 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
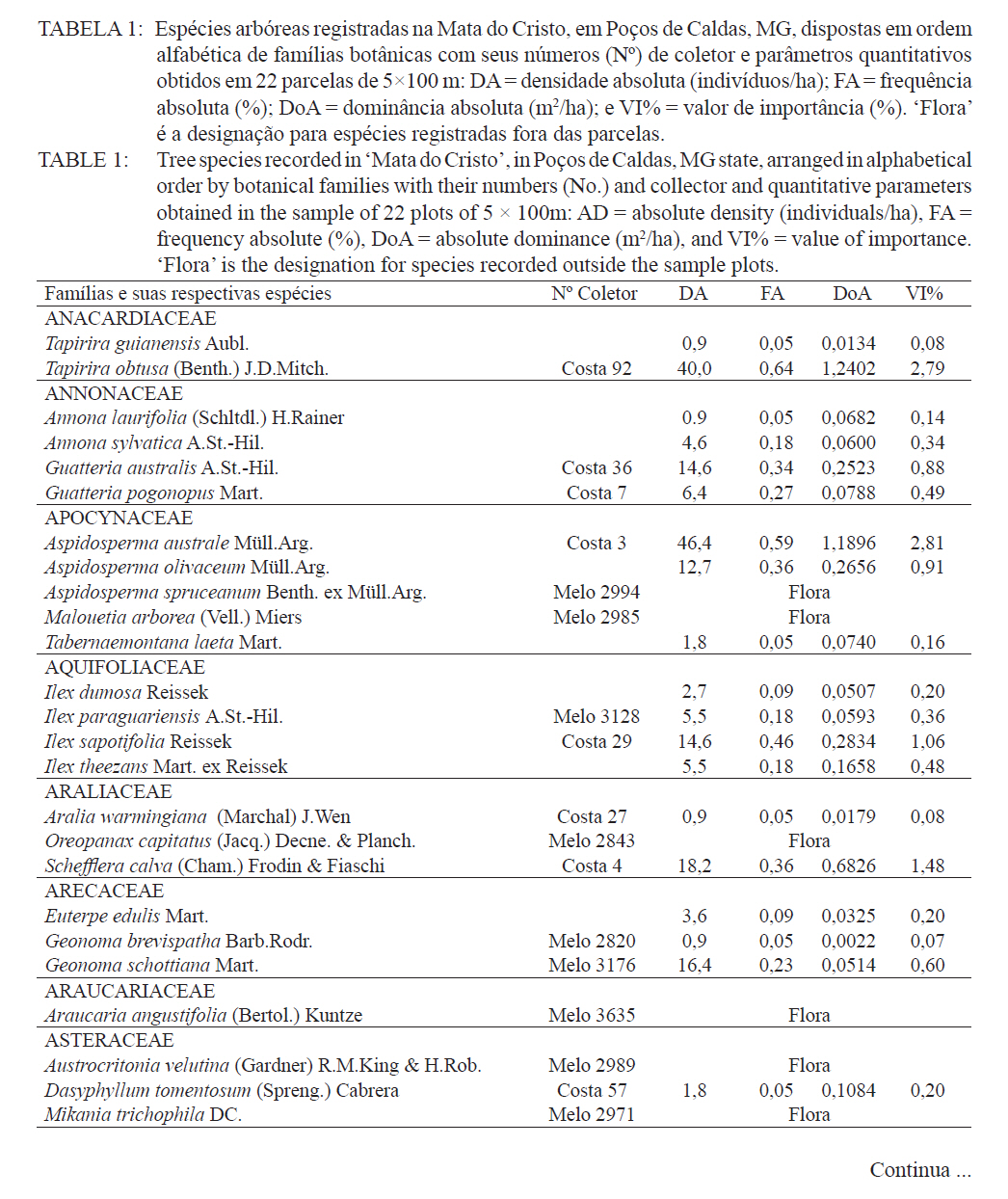
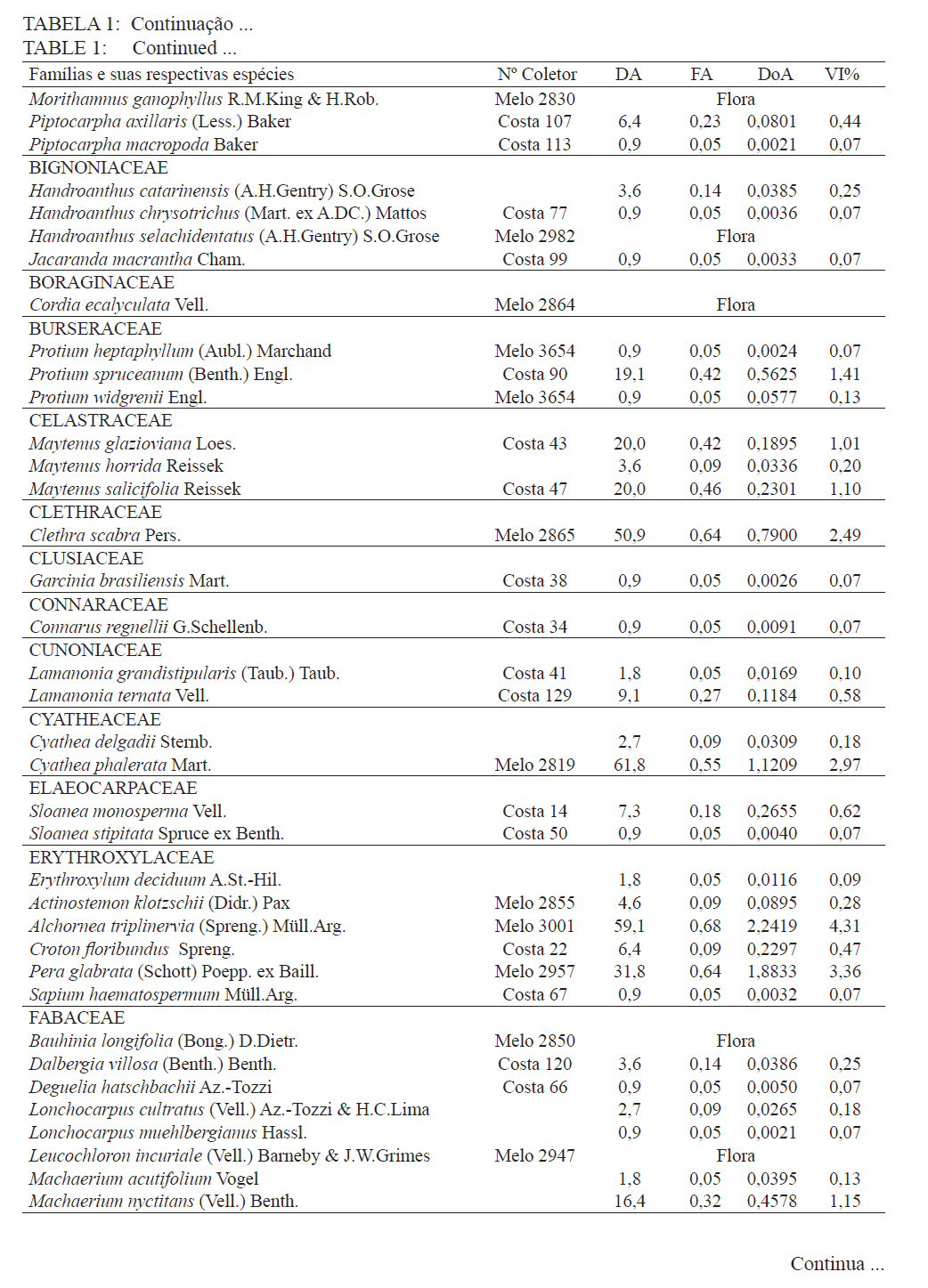
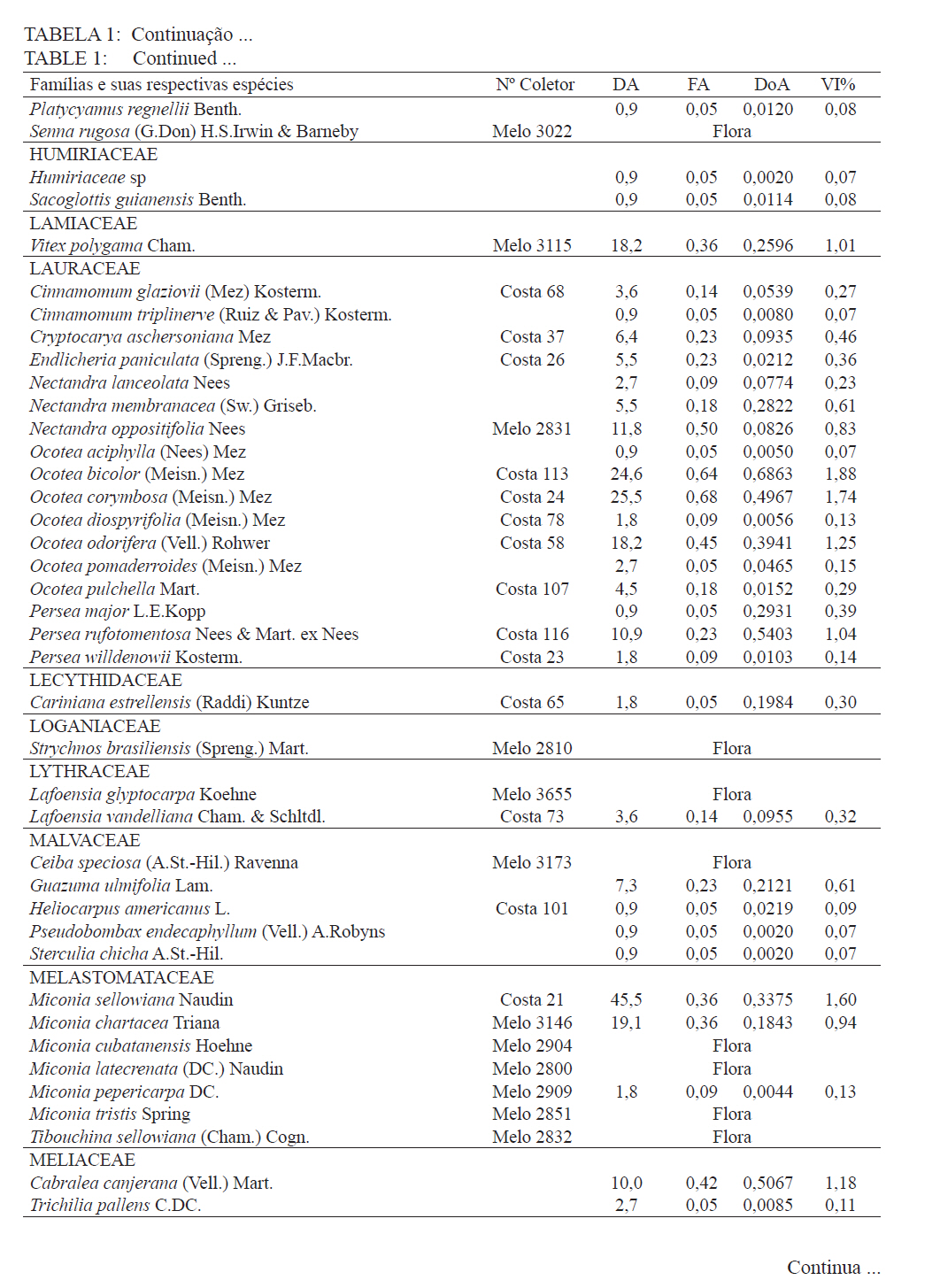
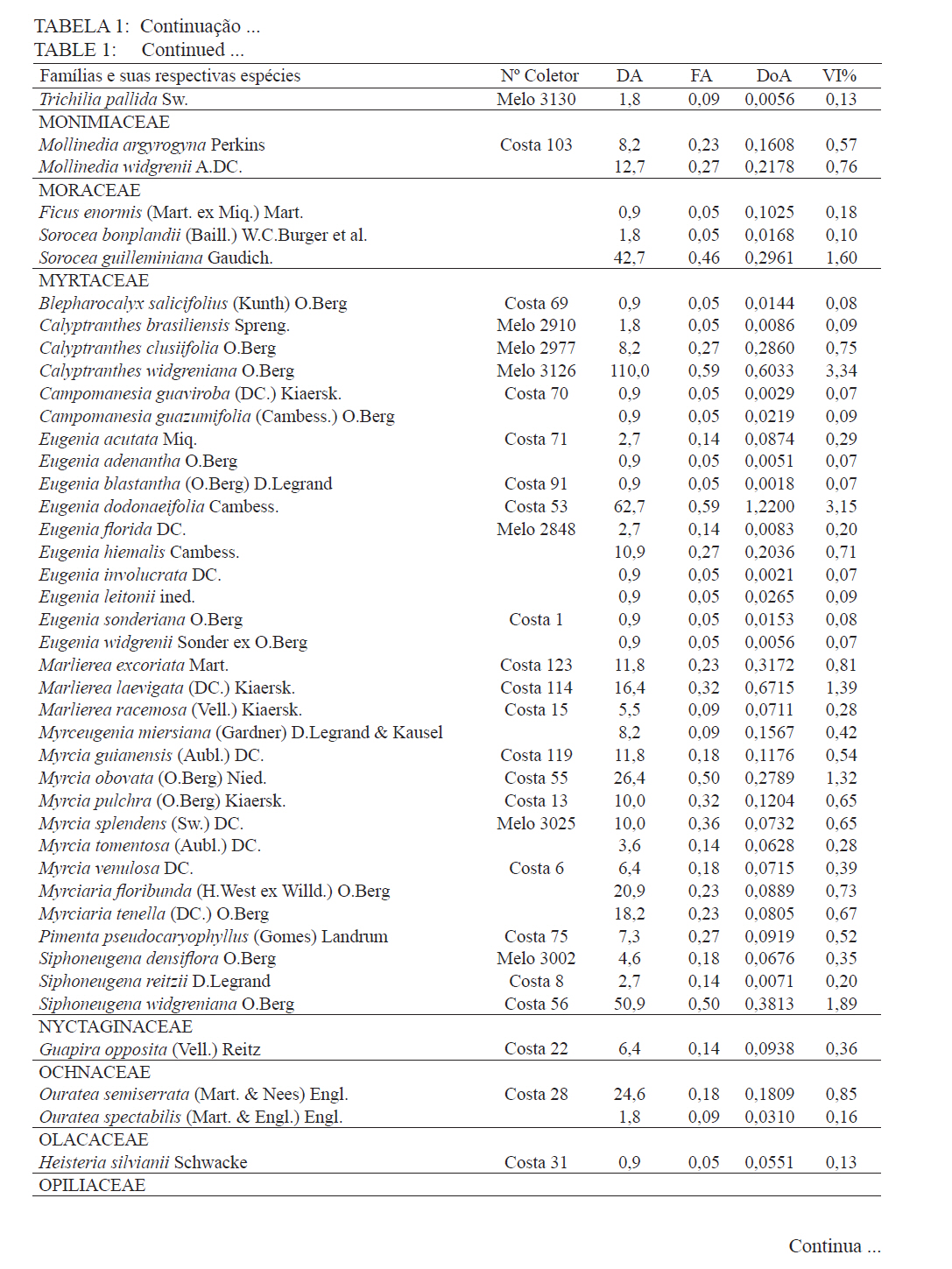
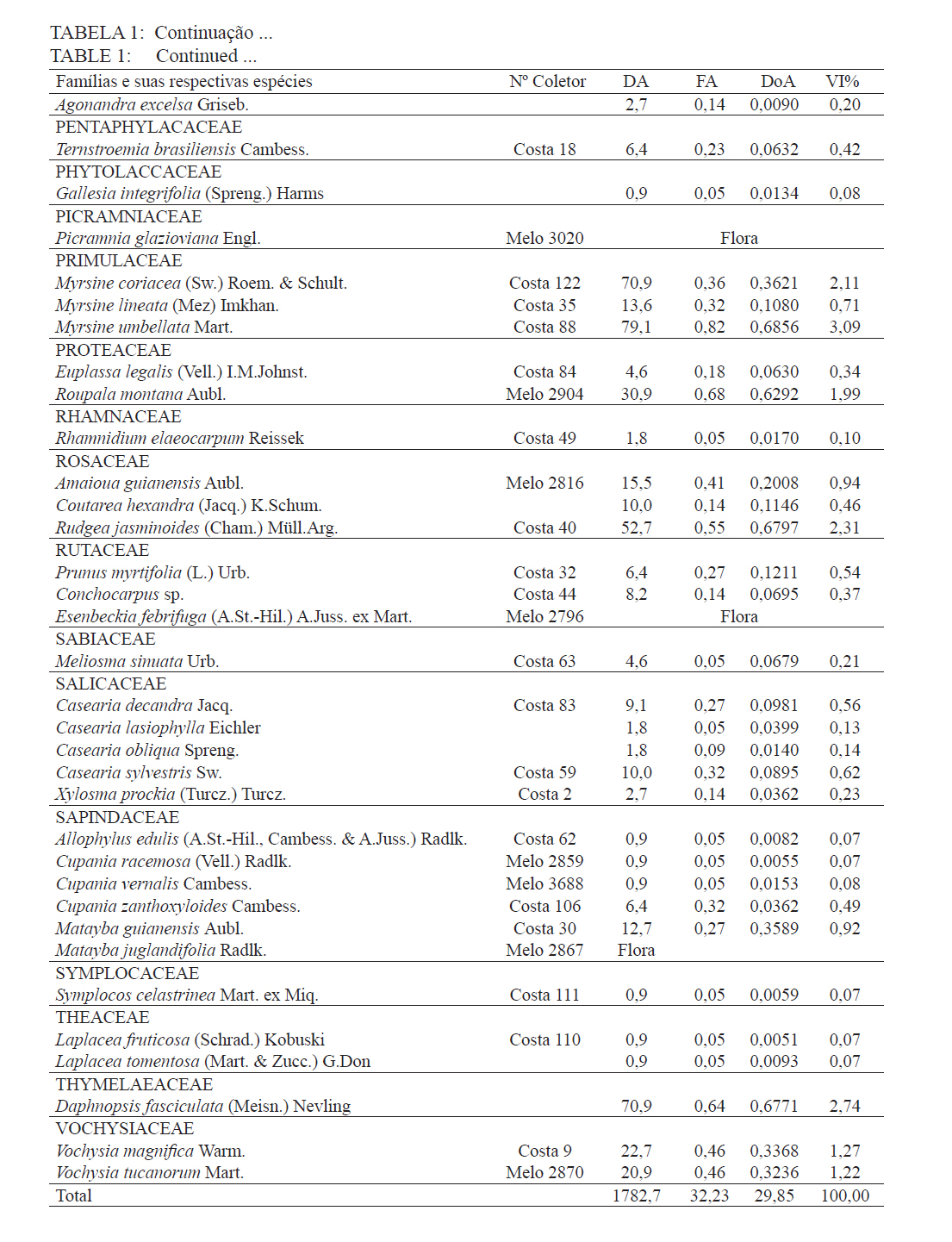
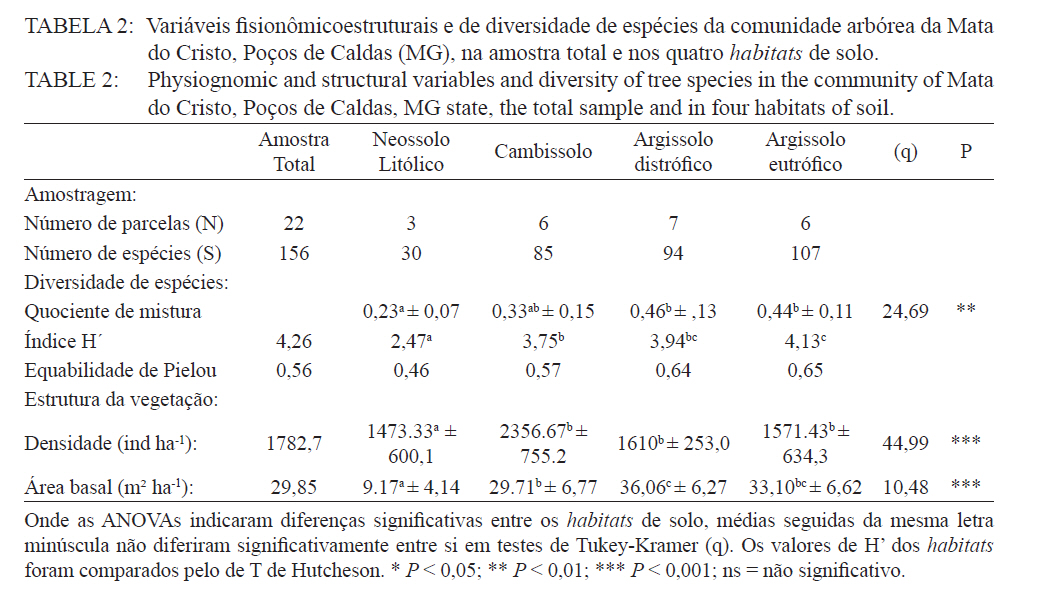
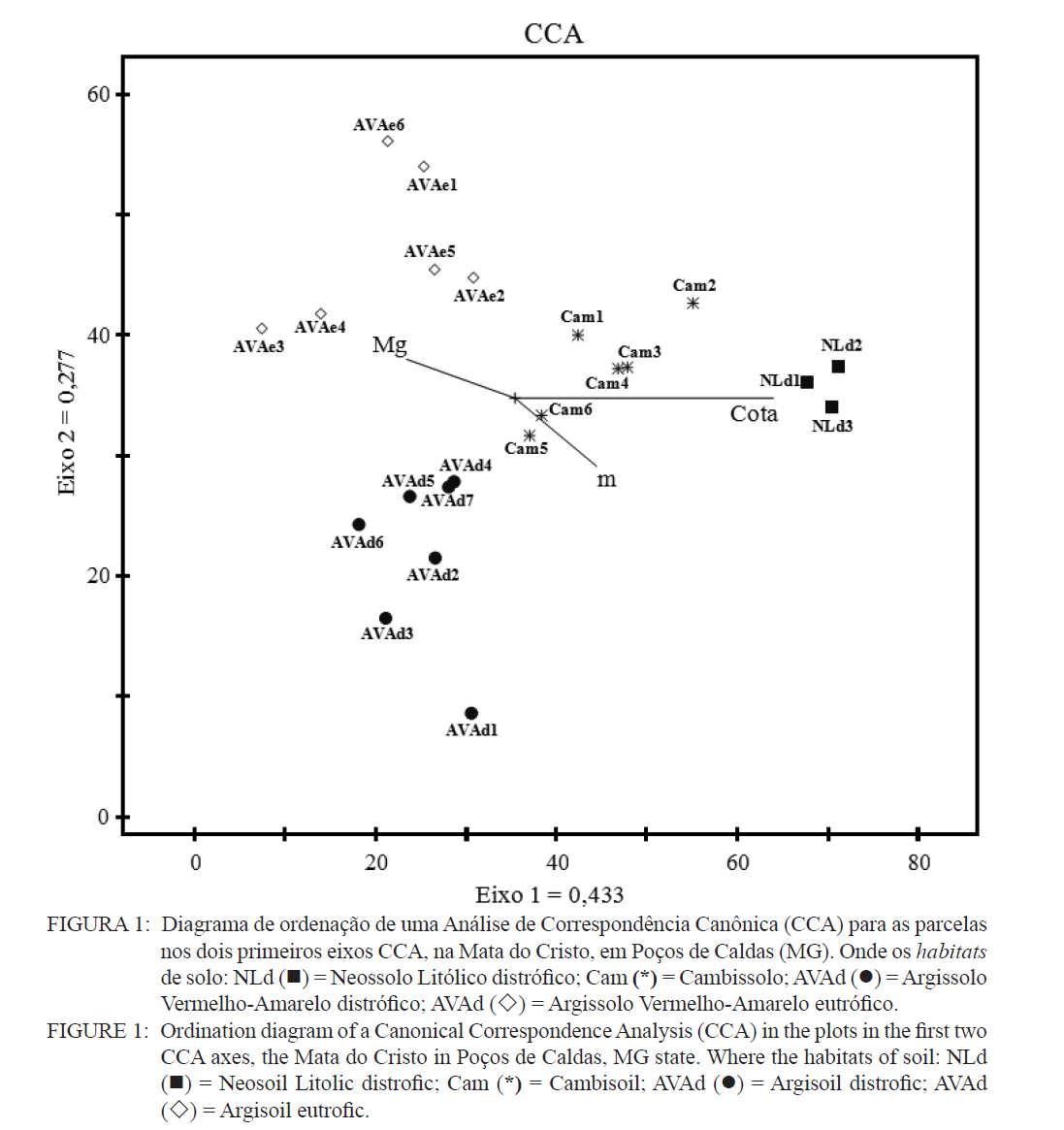
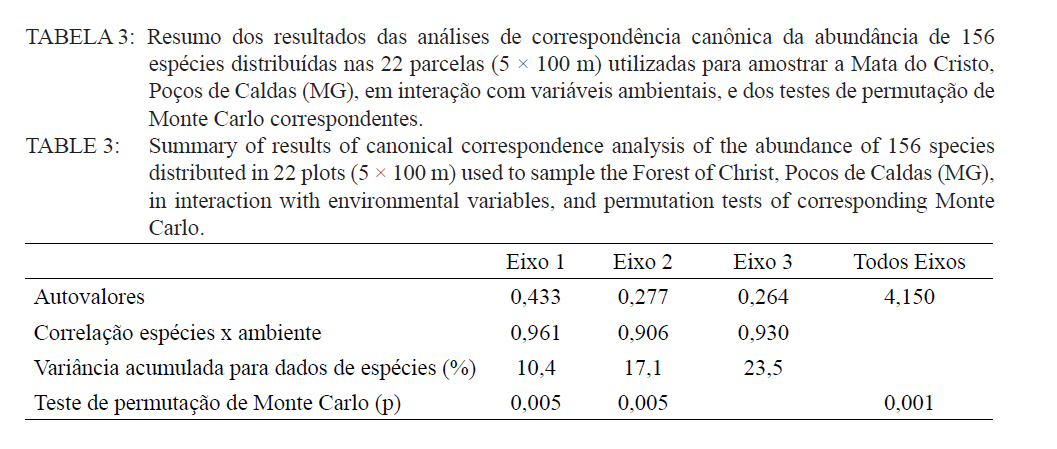
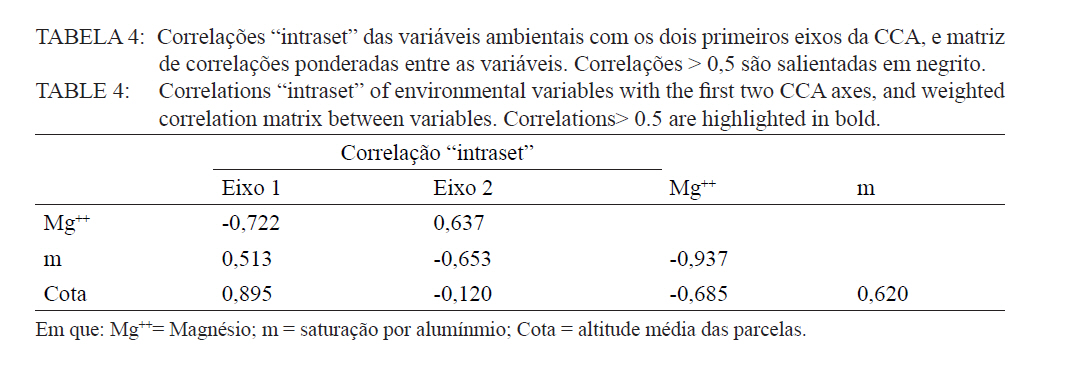
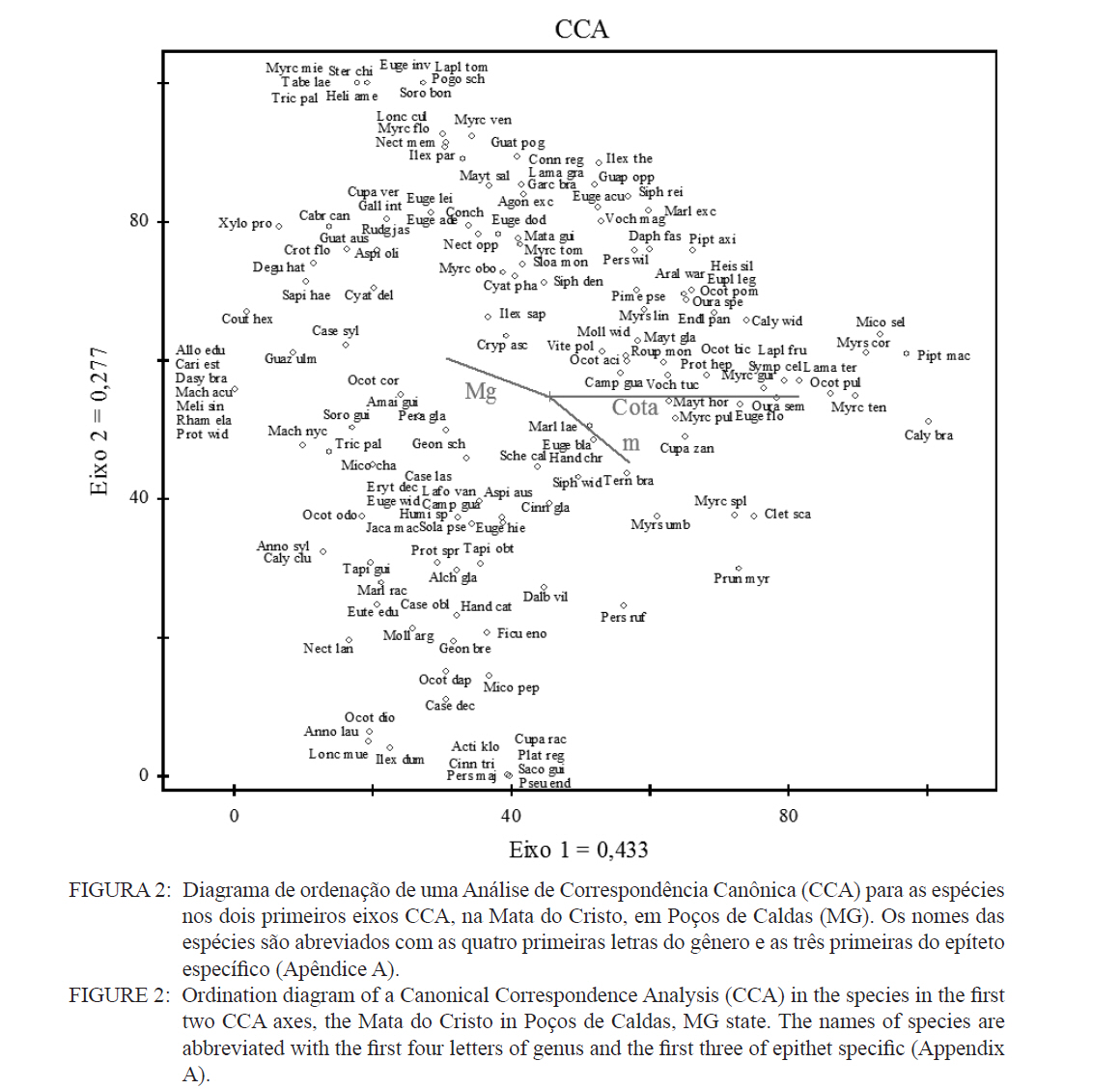
Ciência Florestal, Santa Maria, v. 21, n. 4, p. 711-725, out.-dez., 2011 ESTRUTURA E DIVERSIDADE DA COMUNIDADE ARBÓREA DE UMA FLORESTA SUPEROMONTANA, NO PLANALTO DE POÇOS DE CALDAS (MG) STRUCTURE AND DIVERSITY OF THE TREE COMUNITY OF A UPPER HIGHLANDS FOREST ON THE PLATEAU OF POÇOS DE CALDAS, MG STATE Malcon do Prado Costa1 José Aldo Alves Pereira2 Marco Aurélio Leite Fontes3 Pablo Hendrigo Alves de Melo4 Daniel Salgado Pífano5 André Schatz Pellicciottii6 Patrícia Vieira Pompeu7 Rossi Allan Silva8 1Engenheiro Florestal, MSc., Doutorando pelo Programa de Pós-graduação em Engenharia Florestal, Universidade Federal de Santa Maria, Av. Roraima, 1000, CEP 97105-900, Santa Maria (RS). malconfloresta@gmail.com Recebido para publicação em 2/04/2010 e aceito em 13/12/2010 Code Number: cf11069 RESUMO Foi realizado o levantamento da comunidade arbórea de uma floresta tropical superomontana, situada na Serra de São Domingos, no planalto de Poços de Caldas, Minas Gerais, Brasil (altitude de 1.200 m a 1.575 m; 21°46’25”S, 46’24”10”W), com o objetivo de avaliar as variações estruturais e variáveis ambientais relacionadas. Foram analisados aspectos da estrutura fisionômica (densidade, área basal e distribuição de tamanho das árvores) e comunitária (composição, distribuição e diversidade de espécies). Foram alocadas 22 parcelas de 5 x 100 m no fragmento para amostragem dos indivíduos arbóreos com diâmetro à altura do peito (DAP) ≥ 5 cm, e coleta de dados topográficos e amostras de solo superficial (0-20 cm) para análises químicas e texturais. Foram registradas 53 famílias botânicas, 104 gêneros, 178 espécies e 1963 indivíduos, assim como três subgrupos de solos: Neossolos Litólicos, Cambissolos e Argissolos (distróficos e eutróficos), distribuídos ao longo da variação altitudinal do fragmento. Uma análise de correspondência canônica revelou um gradiente de distribuição das espécies arbóreas principalmente correlacionada com as variações altitudinais, o teor de Magnésio (Mg++) e saturação por alumínio (m). Palavras-chave: floresta de altitude; variáveis ambientais; fitossociologia; florística. ABSTRACT A survey of the tree community of a ombrophilous upper montane forest, situated in Poços de Caldas, Brazil (1200-1575 m of altitude; 21°46’25”S, 46’24”10”W) was carried out in order to assess its environmental and structural variation. The physiognomic (density, basal area and distribution of size of the trees) and community structure (composition, distribution and diversity of species) were analyzed through 22 plots of 5 x 100 m, where the tree individuals with