 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Chilean Journal of Agricultural Research (formerly Agricultura Técnica), Vol. 68, No. 2, Apr-Jun, 2008, pp. 119-127 Research Effect of Nitrogen Source on Some Rhizospheric Properties and Persistence of Mycorrhizal Fungal Propagules in an Andisol Efecto de la fuente de nitrógeno sobre algunas propiedades rizosféricas y persistencia de propágulos de hongos micorrícicos en un Andisol Pablo Cornejo[1]*, Rosa Rubio1 and Fernando Borie1 [1] Universidad de La Frontera, Departamento de Ciencias Químicas, Casilla 54-D, Temuco, Chile. E-mail: pcornejo@ufro.cl* Corresponding author. Received: 20 December 2007. Code Number: cj08011 Abstract Nitrogen fertilization influences plant growth and rhizospheric properties, affecting the functionality and persistence of arbuscular mycorrhizal fungi (AMF). In order to analyze the effect of two N-sources (NH4+ and NO3-) on the persistence of AMF propagules (colonized root, mycelium and spores) and some rhizospheric parameters (pH, P available), a greenhouse experiment was carried out using two wheat (Triticum aestivum L.) cvs. Otto and Metrenco, which were grown in an Andisol. Plant biomass was determined at the dry grain stage (Zadocks 99, 150 days after sowing, DAS), the density of AMF propagules was determined three months later (240 DAS), and pH and available P were determined at both stages. The results showed that N-source and cultivar influenced most of the studied variables. The NO3- + Metrenco combination showed the highest values for biomass, pH, available P and AMF spores (150, 13, 5 and 375% more than NH4+ + Otto interaction, respectively; p < 0.001). On the other hand, close relationships were found between biomass production and density of AMF spores (r2 = 0.89; p < 0.001), suggesting that this propagule is quantitatively affected by host plant biomass production. This propagule is probably also indirectly affected by rhizospheric conditions. This result is of a special agronomic and ecological interest in acidic soils destined to annual crops, in which nitrogen fertilization is a habitual practice, being the N-source very influential in yields. Key words: acidic soils, available P, wheat cultivars, nitrogen fertilization, rhizospheric pH, Triticum aestivum. RESUMEN La fertilización nitrogenada influye en el crecimiento vegetal y propiedades rizosféricas, afectando la funcionalidad y persistencia de hongos micorrícico-arbusculares (AMF). Para analizar el efecto de dos fuentes de N (NH4+ y NO3-) sobre la persistencia de propágulos de AMF (raíz colonizada, esporas e hifas) y algunos parámetros rizosféricos (pH, P disponible), se realizó un experimento en invernadero utilizando dos cultivares de trigo (Triticum aestivum L.), cvs. Otto y Metrenco en un Andisol. La biomasa se determinó en grano seco (Zadocks 99, 150 días después de la siembra, DAS), la densidad de propágulos de AMF se determinó tres meses más tarde (240 DAS), y el pH y P disponible se determinaron en ambas etapas. Los resultados indicaron que la fuente de N y el cultivar influyeron en la mayoría de las variables estudiadas. La combinación NO3- + Metrenco presentó mayores valores de biomasa, pH, P disponible y esporas de AMF (150; 13; 5 y 375% más que la interacción NH4+ + Otto, respectivamente; p < 0,001). Por otra parte, se encontraron estrechas relaciones entre producción de biomasa y densidad de esporas de AMF (r2 = 0,89; p < 0.001), que sugiere que este propágulo es cuantitativamente afectado por la producción de biomasa de la planta hospedera. Este propágulo es probablemente afectado de forma indirecta por las condiciones rizosféricas. Este resultado es de especial interés ecológico y agronómico en suelos ácidos destinados a cultivos anuales, donde la fertilización nitrogenada es una práctica habitual, siendo la fuente de N muy incidente en los rendimientos. Palabras clave: suelos ácidos, P disponible, cultivares de trigo, fertilización nitrogenada, pH rizosférico Triticum aestivum. INTRODUCTION Nitrogen fertilization is a habitual practice used by farmers in intensive agriculture systems, as occurs in volcanic soils (Ultisols and Andisols) of southern Chile, which cover a surface area of about 5 x 104 km2, dedicated mainly to cereal-legume rotation (Besoaín, 1985). Frequently, these types of soils present fertility limitations due to low P availability associated with high levels of exchangeable Al and low pH values (Baligar and Fageria, 