 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Chilean Journal of Agricultural Research (formerly Agricultura Técnica), Vol. 68, No. 2, Apr-Jun, 2008, pp. 183-191 Scientific note Pistillate Flower Abscission Symptoms of ‘Serr’ Walnut (Juglans regia L.) Sintomatología del aborto en flores pistiladas de nogal (Juglans regia l.) ‘SERR’. Cristián González R.[1]y Gamalier Lemus S.1*, y Gabino Reginato[2] [1] Instituto de Investigaciones Agropecuarias, Centro
Regional de Investigación La Platina, Casilla 439/3, Santiago, Chile. E-mail: csalez123@gmail.com; glemus@inia.cl *Author
for correspondence. Received: 2 August 2007. Code Number: cj08018 ABSTRACT Pistillate flower abscission (PFA) severely affects walnut (Juglans regia L.) production, mainly of the cultivar ‘Serr’. A study on symptoms of flower abortion in walnut ‘Serr’, induced by excess or lack of pollen, was carried out following the evolution of flowers and peduncles. Hand pollinated flowers, with different pollen concentrations: 0, 1, 5, 25, 50 and 100% pollen diluted (w/w) in talcum powder, showed external and internal abortion symptoms. External symptoms were recorded directly from the tree, while for internal symptoms, flower excisions stained with blue Toluidin-O at 0.05%, were analyzed under an optical microscope. Excess pollen abortion produced flowers with rapid damage. The growth rate was very low, involucre brightness was lost and the stigma turned necrotic, in contrast to the flower aborted from lack of pollen. Normal flowers continued growing to fruit harvest, without losing brightness. Excess pollen abortion induced flower drop, with or without attached peduncle. Two separation zones can be present: the distal and proximal areas of the peduncle. Peduncle scars allowed for differentiating abortion induced from excess or lack of pollen, or other reasons. Key words: lack of pollen, excess of pollen, pollination, Juglans regia. RESUMEN El aborto en flores postiladas (PFA) por exceso de polen que afecta al nogal (Juglans regia L.), disminuye considerablemente la producción de nueces, principalmente en el cultivar ‘Serr’. Se estudió la sintomatología del aborto por exceso y falta de polinización en flores y pedúnculo de este cultivar. Para esto se realizó polinización manual, utilizando diferentes concentraciones de polen sobre flores en diferentes estados de receptividad. Las concentraciones se obtuvieron mediante diluciones (peso/peso) de polen combinado con talco. Las concentraciones utilizadas fueron 0; 1; 5; 25; 50 y 100% de polen. Se observó la sintomatología externa e interna de la flor, mediante apreciación visual en el árbol y a través de cortes longitudinales en la flor para observación en microscopio óptico, utilizando como tinción azul de toluidina-O al 0,05%. La flor abortada por exceso de polen se deterioró rápidamente, presentó detención del crecimiento, pérdida del brillo del involucro y stigmatic necrosis. La flor abortada por falta de polinización mostró un deterioro lento que se prolongó dos semanas más que la abortada por exceso de polen. La flor cuajada siguió creciendo, sin perder su brillo. La flor abortada por exceso de polen puede caer con o sin pedúnculo adherido. Se pueden presentar dos zonas de separación: zona distal y zona proximal del pedúnculo. La cicatriz en el pedúnculo permite diferenciar el aborto por exceso de polen, falta de polen y por otras razones. Palabras clave: falta de polen, exceso de polen, polinización, Juglans regia. INTRODUCTION In walnut cv. Serr, and to a much lesser degree among other cultivars, fruit production in some seasons is low, mainly because of two factors: the abscission of pistillate flowers owing to a high density of pollen in the orchard, and/or flower drop because of the lack of pollination. Hendricks et al. (1985), Catlin et al. (1990), Polito et al. (1998) and Lemus et al. (2001) reported that both factors are subject to the walnut flowering dynamic and environmental conditions that affect this. The accumulation of winter chilling and the age of the trees are factors that influence the degree of overlap of female and male flowering, and in consequence, in the level of abscission that can occur because of excess or lack of pollen, a situation that can hinder the annual expected production in every orchard (Hendricks et al., 1985; McGranahan et al., 1994; Polito et al., 1998; Lemus, 2005). Pistillate flower abortion (PFA) is understood as the loss of flowers early in the season. According to Polito et al. (1998) this loss is provoked by an excess of pollen in the stigmas, causing decomposition and the production of ethylene, which spreads to the rest of the flower, generating high levels of this hormone and provoking deterioration and harm to vegetal tissue. Research by Rovira and Aletá (1997) and Polito (1998), in field conditions, indicates losses of up to 90% by abscission owing to excess pollen, noting that this situation varies widely among varieties, sites and years. According to Krueger (2000) losses because of PFA are generally greater than losses because of lack of pollination. Evaluations made in Chile during the 2005-2006 season indicate that abscission owing to excess pollen occurs in a range of 40 to 90%, affecting the production (Lemus, 2005); on the other hand, abscission owing to lack of pollen does not exceed 5%, which is confirmed by Krueger (2000). The problem of abscission owing to excess pollen is important in cv. Serr, and