diameter at breast height ≥ 5 cm were sampled. The topographic variables were measured and the surface soil samples were collected to obtain their chemical and texture attributes. The survey resulted in 53 botanic families, 104 genera, 178 species and 1963 individuals, as well as four soil classes: distrofic and eutrofic Argisoil, Cambisoil and Litolic Neosoil, distributed sequentially from the hilltop to the base of the fragment. The canonic correspondence analyses showed a gradient of tree species distribution mainly correlated with variations of quotas, magnesium content (Mg++) and aluminum saturation (m). Keywords: mountain forest; species diversity; environmental variables; phytosociology; floristic. INTRODUÇÃO A Serra de São Domingos, situada no planalto de Poços de Caldas (MORAES e JIMÉNEZ-RUEDA, 2008) abriga formações florestais atlânticas de altitude em suas maiores altitudes, acima dos 1.500 m, sendo estratégicas para a conservação da biodiversidade e serviços ambientais, como fornecimento de água (DUDLEY e STOLTON, 2003). A variação da altitude é um dos principais gradientes ambientais indiretos, capaz de influenciar o crescimento vegetal, a florística e a fisionomia das formações vegetacionais (PENDRY e PROCTOR, 1996; SANCHEZ, 2001). As florestas denominadas superomontanas na região SE situam-se entre 16º e 24º de latitude S e ocorrem acima dos 1.000 m (OLIVEIRA-FILHO, 2009). Dentre as tipologias Florestais Superomontanas, há uma fitocenose peculiar, as Nanoflorestas latifoliadas (sensu OLIVEIRA-FILHO, 2009), que ocorrem em ambiente nebular com árvores latifoliadas formando dossel baixo (3 a 5 m), com plantas emergentes mais altas e espalhadas; e requer conhecimentos ecológicos básicos para sua preservação, conservação e restauração. A transição entre mata atlântica de encosta e mata nebular demanda estudos fitossociológicos, inclusive para uma definição mais fundada do caráter da mata nebular, isto é, se corresponde a uma formação própria, individualizável, ou se pode ser incluída na formação em contato. Para Falkenberg e Voltolini (1995) é prematura uma tentativa de definição, sendo necessários mais estudos em mata atlântica de altitude, de porte menor (10-15 m), em áreas íngremes, para saber se suas submatas são semelhantes ou não às matas nebulares. Oliveira-Filho e Fontes (2000) realizaram uma análise ampla do complexo geográfico e altitudinal da mata atlântica no sudeste do Brasil, mas Falkenberg (2003) não dispunham de amostras de gradientes inteiros. Essas vegetações podem formar contínuos sob certas condições e serem discretas em outras, sendo a maioria intermediária entre tais extremos (BEALS, 1969), conforme as características biofísicas locais (KITAYAMA, 1995). Dessa forma, avaliações de gradientes altitudinais em florestas de altitude são importantes para compreensão das causas de diferenciação (KITAYAMA, 1992). Em Floresta Superomontana da Serra da Mantiqueira, Meirelles et al. (2008) verificaram uma florística distinta entre diferentes altitudes, com táxons restritos