1997). Thus, the application of different N-sources (in particular NO3- and NH4+) is of special relevance, since they represent an important fraction of cations or anions absorbed and/or accumulated by plants, affecting directly soil pH due to the physiological extrusion of H+ or OH- by the roots (Stange et al., 1995; Gerendás et al., 1997). Particularly, NH4+ also strongly influences the reduction of soil pH through the generation of H+ produced by the nitrification process (Campillo and Rodríguez, 1984; Martens, 2001). These rhizospheric changes will directly affect the solubility and mobility of several nutrients (Siqueira and Moreira, 1997) and the activity of diverse groups of microorganisms, including both symbiotic and free-living species (Jeffries and Barea, 2001). Among the soil microbial groups that play a key role in the different rhizospheric processes, the arbuscular mycorrhizal fungi (AMF) (phylum Glomeromycota) are able to establish symbiotic associations known as arbuscular mycorrhizas (AM) with the roots of most terrestrial vascular plants (Smith and Read, 1997; Barea, 1998; Schüssler et al., 2001). The main function of AM is nutritional, since the fungal hyphae act as a complementary root system, increasing the absorption area and allowing acquisition of nutrients with low mobility in the soil, or that are situated far from the root (Baligar and Fageria, 1997; Bago et al., 2000; Clark and Zeto, 2000). These characteristics of mycorrhizal symbiosis present a special relevance in acidic soils, since it favors the root absorption of typically limited elements in this type of soil, particularly P, N and several microelements (Clark et al., 1999; Clark and Zeto, 2000; Borie et al., 2002). Different agronomic practices can affect AM functionality, as well as the density of spores, mycelia and colonized roots present in the soil. Among others, the crop system, the rotation design and the amount and type of fertilizer used are of particular relevance (Mendoza and Borie, 1998; Borie and Rubio, 1999; Jeffries and Barea, 2001). In contrast, it had been demonstrated that extraradical AMF hyphae can uptake NH4+ as NO3- from the soil (Johansen, 1999; Bago et al., 2001; Hawkins and George, 2001), in addition to significant amounts of N-organic and amino acids (Hawkins et al., 2000); however, to date, only a high-affinity NH4+ transporter expressed in extraradical AMF hyphae has been characterized (López-Pedrosa et al., 2006). Functional AM can generate important changes of pH in the mycorrhizosphere, as has been detected in various assays using in vitro culture systems (Bago et al., 1996; Bago and Azcón-Aguilar, 1997). It has been observed that the development of mycelium in the presence of NO3- is associated with a pH increase, and the opposite occurs in the presence of NH4+. No concordance exists with respect to the effect of fertilization with different N-sources on mycorrhizal plants: some studies suggest their preference for NO3- (Azcón et al., 1992), while others suggest their preference for NH4+ (Cuenca and Azcón, 1994), with genotypical variations being able to exist within the same plant species, mainly in limited N environments (Nakamura et al., 2002). The aims of the present study were: i) to determine the effect of two N fertilizers, such as urea and sodium nitrate, and two commonly cropped wheat cultivars on the persistence of AMF propagules in an Andisol, and ii) to analyze the interactive effect of both factors on the changes of some soil chemical characteristics. MATERIALS AND METHODS Experimental design. A 2 x 2 full factorial randomized experiment design was used, including two wheat cultivars commonly used in the study zone (‘Otto’ and ‘Metrenco’) and two N sources (N-NH4+ and N-NO3-). Each treatment combination had four replicates for each of two measurement stages. Soil characteristics. The test soil used was collected from a 5 to 25 cm depth of an annual crop site in the commune of Vilcún (38º41` S; 72º24` W), Region of the Araucanía, Chile. Some characteristics of the soil, an Andisol of the Vilcún series (Entyc Dystrandept), are presented in Table 1. Soil was air-dried, sieved through a 5 mm mesh, and supplied with 0.06 g P kg-1 soil as Triple Superphosphate and 0.063 g K kg-1 soil as KCl in solution. One liter pots were filled with 800 g of soil. Table 1. Selected chemical and microbiological characteristics of the Vilcún series soil (Entyc Dystrandept) used in this study.
