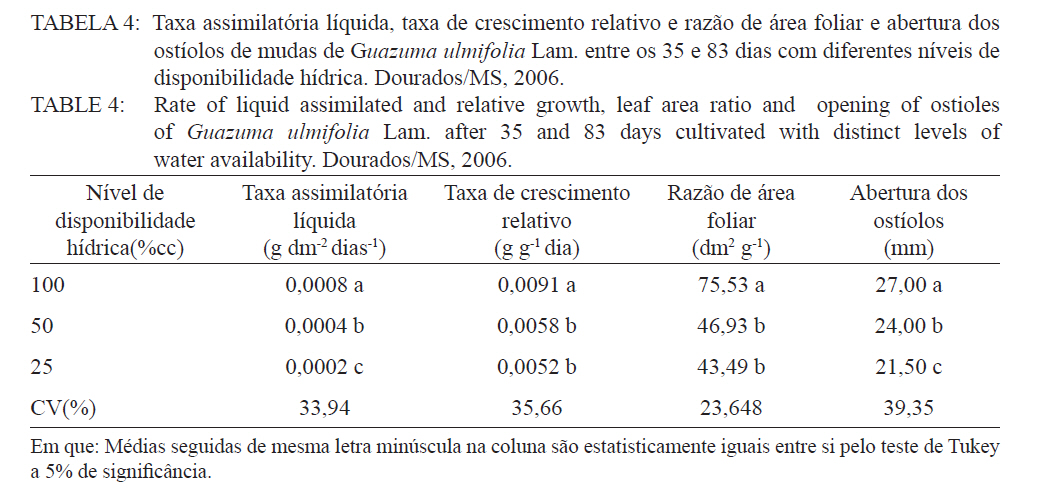
Ciência Florestal, Santa Maria, v. 21, n. 4, p. 655-662, out.-dez., 2011 ESTRESSE HÍDRICO NO METABOLISMO E CRESCIMENTO INICIAL DE MUDAS DE MUTAMBO (Guazuma ulmifolia Lam.) WATER STRESS IN METABOLISM AND INITIAL GROWTH OF MUTAMBO (Guazuma ulmifolia Lam.) SEEDLINGS Silvana de Paula Quintão Scalon1 Rosilda Mara Mussury2 Verusca Luzia de Mello Euzébio3 Flavia Mitsuko Kodama3 Camila Kissmann4 1Bióloga, Drª., Professora Adjunta da Universidade Federal da Grande Dourados, Rodovia Dourados Itahum,
Km 12, CEP 79804970, Dourados (MS). Bolsista de produtividade CNPq. silvanascalon@ufgd.edu.br /
maramussury@ufgd.edu.br Recebido para publicação em 20/05/2009 e aceito em 20/08/2010 Code Number: cf11064 RESUMO Este trabalho teve como objetivo avaliar o crescimento inicial das mudas de Guazuma ulmifolia (mutambo) sob diferentes regimes hídricos. As mudas receberam os seguintes tratamentos de água: 100%, 50%, 25% e 12,5% da capacidade de campo e foram avaliadas aos 35 e 83 dias de cultivo nos diferentes tratamentos. A sobrevivência das mudas a 12,5%cc foi menor que nos demais tratamentos, podendo ser avaliadas apenas aos 35 dias. A altura, diâmetro, área foliar, massas seca de parte aérea e raiz foram maiores sob maior disponibilidade de água, entretanto o comprimento médio das raízes não variou entre os tratamentos. A condutância estomática, fotossíntese e transpiração foram menores sob 12,5%cc, porém a eficiência no uso da água não variou entre as diferentes cc. A taxa assimilatória líquida, taxa de crescimento relativo, razão de área foliar e a abertura dos ostíolos diminuíram com as menores cc. Para a produção de mudas de Guzuma ulmifolia o regime hídrico na fase inicial deve ser de 100% da capacidade de campo, porém as mudas crescem satisfatoriamente sob 50%cc. Abaixo de 25%cc as mudas entram em estresse hídrico antes dos 35 dias de cultivo. Palavras-chave: ecofisiologia; déficit hídrico; árvore nativa. ABSTRACT This work aimed to evaluate the initial growth of Guazuma ulmifolia (mutambo) seedlings under different water regimen. The seedlings received the following treatments of water: 100%, 50%, 25% and 12.5% of the capacity of field and were evaluated in 35 and 83 days of cultivation in different treatments. The seedlings survival in 12.5%cc was smaller than in others treatments, being evaluated only on 35 days after the seedlings did not resist, lost completely the leaf and died. The height, diameter, the leaf area, dry mass of airy part and roots were higher under high water availability, however the medium length of the roots did not vary among the treatments. The stomatal conductance, photosynthesis and transpiration rates were lower under 12.5%cc, but the efficiency in the use of water did not vary between the different cc. The rate of liquid assimilated and relative growth, leaf area ratio and opening of ostioles decreased with low field capacity. To the production of seedlings of Guzuma ulmifolia the water regimen in initial phase must be of 100%cc, however the seedlings growth is satisfactory below 50%cc. Bellow of 25%cc the seedlings get in water stress before the 35 days of cultivation. Keywords: eco-physiology; deficit water; native tree. INTRODUÇÃO Muitas espécies arbóreas nativas são potencialmente aptas para o cultivo, podendo servir para diversos fins quer seja pelo valor ornamental, madeireiro, alimentício ou de preservação. Dessa forma, a melhoria do