às maiores elevações, distinguindo-se também a altura do dossel, densidade e diâmetro dos indivíduos. Entretanto, as modificações ocorreram de forma gradual. O presente estudo foi realizado em uma floresta nebular de altitude, com transição entre fisionomia de Floresta latifoliada nas encostas e Nanoflorestas latifoliadas acima. A hipótese de nulidade foi que existe gradiente altitudinal que influencia a abundância das espécies. O objetivo deste trabalho foi caracterizar a comunidade arbórea de uma Floresta Superomontana na Serra de São Domingos através das análises: das correlações entre a distribuição das espécies arbóreas e as variáveis de solo e de relevo; dos padrões de diversidade e estruturais da vegetação ao longo do gradiente; das mudanças florísticas e demográficas na variação de altitude. MATERIAL E MÉTODOS Caracterização do local de estudo A área, chamada de Mata do Cristo, localiza-se no Parque Municipal da Serra de São Domingos, em Poços de Caldas, Minas Gerais, Brasil, nas coordenadas 21º46’25”S e 46º34’10”W, com altitudes de 1.200 a 1.575 m e área de 271 ha. Trata-se de mosaico de Floresta Tropical Superomontana (OLIVEIRA-FILHO, 2009), com florestas latifoliadas ribeirinhas e de média encosta e Nanoflorestas nebulares nas maiores elevações, na transição com Campos de Altitude. A precipitação média anual é de 1.695,5 mm e a temperatura média de 17,7ºC (ANA, 2009), com clima Cwb, na classificação Köppen, ou subtropical chuvoso, com frequentes chuvas orográficas. A Serra insere-se no Planalto de Poços de Caldas, na borda ocidental da Serra da Mantiqueira, entre o planalto alto (1.200 a 1.400 m) e muito alto (acima de 1.400 m), constituída de cristas e serras que podem atingir 1.700 m (MORAES e JIMÉNES-RUEDA, 2008). Os solos, pobres (com baixos teores de bases trocáveis e saturação de bases) ou de fertilidade média próximos aos riachos, na Mata do Cristo classificam-se como: Argissolo Vermelho-Amarelo (AVA cascalhentos ou, nas partes baixas e mais férteis, AVA eutróficos), Cambissolo (em associação com o primeiro) e Neossolo Litólico. Amostragem da vegetação Alocaram-se 22 parcelas de 5 x 100 m (total de 1,1 ha), distribuídas segundo o gradiente de relevo (Mapa, Apêndice B), com alinhamentos preferenciais L-W para captar menor variabilidade do relevo dentro de cada parcela e modificados em locais mais acidentados. As altitudes foram levantadas com GPS (Garmim CSx) para avaliação de seus efeitos na estrutura da comunidade, sendo o desnível mensurado pelas diferenças entre a maior e menor cota por parcela. Registraram-se todos os indivíduos arbóreos e arbustivos vivos com circunferência à altura do peito (CAP; 1,3 m) ≥ 15,7 cm, equivalente a um diâmetro de 5 cm (ou diâmetro médio quadrático equivalente, quando em indivíduos ramificados). Foram coletadas amostras de material botânico dos indivíduos registrados e realizado caminhamento expedito para coleta fora das parcelas. Herborização e identificação do material botânico As espécies encontradas férteis foram coletadas e as demais