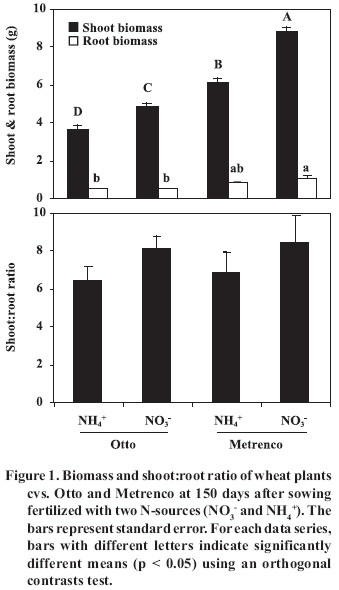
CEC: cation exchange capacity; AMF: arbuscular mycorrhizal fungi. Plant material. Wheat cvs. Otto and Metrenco were used as host plants. Seeds of each cultivar were surface sterilized with a 2% Cloramin-T (CH3C6H4SO2N(Cl)Na x H2O) solution for 3 min and rinsed thoroughly with distilled water. Four seeds were germinated between wet tissue paper and planted 7 d later. Pots were thinned to two plants after establishment. Growth conditions. Plants were grown under greenhouse conditions with temperatures ranging from 25 ± 3 ºC day-time to 15 ± 3 ºC night-time, a 16:8 h light:dark photoperiod and a relative humidity of 80-90%. A photosynthetic photon flux density of 0.4-0.5 mmol m-2 s-1 was applied as supplementary light when necessary. Plants were irrigated manually with distilled water as needed during the experiment (judged by weighing the pots). Every 2 wk, 10 mL of a nutrient solution (Johnson et al., 1996), without P and N, was added to each pot. The N was supplied twice, at establishment (30% total N in Zadocks 11 stage) and at 6 wk of cultivation (70% total N in Zadocks 31 stage) (Zadocks et al., 1974) to an equivalent of 0.126 g N kg-1 soil, which represents a normal fertilization rate equivalent to 200 kg N ha-1 . In the NO3- treatments, NaNO3 was used; whereas CO(NH2)2 was used in the NH4+ treatments. In both cases, N was supplied in solution. Harvest and analyses. Two harvest stages were considered. The first stage was at dry grain (150 days after sowing (DAS), at Zadocks 99 stage), and the second stage was at 240 DAS (3 months after grain harvest). Only soil and roots were present in the pots in the last stage, and they had been watered once a week since grain harvest. The harvested roots and shoots were dried at 65 ºC for 48 h in a forced-air oven (Memmert Gmbh + Co., Mod. UIM 400, Schwabach, Germany) and weighed. Before drying, 1 cm root fragments were separated and AM colonization was estimated by the method described by Giovanetti and Mosse (1980), after clearing and staining with Trypan blue (Phillips and Hayman, 1970). Root length was determined by the gridline intersect method (Tennant, 1975). The colonized root was determined at 150 and 240 DAS. The total extraradical mycorrhizal hyphae were determined by the method described in Borie et al. (2000), and the active hyphae were determined using dehydrogenase activity (Sylvia, 1988; Kabir et al., 1997). Total and active hyphal density were quantified using the intersect gridline method (Newman, 1966). Mycorrhizal spores were separated from soil by wet sieving and decanting in a 70% w/v sucrose solution (Gerdemann and Nicholson, 1963); spores were then quantified under a stereoscopic microscope at 50X. Extraradical hyphae and mycorrhizal spores were determined at 240 DAS. Soil pH was measured in a soil:water mix (2:5), and available P in the soil by the method described by Olsen and Sommers (1982) after extraction with 0.5 M NaHCO3 (pH 8.5). Soil pH and available P were determined at 150 and 240 DAS. Statistical analyses. The main effects of wheat cultivar, N source and their interaction were tested by means of a two-way analysis of variance (anova) using the general linear model procedures of the SPSS software, version 10.0 (SPSS Inc.) (Pérez, 2001). Means were compared by the orthogonal contrast test (Petersen, 1977). Data sets not meeting assumptions for anova were transformed as required, but the results were presented in their original scale of measurement. The analysis of the relationship between different variables was carried out by lineal correlation. Statistical significance was determined at p < 0.05. RESULTS AND DISCUSSION The wheat cultivars and the N-sources used as fertilizers influenced the behavior of most of the analyzed variables (Table 2). In general, variables significantly affected by one factor were also affected by the other, as well as by their interaction. The shoot and root biomass