sistema de produção de mudas de espécies arbóreas nativas é necessária devido ao aumento na procura por tais espécies para a produção comercial, bem como para a recuperação de áreas degradadas (NIETSCHE et al., 2004). A regeneração natural das florestas tropicais é afetada pelas condições ambientais presentes durante o estabelecimento das plântulas e pela capacidade destas em resistir a estresses. Nesses ambientes, a disponibilidade de luz, de água e de nutrientes varia no tempo e espaço e são considerados os fatores abióticos que mais afetam o estabelecimento das plântulas (EVANS e EDWARDS, 2001). Em condições de baixa disponibilidade de água no solo, vários processos metabólicos das plantas podem ser influenciados, como o fechamento estomático, redução da condutância estomática, redução da fotossíntese e transpiração, levando ao declínio da taxa de crescimento (SILVA et al., 2002; GOMES et al., 2004; PORTES et al., 2006). Silva et al. (2009) observaram que plantas cultivadas em ambientes áridos e semiáridos por diversos pesquisadores, estão expostas a longos períodos de déficit de água no solo e desenvolveram adaptações para tolerar a seca. A redução na taxa fotossintética associada com fechamento estomático devido a mudanças no status de água da folha, foi comumente observada levando à redução da transpiração e da possibilidade de morte por dessecação. Por outro lado, a saturação hídrica do solo pode afetar o crescimento de raízes e também da parte aérea das plantas, tanto pela inibição do alongamento dos entrenós, quanto pela inibição da expansão das folhas, podendo também acelerar a senescência e abscisão destas. Nessa condição, a dinâmica da física do solo é alterada de maneira significativa pela saturação hídrica, uma vez que diminui os espaços aeríferos do solo limitando as trocas gasosas com a atmosfera, trazendo como consequência, alterações no metabolismo das células radiculares (LARCHER, 2000; TAIZ e ZEIGER, 2004, BATISTA et al., 2008). São escassos os trabalhos sobre a tolerância ou a adaptação de espécies arbóreas nativas a condições de estresse hídrico. A análise de crescimento permite avaliar o crescimento final da planta como um todo e, a partir dos dados de crescimento, pode-se inferir fisiologicamente, estimando de forma bastante precisa, as causas de variações de crescimento entre as plantas geneticamente diferentes ou entre plantas semelhantes crescendo em ambientes diferentes (LARCHER, 2000). A Guazuma ulmifolia Lam. Sterculiaceae é uma árvore de 8 a 16 m de altura, conhecida popularmente por mutambo, fruta de macaco, embireira, embiru, pau-de-pomba, mutamba-verdadeira, pau-de-bicho. Apresenta pequenas flores amarelas, reunidas em cimeiras axilares, com muitos estames. Fruto globoso, deiscente, com epicarpo dotado de excrescência espinhosa com muitas sementes pequenas e esbranquiçadas. Floresce de setembro a novembro. Ocorre desde a Amazônia até o Paraná, comum nas orlas de cerradão, no Pantanal Mato-Grossense e às margens de pequenos cursos de água (BRANDÃO et al., 2002; CARVALHO, 2006). A madeira é empregada nas confecções de tonéis, coronhas de armas, construções internas, caixaria e pastas celulósicas. O lenho produz ótimo carvão que pode ser transformado em pólvora de excelente qualidade. A casca fornece material para fabricação de cordas. A árvore apresenta bela copa que proporciona ótima sombra podendo ser utilizada com sucesso em paisagismo em geral. Produz anualmente