identificadas em campo. A herborização e a incorporação do material foram realizadas no Herbário ESAL da UFLA e a identificação com utilização da coleção botânica e por meio de consultas à literatura e especialistas. Os nomes foram atualizados por consulta ao índice de espécies do banco de dados do Missouri Botânical Garden (MOBOT, 2010). As identificações de angiospermas (exceto Cyatheaceae) seguem o sistema APG III (2009). Amostragem do solo Coletou-se uma amostra composta de solo superficial (0-20 cm) a partir de cinco amostras simples aleatórias por parcela. As amostras foram analisadas no Laboratório de Análise de Solos UFLA, segundo o protocolo da Embrapa (1997), obtendo-se as variáveis: pH; teores de P, K+, Ca++, Mg++ e Al+++; soma de bases (SB); saturação por bases (V) e por alumínio (m); capacidade de troca de cátions efetiva (t), capacidade de troca de cátions (CTC) a pH 7,0 (T), H+Al, teor de matéria orgânica (MO), fósforo remanescente (P-rem) e proporções de matéria orgânica, areia, silte e argila. Análise de dados Analisou-se a estrutura da floresta a partir dos parâmetros quantitativos por espécie: densidade, frequência e dominância absolutas e valor de importância (MUELLER-DOMBOIS e ELLEMBERG, 1974). A diversidade e riqueza de espécies, total e por habitat segundo a classe de solo (conforme EMBRAPA, 2006), foram avaliadas através dos índices de diversidade de Shannon (H’) e de equabilidade de Pielou (BROWER et al., 1997), sendo os valores H’ comparados pelo teste T de Hutcheson (ZAR, 1996), e da densidade de espécies, pelo quociente de mistura de Jentsch, que é a razão entre número de espécies e de indivíduos e informa sobre mudança na composição florística no gradiente (LONGHI, 1980). Nas comparações entre os habitats foram utilizadas análises de variância seguidas de testes de Tukey-Kramer, adotando-se um delineamento em blocos casualizados com números de parcelas diferentes. Para analisar correlações entre gradientes ambientais e vegetação, empregou-se uma análise de correspondência canônica (CCA) (TER BRAAK, 1987) utilizando o programa PC-ORD 5 (McCUNE et al., 2006), onde a matriz de abundância das espécies foi constituída do número de indivíduos por parcela, transformados por log10(a+1) para compensar desvios causados por valores extremos (TER BRAAK, 1995); e a matriz de variáveis ambientais constituída das duas variáveis topográficas (cota altimétrica e desnível) e as 17 variáveis dos solos, o que após uma CCA preliminar reduziu-se pela eliminação das 16 variáveis ambientais não correlacionadas ou redundantes com outras variáveis, sendo a CCA final processada com aquelas correlacionadas com os eixos de ordenação: cota média, magnésio e saturação por alumínio. RESULTADOS E DISCUSSÃO Estrutura fisionômica e diversidade de espécies da comunidade arbórea Amostraram-se 1.963 indivíduos, distribuídos em 156 espécies, 88 gêneros e 46 famílias (somando-se o levantamento expedito foram 178 espécies, 104 gêneros 53 famílias botânicas). As espécies mais importantes foram Alchornea triplinervia, Pera