production was greater in Metrenco cultivar, in comparison to Otto cultivar (Figure 1). The NO3- use was related to a biomass increase in both cultivars, the Metrenco + NO3- treatment registering a shoot biomass 150% higher than the Otto + NH4+ treatment. These differences could be related to different genotype and phenotypic characteristics of cultivars, as is the case of root length or biomass. Previous observations of Nakamura et al. (2002), working with native and hybrids sorghum (Sorghum bicolor (L.) Moench) cultivars, suggest that greater growth and radical activity are the main determining factors of higher biomass production by a particular cultivar. In the present study, the different cultivars presented differences in shoot biomass production (p < 0.01), independent of the N-source used, and similar shoot:root ratios in both cultivars were observed, although the use of NO3- was associated with moderate increases in this ratio (Figure 1). Table 2. F-values and probabilities of significance for the main effects and factor interaction for the variables analyzed by means of a two-way anova at 150 and 240 days after sowing (DAS).
AM,
arbuscular mycorrhizal; ns,
not significant; *p < 0.05; **p < 0.01; ***p
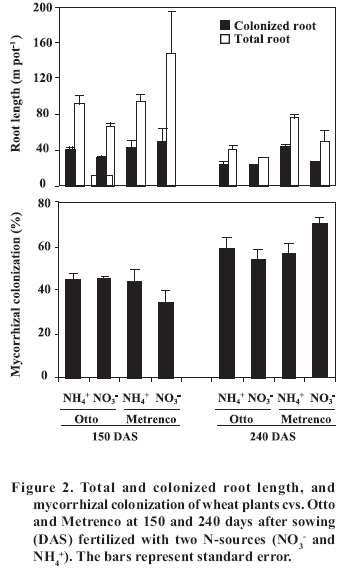
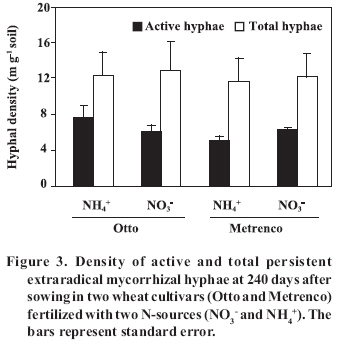
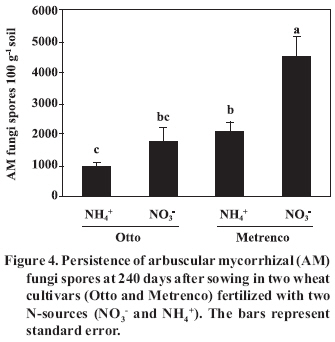
< 0.001; Total and AM colonized root length did not present significant differences in any of the combinations studied at 150 DAS (Figure 2). However, it was observed that a greater amount of roots in cv. Metrenco (78.6% more than cv. Otto) at 240 DAS remained persistent, especially with NH4+ use. In contrast, the mycorrhizal colonization was not affected by the analyzed factors in any of the studied stages, although an important increase of the mycorrhizal colonization at 240 DAS with regard to the previous stage was observed (Figure 2). The effect of seasonal changes in levels of colonization has been observed in other studies, although with different behavior. Thus, Kabir et al. (1997) found that higher levels of colonization appeared in summer, coinciding with more advanced development stages of maize (Zea mays L.) plants. On the other hand, Cornejo et al. (2007), in a greenhouse study carried out under similar conditions to the present ones, observed differences in levels of colonization only in early stages of a wheat crop; in addition, colonization was found to be greater when NO3- was used. As in the present study, colonization levels increased after plant harvest, which probably does not indicate new colonization, but rather a greater prevalence of the mycorrhizal roots at the end of the plant life cycle. Therefore, we can suppose that differences in the mycorrhizal colonization were present in previous growth stages, not considered in the present study. However, the high prevalence of mycorrhizal roots is an aspect of special agro-ecological relevance, since the mycorrhizal root fragment pieces are an important AMF inoculum source for the succeeding crop in the rotation (Brundrett, 1991). A similar behavior was observed in the case of mycorrhizal colonization and persistent hyphae in the soil, and no significant differences between different combinations of cultivars and N-source were observed, nor were the total and active hyphae density or proportion of active hyphae (Figure 3). In a similar way, the study carried out by Cornejo et al. (2007) found, in a wheat crop, higher densities of active hyphae in NO3- fed soils only at dry grain harvest. This behavior could be determined by the AMF life cycle, a basal or residual hyphal density appearing in the first plant and symbiosis development stage, later being dependant on the intraradical colonization extension, finally becoming stabilized when the bidirectional nutrient exchange between plant and fungus stops because of vegetal senescence or, as in this study, because of the shoot harvest. In contrast, densities of persistent