grande quantidade de sementes viáveis (LORENZI, 2002). Apesar das informações disponíveis, não foram encontrados na literatura consultada trabalhos sobre a ecofisiologia dessa espécie. Diante do exposto, este trabalho teve como objetivo avaliar o crescimento inicial das mudas de Guazuma ulmifolia (mutambo) sob diferentes regimes hídricos. MATERIAL E MÉTODOS O experimento foi realizado no período de 2005 a 2006, no Horto de plantas medicinais da UFGD (Universidade Federal da Grande Dourados) na região de Dourados, Mato Grosso do Sul. Os frutos de Guazuma umifolia Lam. foram colhidos em diversas matrizes na mesma região. A semeadura ocorreu em bandejas de células contendo substrato Plantmax®, e quando as plântulas alcançaram 15 cm de altura elas foram transplantadas para embalagem de plástico com capacidade para 3 kg contendo o substrato terra + areia + cama de frango semidecomposta (1:1:1), mantidas em sombrite 50%. A cama de frango era composta de esterco de frango e casca de arroz. Após 10 dias de aclimatação, as mudas foram mantidas em casa de vegetação e receberam o seguinte regime de água: 100, 50, 25 e 12,5% da capacidade de campo (cc), calculada com base na densidade do solo empregado. Aos 35 e 83 dias de tratamentos (185 e 234 dias após a semeadura) as mudas foram avaliadas quanto à altura do caule e ao diâmetro do colo e 3 mudas de cada tratamento e repetição foram destruídas para quantificar a massa seca e fresca das raízes e da parte aérea (g), o comprimento da raiz (cm) e área foliar (dm²), e com os dados de massa seca de raiz e parte aérea foi calculada a razão raiz/parte aérea. Foi calculada também a taxa assimilatória líquida (TAL g dm-2 dia-1), a taxa de crescimento relativo (TCR g g-1 dia), a razão de área foliar (RAF dm2 g) segundo Benincasa (2003), além da abertura dos ostíolos (mm), realizada com auxílio de microscópio estereoscópio. Aos 60 dias de tratamento foram avaliados também a condutância estomática, a taxa fotossintética e a transpiração com auxílio do medidor de fotossíntese LCi portátil. A eficiência no uso da água foi calculada com base na razão entre a taxa fotossintética e a taxa de transpiração. O experimento foi realizado em delineamento inteiramente casualizado com quatro repetições de doze mudas. Os resultados foram submetidos à análise de variância e as médias comparadas pelo teste de Tukey a 5% de significância (BANZATTO e KRONKA, 2003). RESULTADOS E DISCUSSÃO A sobrevivência das mudas de mutambo em condição de menor disponibilidade hídrica (12,5%cc) foi menor que nos demais tratamentos, podendo ser avaliada apenas aos 35 dias de tratamentos com regimes hídricos diferenciados após os quais as mudas não resistiram, perdendo completamente as folhas e secando. Nos demais tratamentos, as mudas puderam ser observadas até 83 dias de tratamento quando se encontravam com boa aparência. Na primeira avaliação realizada aos 35 dias de tratamento, a altura das mudas foi maior com a maior disponibilidade de água, não variando entre os níveis de 50 e 100% cc. Nos tratamentos de menor disponibilidade hídrica, observou-se que a altura foi cerca de 50% menor que nos demais tratamentos (Tabela 1). Na segunda avaliação, aos 83 dias de tratamento, a altura foi maior sob 100%cc, observando-se também uma redução de 50% nas mudas sob menor disponibilidade hídrica. O diâmetro das mudas cultivadas com 12,5%cc aos 35 dias foi metade dos demais tratamentos que não variaram entre si, na