glabrata, Calyptranthes widgreniana, Eugenia dodonaefolia, Myrsine umbellata, Cyathea pharelata e Aspidosperma australe (Tabela 1a Tabela 1b Tabela 1c Tabela 1d Tabela 1e). Nos fundos de vales houve dominância de Cyathea phalerata e em menor escala Cyathea delgadii, sendo o sub-bosque ralo e composto por uma mescla de espécies, as próprias de sub-bosque e as recrutas de espécies de dossel como Cupania vernalis, Matayba guianensis, Euplassa legalis e Handroanthus catarinae; no subdossel destacaram-se as Mirtáceas como Myrceugenia miersiana, Calyptranthes clusaefolia, Calyptranthes widgreniana e Eugenia dodonaefolia, e as Lauráceas Ocotea bicolor e Nectandra membranacea. A Floresta Latifoliada Montana correspondeu à maioria das biocenoses encontradas, com dossel estruturado e densidade reduzida, ocorrendo espécies de maior área basal como Persea rufotomentosa, Schefflera calva, Eugenia dodonaefolia e Cabralea canjerana; no sub-bosque destacaram-se Geonoma schottiana e Geonoma brevispatha, enquanto o dossel e o subdossel partilharam espécies nos estratos verticais como Machaerium nictitans, Myrsine umbellata, Myrsine coriacea, Clethra scabra e Daphnopsis fasciculata. Nas Nanoflorestas Latifoliadas Nebulares não há estratificação vertical e as árvores são pequenas e adensadas formando um falso dossel que não ultrapassa 5 m de altura, onde ocorrem algumas espécies comuns às outras fisionomias como Myrsine coriacea, Eugenia dodonaefolia, Clethra scabra e Miconia urophylla, o que se explica pela mudança gradativa de nanoflorestas para matas de encostas, conforme observado em campo. O remanescente estudado constitui vestígio da vegetação arbórea original que, segundo Azevedo (1962), cobria os altos dos morros pertencentes ao complexo da Serra da Mantiqueira, nos estados de São Paulo, Minas Gerais e Rio de Janeiro, acompanhando as variações climáticas e de altitude. Devido à ocupação humana nessas regiões, a extensão florestal foi reduzida a vários remanescentes nas áreas mais elevadas e de difícil exploração, mas que guardam características da floresta pretérita (FRANÇA e STEHMAN, 2004). A família Myrtaceae, que aparece como mais rica em espécies, também o foi nas florestas de altitude de Camanducaia (FRANÇA e STEHMANN, 2004), Bocaina de Minas (CARVALHO, et al. 2005) e Monte Verde (MEIRELES et al., 2008). As famílias Lauraceae, Melastomataceae e Asteraceae, importantes aqui, são também nas áreas citadas (OLIVEIRA-FILHO e FONTES, 2000). Fabaceae, de baixa expressão em Camanducaia, a 1.900 m de altitude (FRANÇA e STEHMANN, 2004), com apenas uma espécie, neste trabalho ocorreu de forma expressiva, assim como em Bocaina de Minas, a 1.285 m (CARVALHO et al., 2005), confirmando Gentry (1988), que afirma existir uma redução drástica na família acima dos 1.500 m. Ademais, na Mata do Cristo foram encontradas 19 espécies ameaçadas em diferentes status (IUCN, 2009). As de baixo risco de extinção foram: Ilex paraguariensis, Lafoensia vandelliana, Mollinedia argyrogyna, Myrceugenia miersiana, Ocotea aciphylla e