hyphae were 2-3 folds higher in comparison to those of the Cornejo et al. (2007) report, probably because of the contribution to this component of several AMF species native to the natural soil. Pringle and Bever (2002) found that the AMF community composition was modified over time due to the proliferation of different fungi species, independently of other factors analyzed, such as the floristic community composition. It is possible that the adaptation to specific situations by some AMF species, as in this case of different N-sources, the wheat cultivar used, or the different conditions and variations of soil characteristics, confers a high plasticity to the natural AMF communities, allowing them to reach similar densities of fungal propagules by the contribution of different species. Similarly, Munkvold et al. (2004) found a high intraspecific diversity in the mycelial growth of different AMF species, suggesting the existence of an important functional complementariness, including at low levels of fungal diversity. Thus, studies of diversity and population dynamics that present different species within the AMF communities become quite necessary. The fungus life cycle is completed with the production of resistance spores (Brundrett, 1991), which are able to generate new colonization. In this study, highly significant differences in the AMF spore density between the different treatments, attributable to both studied factors, were registered(Figure 4). ‘Metrenco’ presented higher spore density than ‘Otto’; in addition, the use of NO3- as N-source favored the formation of spores in both cultivars. The Metrenco + NO3- treatment produced 375% more spores than did the combination Otto + NH4+. No clear relationship between the persistent spore density and some other AMF propagule was found. However, shoot biomass production was high and directly related to spore production (r2 = 0.89; p < 0.001). This result could be based on the greater carbonated flux than presumably was obtained in plants with a greater growth, allowing a greater carbonated compounds contribution to intraradical fungal mycelium, which by glyconeogenesis in external mycelium would later be transformed into reserve lipids (Pfeffer et al., 1999; Bago et al., 2002), an important component of resistance spores. The results presented by Cornejo et al. (2007) suggest greater spore production in the final growth stages in annual crops, which could be based on the functionality of other AM components in previous stages of the fungal life cycle, such as intraradical colonization and subsequent soil colonization by extraradical hyphae. These results and observations are of significant agronomic and ecological interest, since a greater mycorrhizal propagule density will lead to a greater probability of generating new and early-effective fungal colonization in the succeeding crop in the rotation. In the case of soil-available P, we observed that the Metrenco + NO3- combination had significant differences only with regard to the Otto + NO3- combination (3 mg P kg-1 of difference between both) (Table 3). In particular, cv. Otto exhibited a significant difference between the N-sources used, the NH4+ use being associated with greater soil P availability. Previous studies have also reported greater P availability in soils fertilized with NH4+ sources relative to NO3- sources (Ortas et al., 1996). In the present study, the increase in P availability was associated with a greater colonized root length in ‘Otto’ at 150 DAS; a greater functionality of the established mycorrhiza could explain this greater P availability. However, Ortas et al. (1996) suggest that a greater P availability can result from a stimulating effect of NH4+ on the roots, producing a concomitantly greater organic acid exudation that would release P from different soil compartments, a mechanism that has been widely studied (Hoffland et al., 1989; Johnson et al., 1996; Montenegro and Zapata, 2002). Of