segunda avaliação aos 83 dias, o diâmetro foi menor a 25%cc. O comprimento da maior raiz principal não variou entre as diferentes disponibilidades hídricas e épocas de avaliação, entretanto a área foliar foi maior com a maior disponibilidade hídrica em ambas as avaliações. Na primeira avaliação a área foliar das mudas cultivadas com 100%cc foi de 400 a 800% maior que naquelas sob menores disponibilidades hídricas (25%cc e 12,5%cc respectivamente) e na segunda avaliação foi 300% maior que aquela observada nas mudas cultivadas a 25%cc. Resultado semelhante foi observado para Myracrodruon urundeuva Allemão que não apresentou diferença significativa do comprimento radicular entre os tratamentos, porém a área foliar foi menor sob 25%cc. Nessa espécie, a área foliar não variou entre os tratamentos com maior disponibilidade de água, entretanto com o déficit hídrico (25%cc) a área foliar chegou a ser aproximadamente 70% menor que nas mudas com 70%cc (FIGUEIROA et al., 2004). A altura das mudas de Tabebuia aurea (Manso) Benth. & Hook f. ex S. Moore (ipê-amarelo) cultivadas com 100, 50 e 25%cc foi maior nas plantas submetidas a maior disponibilidade de água e a área foliar foi menor sob 25%cc (CABRAL et al., 2004). Entretanto Batista et al. (2008) trabalhando com Cecropia pachystarchya Trec em solo drenado e alagado não observaram diferenças no diâmetro em nenhum dos tratamentos. Figueiroa et al. (2004) observaram que o aumento da área foliar total é explicado pela boa disponibilidade hídrica e tal fenômeno é devido à relação existente entre o tamanho alcançado pelas folhas e a umidade do solo. Assim, em solo seco são produzidas folhas pequenas, o que contribui para reduzir o aquecimento dos tecidos foliares e a transpiração na estação mais quente. Sob essas condições, o equilíbrio entre a produção de assimilados e a demanda para o desenvolvimento dos órgãos reprodutivos é severamente afetada pela redução na área foliar fotossinteticamente ativa. Observaram também resultados semelhantes para espécies da caatinga como Anadenanthera macrocarpa (Benth.) Brenan, Parkinsonia aculeata L. e Acassia farnesiana (L.) Willd. Os valores médios da massa seca da parte aérea foram maiores com a maior disponibilidade de água, chegando a ser 50% menor sob menor disponibilidade de água na primeira avaliação com 25 e 12,5%cc em relação aos outros e do 25%cc em relação ao 100%cc na segunda avaliação (Tabela 2). A massa seca de raiz também foi maior com a maior disponibilidade de água não variando entre os níveis 100, 50 e 25%cc na primeira avaliação, mas sendo 50% menor a 12,5%cc em relação a 100%cc. Na segunda avaliação as mudas cultivadas com apenas 25%cc apresentaram acúmulo de massa seca radicular menor sob 25%cc. Observa-se que embora não tenha havido diferença significativa entre o comprimento médio das raízes com a redução da disponibilidade hídrica, estas apresentavam menor massa seca sugerindo que deveriam ser bem mais finas. Embora a adaptabilidade e tolerância ao estresse hídrico sejam estratégias intrínsecas de diferentes espécies vegetais, resultado semelhante foi encontrado por Figueroa et al. (2004) que também verificaram menor massa seca da raiz de mudas de Myracrodruon urundeuva sob déficit de água, e por Silva et al. (2002) para Melaleuca alternifolia Cheel quando a deficiência hídrica severa diminuiu o crescimento e a produção de biomassa verde e seca das plantas. Em mudas de Mimosa caesalpiniifolia