Trichilia pallens. Já as espécies Ilex theezans, Pimenta pseudocaryophyllus, Siphoneugena densiflora, Siphoneugena widgreniana, Sorocea guilleminiana são vulneráveis. Em perigo foram encontradas Casearia lasiophylla, Gloxinia sylvatica, Handroanthus catarinensis, Ocotea odorifera, Ocotea pomaderroides e Ocotea pulchella. E criticamente em perigo as espécies Euterpe edulis e Persea rufotomentosa. Das 156 espécies amostradas, 107 ocorreram no Argissolo Vermelho-Amarelo eutrófico, 94 no Argissolo distrófico, 85 no Cambissolo e 30 no Neossolo Litólico (Tabela 2). Considerando o quociente de mistura de Jentsch, os Argissolos apresentam maior riqueza que os Neossolos Litólicos e os Cambissolos foram indiferentes entre habitats. A diversidade expressa por H’ decresceu no sentido Argissolo eutrófico para Neossolo. A equabilidade de Pielou seguiu o mesmo padrão, decrescendo do Argissolo eutrófico para distrófico, Cambissolo e Neossolo Litólico, sugerindo dominância ecológica crescente, ou seja, concentração de abundâncias em poucas espécies no Neossolo Litólico. A “Teoria da Fertilidade Intermediária dos Solos” (TILMAN, 1982) afirma que ambientes com disponibilidade nutricional intermediária apresentam maior diversidade que locais com baixo ou elevado status nutricional, o que não acontece na área estudada. Já a “Hipótese de Distúrbios Intermediários” (CONNELL, 1978) preconiza que as florestas tropicais encontram-se submetidas a frequentes distúrbios a partir dos quais o equilíbrio nunca é alcançado, ocorrendo um máximo de diversidade em locais submetidos a distúrbios intermediários em intensidade, frequência e duração, acrescida a possibilidade de que os estágios sucessionais ocorram simultaneamente em uma dada área (ROBERTS e GILLIAM, 1995), o que parece se ajustar aos valores de diversidade encontrados, com variações entre os habitats, mas diversidade decrescente com a altitude, onde o regime de distúrbios é mais frequente. Além disso, na Mata do Cristo, com aumento da altitude há adensamento de árvores finas e pequenas nos habitats de Neossolo e Cambissolo, o que pode ser relacionado a particularidades desses habitats, por exemplo, pela baixa disponibilidade de água devido à elevação topográfica. Os Neossolos apresentaram baixos valores de densidade, pois as parcelas eram de nanoflorestas em transição com campos de altitude, com trechos sem indivíduos dentro do critério de inclusão. Correlações entre espécies e variáveis ambientais A CCA separou com clareza as parcelas de acordo com seus habitats de solo, concentrando os Neossolos na porção direita, os Cambissolos na parte central e os Argissolos à esquerda, além dos últimos se separarem no eixo 2, com os distróficos na parte inferior e os eutróficos na parte superior (Figura 1). Os autovalores para os três eixos foram baixos e a variância total explicada de apenas 23,5% (Tabela 3), indicando que as variáveis florísticas utilizadas contêm muito ruído, ou seja, alta proporção de variância não explicada, o que é comum em dados de vegetação (TER BRAAK, 