the soil-root interface properties analyzed, only pH exhibited important differences by effect of the studied factors, these differences being higher at 150 DAS (Table 2 and 3). The pH values were greater in ‘Metrenco’, especially when NO3- was used; a difference of 0.69 units of pH at 150 DAS was measured with regard to the Otto + NH4+ combination. This difference persisted at 240 DAS. These results agree with previous observations demonstrating that the rhizospheric pH changes are due to the N-source used and to the mycorrhizal plant status (Vaast and Zasoski, 1992; Bago and Azcón-Aguilar, 1997). The observed alkalinization due to NO3- absorption by the roots (and the mycorrhiza) is well-known (Marschner, 1995; Bago and Azcón-Aguilar, 1997). Nevertheless, the effect that different wheat cultivars exhibited on pH can be explained by different biomass production. The greater growth achieved by ‘Metrenco’ could have made possible a greater absorption and a faster assimilation of NH4+, avoiding its use as a substrate in the nitrification processes that, as has been observed in volcanic soils in southern Chile, are able to induce strong pH reductions (Campillo and Rodríguez, 1984). In the present study, highly significant correlations between pH values and shoot biomass production (r2 = 0.83 at 150 DAS ; p < 0.001) were found, which could reinforce the previous observation. As well, a highly significant correlation between pH values and AMF spore production at 240 DAS (r2 = 0.80; p < 0.001) was found, although this could reflect an indirect effect of a greater biomass production of the Metrenco + NO3- treatment and a greater contribution of carbonated compounds than a direct influence of the soil acidity. Table 3. Soil-root interface traits at 150 and 240 days after sowing (DAS) in two wheat cultivars (Otto and Metrenco) fertilized with two N-sources (NO3- y NH4+). Cuadro 3. Propiedades de la interfase suelo-raíz a los 150 y 240 días después de la siembra (DAS) en dos cultivares de trigo (Otto y Metrenco) fertilizados con dos fuentes de N (NO3- y NH4+).
For each cultivation stage, means followed by the same letter in a column are not significantly different using orthogonal contrasts test (p < 0.05; n = 4). CONCLUSIONS The use of best adapted cultivars and/or those with a greater biomass production potential, as was observed in the case of Metrenco, can be associated with an increase in the mycorrhizal functionality, which will produce an increase in propagule density. On the other hand, the application of NO3- favored plant growth, which was related to a significant decrease of soil acidity, one of the commonest limiting characteristics of volcanic soils such as that used in this study. The improvement of soil characteristics using NO3- was directly related to higher persistence of AMF spores in the soil, even at similar levels of hyphal length and colonized roots, demonstrating its strong impact on the native AMF populations present in Andisols in southern Chile. In summary, the use of suitable agricultural practices allows greater plant biomass production, an important improvement of soil chemical conditions and superior persistence of mycorrhizal propagules, particularly spores, which will produce earlier effective plant colonization in the succeeding crop in the rotation. This last aspect is very important from a realistic point of view, considering that these kinds of soils are used mainly in annual rotations, providing supplementary tools to farmers for a better choice of N-source. ACKNOWLEDGMENTS This work was supported by FONDECYT, grant Nº 1990756, from Comisión Nacional de Investigación Científica y Tecnológica (CONICYT), Chile. LITERATURE CITED
Copyright 2008 - Instituto de Investigaciones Agropecuarias, INIA (Chile). The following images related to this document are available:Photo images[cj08011f2.jpg] [cj08011f3.jpg] [cj08011f1.jpg] [cj08011f4.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}