Benth. cultivadas sob 100, 50 e 25% da capacidade de recipiente não foram observadas diferenças entre os tratamentos aos 25 dias, contudo aos 50 dias, em plantas sob restrição hídrica mais severa, a produção de massa seca foliar diminuiu (SANTIAGO et al., 2002). A relação raiz/parte aérea na primeira avaliação foi maior nas plantas cultivadas com 25%cc e na segunda com 50%cc. Provavelmente o nível de tolerância das mudas dessa espécie tenha sido atingido aos 35 dias no tratamento com 25%cc quando a planta investiu maior quantidade de fotoassimilados para o desenvolvimento radicular em detrimento da parte aérea, entretanto, aos 83 dias as mudas sob o mesmo nível de água, provavelmente já não conseguiam realizar atividade fotossintética e produzir fotoassimilados necessários para a manutenção do crescimento, sendo 25%cc um nível restritivo para o crescimento da planta. Essa maior alocação de fotoassimilados para as raízes na muda em condição de déficit hídrico sugere uma priorização do crescimento radicular, o que favorece a absorção de água e menor perda de água por transpiração (FIGUEROA et al., 2004; TAIZ e ZEIGER, 2004). A arquitetura radicular e sua capacidade de exploração das camadas mais profundas e úmidas do solo, juntamente com maior razão entre raiz e parte aérea, são características importantes de escape aos déficits hídricos. A continuidade do crescimento radicular, sob condições de menor disponibilidade hídrica, depende da manutenção de uma pressão de turgor mínima nas células, que seja suficiente para permitir o alongamento da parede celulósica e o crescimento celular. Quando o potencial da água é reduzido nas raízes, pode-se verificar em várias espécies um rápido ajuste osmótico, auxiliando o restabelecimento da pressão de turgor e permitindo a manutenção do alongamento celular. Em contraste, o ajuste osmótico nas folhas ocorre mais lentamente, levando à diminuição ou à parada da extensão das paredes celulares e ao menor crescimento da parte aérea (HSIAO e XU, 2000). O crescimento das mudas de mutambo foi maior com maior disponibilidade de água, sugerindo que a espécie não tolera deficiência hídrica. De acordo com Taiz e Zeiger (2004) a redução do turgor é o efeito mais precoce da deficiência de água na planta, e todos os processos que dependem diretamente dele, como a expansão foliar e alongamento de raízes, tornam-se prejudicados. A falta de água pode tornar as plantas vulneráveis a pragas e doenças, além de induzir o fechamento estomático, reduzir o crescimento, causar o acúmulo de solutos e antioxidantes, reduzir a área foliar e o crescimento do caule (SILVA et al., 2002). O estresse hídrico desencadeia a síntese de ácido abscísico nas raízes onde estimula também o crescimento das raízes e a formação de raízes laterais, e ao ser transportado para diferentes partes da planta, irão apresentar dentre outras respostas, a supressão do crescimento da folha e a sua abscisão, levando também à antecipação do processo reprodutivo (LARCHER, 2000; TAIZ e ZEIGER, 2004). Essa resposta de abscisão foliar foi observada nas mudas de mutambo cultivadas sob 12,5%cc já a partir de 35 dias de tratamento. A queda no conteúdo de água no solo aos 60 dias de tratamentos com maior restrição de água, agravada pela evapotranspiração, pode ter acarretado em fechamento parcial dos estômatos das mudas, levando também à redução da condutância estomática que implica, por sua vez, em queda da transpiração e da assimilação