1987), devido à estocasticidade comum em ambientes florestais (HUBBEL, 2001), pois a competição, luminosidade, umidade e efeitos de fatores bióticos como síndromes de dispersão e polinização não são facilmente perceptíveis. No entanto, isso não compromete os resultados, já que as correlações entre a abundância das espécies e as variáveis ambientais foram elevadas (eixo 1 = 0,961; eixo 2 = 0,906) e significativas para os dois primeiros eixos e para o somatório de todos os eixos, pelo teste de permutação de Monte Carlo (Tabela 3). Os autovalores baixos (<0,5) indicam gradientes curtos, ou seja, pouca substituição de espécies e a maioria delas se distribuindo ao longo dos gradientes, variando sua abundância (TER BRAAK, 1995). Na Tabela 4, apresentam-se as correlações entre os dois primeiros eixos da CCA e as variáveis ambientais, e destas entre si. O primeiro eixo foi positivamente correlacionado com a cota e saturação por Al e negativamente o Mg. O segundo eixo foi positivamente correlacionado com Mg e negativamente com a saturação por Al. Nas correlações elevadas entre as variáveis (>0,5), o Mg foi negativamente correlacionado com a saturação por Al e a cota, enquanto as mesmas foram correlacionadas entre si positivamente. Assim, foram claras as correlações entre características da comunidade arbórea e as variáveis ambientais relacionadas ao substrato. É comum que a fertilidade química dos solos de uma catena cresça do topo para base da encosta (KITAYAMA e AIBA, 2002; OLIVEIRA, 2005; RESENDE et al. 2007). De uma maneira geral, a radiação solar e a disponibilidade de água e de nutrientes minerais são os fatores preponderantes na determinação dos padrões de distribuição de plantas ao redor do mundo (HUGGET, 1995). Na Mata do Cristo, as variações da radiação solar foram avaliadas unicamente pelo efeito das cotas altimétricas, ao passo que água e nutrientes foram avaliados pelas propriedades do substrato. Além disso, a altitude foi um fator com relação expressiva com as mudanças na estrutura da comunidade, corroborando com os resultados encontrados por Meireles et al. (2008), em uma floresta superomontana do município de Monte Verde - MG, inserido na Serra da Mantiqueira. As variações ambientais da Mata do Cristo foram correspondidas por distribuições diferenciais da abundância das espécies, com consequências para os padrões de diversidade. A heterogeneidade ambiental em florestas tropicais não controla diretamente a distribuição do número de espécies por meio da variação espacial da disponibilidade de recursos, mas age sobre a exclusão competitiva, o que permite a distribuição diferencial de algumas espécies (STEVENS e CARSON, 2002). Desta forma, a quantidade de habitats tem influência nas abundâncias relativas de algumas espécies, embora isso não seja estático no tempo, pois varia com as fases de regeneração da floresta, além de sofrer a influência de eventos estocásticos (DECOCQ, 2006). A ordenação das espécies pela CCA (Figura 2) sugere que espécies como Ocotea pulchela, Myrciaria tenella, Miconia sellowiana, Calyptranthes brasiliensis, Ocotea pulchela e Lamanonia ternata são abundantes nas áreas de maior altitude, nas Nanoflorestas, onde existe predominância de Cambissolos e Neossolos. No outro extremo do gradiente, correspondente aos sítios mais úmidos, adjacentes ao rio (Florestas Latifoliadas ribeirinhas) e com predominância dos Argissolos eutróficos, concentram-se espécies como Allophylus edulis, Coutarea hexandra, Nectandra lanceolata, Annona sylvatica, Xylosma prockia e Croton floribundus. Em condições intermediárias do gradiente (Florestas de encosta), ou indiferentes ao mesmo, relacionam-se espécies como Cryptocarya aschersoniana, Vitex polygama, Marlierea laevigata, Eugenia blastantha, Schefflera calva e Ilex sapotifolia. Os fatores ambientais bióticos e abióticos influenciam a dinâmica das populações vegetais no tempo e no espaço (SCUDELLER et al., 2001). A grande proporção de variação não explicada reforça a ideia que padrões estocásticos podem preponderar sobre os ambientais não como um agente causal, mas casual, na distribuição de espécies e na diversidade alfa, implicando no fato de a maioria delas serem, de fato, generalistas de habitats (HUBBELL, 2001). A Mata do Cristo apresentou correlação entre a distribuição de espécies e as características do substrato, com ênfase nas cotas altitudinais e na disponibilidade de Mg e acidez nos solos. Apesar disso, também foi identificado um razoável montante de variância não explicada pelas variáveis ambientais utilizadas (‘ruído’), e isso se deve, provavelmente, à estocasticidade dos fenômenos de estabelecimento e crescimento, embora não possamos descartar a possível existência de variáveis importantes, mas nem sempre facilmente perceptíveis ou mensuráreis, como condições de luz atuais e pretéritas e variáveis associadas à dispersão das espécies, além da competição por recursos. CONCLUSÕES A estrutura e diversidade da comunidade arbórea da Floresta Superomontana estudada no planalto de Poços de Caldas, MG, apresentou pouca variação na composição de espécies entre suas diferentes fisionomias, havendo mais variação na abundância dessas espécies; mostrou uma estrutura correlacionada com altitude, teor de Mg e saturação por Al, por sua vez ligados ao gradiente ao longo da encosta, mas também forte conteúdo estocástico; exibiu um padrão de decréscimo da diversidade com a elevação, assim como aumento de abundância e redução na área basal. Assim, a transição entre floresta e nanofloresta apresentou pouca distinção florística, diferenciando-se demográfica e fisionomicamente, compondo uma vegetação com características peculiares para a região e de grande importância para a conservação da biodiversidade. REFERÊNCIAS BIBLIOGRÁFICAS
Copyright © 2011 - Ciência Florestal The following images related to this document are available:Photo images[cf11069t4.jpg] [cf11069t1a.jpg] [cf11069f1.jpg] [cf11069t1d.jpg] [cf11069t1e.jpg] [cf11069t1b.jpg] [cf11069t1c.jpg] [cf11069t3.jpg] [cf11069f2.jpg] [cf11069t2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}