de CO2 (Tabela 3), visto que ambos são processos difusivos, mas que levam à redução na produção de fotoassimilados. Esses efeitos foram mais pronunciados nas mudas cultivadas sob 12,5%cc. Entretanto, a eficiência no uso da água não variou significativamente entre os tratamentos. O mutambo é uma espécie pioneira que apresenta melhor capacidade de dissipação térmica comparado às espécies tardias ao longo de um dia ensolarado, desde que não exista restrição hídrica. Souza e Ribeiro (2008) observaram que o valor máximo de fotossíntese (7 mol m-2 s-1) foi alcançado às 9h, e as maiores taxas transpiratórias às 11h (5 mmol m-2 s-1) horário de maior demanda atmosférica em ambiente com déficit de pressão de vapor de 3-5 KPa entre 9 e 11h. Quando bem irrigadas, os resultados demonstram uma capacidade dissipativa da incidência luminosa (fluxo de fótons fotossinteticamente ativos) significativamente superior ao longo do dia, em relação a uma espécie tardia da sucessão. Em condições de restrição hídrica o mutambo mostrou melhor eficiência de dissipação apenas nas primeiras horas do dia (entre 7 e 9h). O déficit de água normalmente leva a uma diminuição da taxa de fotossintética, embora os níveis de tolerância possam variar para diferentes espécies vegetais. Mudas de Myracrodruon urundeuva com um ano de idade e submetidas à suspensão da irrigação por 14 dias apresentaram redução da condutância estomática (QUEIROZ et al., 2002). De maneira semelhante, laranjeiras apresentaram reduções significativas da transpiração e condutância estomática (GOMES et al., 2004) e mudas de Schinopsis brasiliensis redução da transpiração (NOGUEIRA e SILVA, 2002) ambas após sete dias de suspensão da irrigação. Redução na assimilação do carbono tanto na espécie pioneira Bauhinia forficata quanto na secundária Esenbeckia leiocarpa após 45 dias sem chuva e avaliadas em condição de sub-bosque e de clareira também foi observada por Portes et al. (2006). Medrano et al., (2002) e Parry et al., (2002) comentam que a seca reduz o metabolismo do mesofilo ao diminuir a ativação e a atividade da Rubisco carboxilase. Além disso, a menor eficiência da Rubisco pode ser causada por uma resistência maior do mesofilo devido ao fechamento estomático, restringindo a absorção de CO2 nos cloroplastos e aumentando a ação oxigenase de Rubisco, e em consequência, a fotorrespiração. A Taxa assimilatória líquida (TAL), taxa de crescimento relativo (TCR) e razão de área foliar (RAF) foram maiores nas mudas com 100%cc (Tabela 4). Esses resultados podem ser devidos à maior área foliar e à maior abertura dos ostíolos (Tabela 1 e Tabela 4), que propiciaram uma maior taxa fotossintética (Tabela 3) logo uma maior produção de fotoassimilados, uma vez que folhas com maior abertura estomática podem absorver melhor o CO2 atmosférico e fotossintetizar melhor. CONCLUSÃO Observou-se que para a produção de mudas de Guzuma ulmifolia o regime hídrico ideal na fase inicial é de 100% da capacidade de campo, porém as mudas crescem satisfatoriamente sob 50%cc. Abaixo de 25%cc as mudas entram em estresse hídrico antes dos 35 dias de cultivo. AGRADECIMENTOS A FUNDECT pelo apoio financeiro e ao CNPq e CAPES pela concessão de bolsas. REFERENCIAS BIBLIOGRÁFICAS
Copyright © 2011 - Ciência Florestal The following images related to this document are available:Photo images[cf11064t1.jpg] [cf11064t3.jpg] [cf11064t2.jpg] [cf11064t4.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}