to a lesser degree in ‘Hartley’ and ‘Chandler’ (Catlin et al., 1990; McGranahan et al., 1994; Lemus, 2005; González, 2006). It was recently reported that the application of growth regulators that inhibit the synthesis of ethylene, such as aminoethoxyvinylglycine (AVG) reduce the problem of abscission owing to excess pollen (Lemus et al., 2007). Other alternatives that have been evaluated are shaking trees with the objective of eliminating part the plant’s catkins (Lemus, 2005); currently the growth regulator 1-methylclyclopropene (1-MCP) is being tested. It acts at the level of ethylene receptors, inhibiting their action. (Lemus, 2005). The hypothesis of this experiment is that the symptomology of the aborted flower, both due to excess and the lack of pollen, is characteristic of the respective type of abscission, which allows for distinguishing both events when they have occurred. The objective of this study was to describe the external and internal symtomology of pollinated and non-pollinated flower, in comparison to the flower that develops normally without aborting. MATERIALS AND METHODS Location of the study The experiment was carried out during the spring of 2004, in the walnut orchard of the Los Tilos Experimental Field Station (33º44’ S; 70º38’ W; 513 m.a.s.l.) of the Instituto de Investigaciones Agropecuarias (INIA), located in the community of Buin, the Metropolitan Region of Chile. Adult trees of the Serr variety were used, grafted from J. regia in 1983, planted at a distance of 6 x 6 m. The trees were pruned on a central axis and irrigated by furrows. 48 trees were selected; a branch from the mid-level of each tree was marked, on East face, and 20 pistilate flowers. Pollen concentrations During the phenological period of pollen release, termed Fm and Fm2 (Solignat cited by Gil-Albert, 1996), which represents the beginning and end of anther dehiscence, respectively, pollen was collected shaking the catkins in glass flasks; the pollen was then diluted with talcum powder (w/w), in order to obtain different concentrations to carry out manual pollination tests. The concentrations used were 0, 1, 5, 25, 50, and 100% (weight/weight). To estimate the quantity of pollen applied, a counting was made by application or load with a hematocymeter (HerkaR, Neubauer, Germany) under an optical microscope (Carl Zeiss, Zeiss, Germany). A load of pollen by concentration was used, with five replications. Each load was diluted on two drops of distilled water, and four readings were made of the visual field. In this way, by averaging the number of pollen grains counted in each reading, estimation was made of the equivalence between the number of pollen grains per mm2 of visual field and the concentrations applied. Pollen application Pollen application was done on pistillated flowers in the receptive phenological states Ff1, which represents the beginning of the separation of the stigma, and Ff2 which corresponds to the separated and pale green stigma: full female flowering, according to Solignat as cited by Gil-Albert (1996). For pollination, the flowers were covered with cotton and gauze in the pre-receptive state. The 20 pistillate flowers were pollinated when they reached the indicated state. Following pollination, the flowers were again covered with cotton to avoid contamination by pollen external to the study. Pollination was done by depositing a load of pollen on the stigmas of each flower using the eraser end of pencils, considered as the contact surface of a known area. The degree of covering was superficial, that is to say, without agglomeration of pollen. Figure 1 shows the state of development of the pistillate flowers used in this study. The number of aborted flowers was evaluated weekly, and the presence of the peduncle attached to the flower was determined visually, and the presence of stigmatic necrosis upon removing the cotton. The zones of abscission were observed. The fruit was considered set when the involucre had a diameter greater than 7 mm. Study of the pistil Longitudinal cuts were made with a scalpel in order to observe the pistils. Subsequently, each excised piece was placed in Toluidin-O blue stain at 0.05% for 5 min; and then observed under an optical microscope. The internal symptomology of the pistillate flowers was observed with this procedure. Experimental design A randomized block design was used with a factorial structure (6 x 2). The factors corresponded to pollen concentrations (6) and development stages of the pistillate flower (2). Four blocks were considered according to trunk perimeter: 82 - 110, 70 - 74.5, 60 - 67.5, and 45 - 53.5 cm. The study consisted of 12 treatments and four replications; each replication represented a branch with 20 pollinated flowers per tree. Abscission, the presence of the peduncle attached to the flower and stigmatic necrosis were evaluated in percentages and analyzed by variance analysis. The multiple comparison Tukey test was used to separate the means. The statistical program Stat Graphics (2.1) (Statistical Graphics Corporation, 1999) was employed. A level of significance of 5% was used in all of the analyses. RESULTS AND DISCUSSION Pollen count The number of pollen grains applied per mm2 of surface varied from 0 to 111 for the different concentrations of pollen applied (Table 1). Table 1. Pollen density on the contact surface (grains mm-2) for each evaluated pollen concentration.
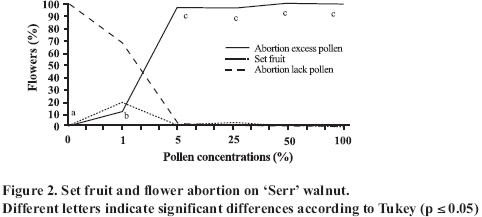
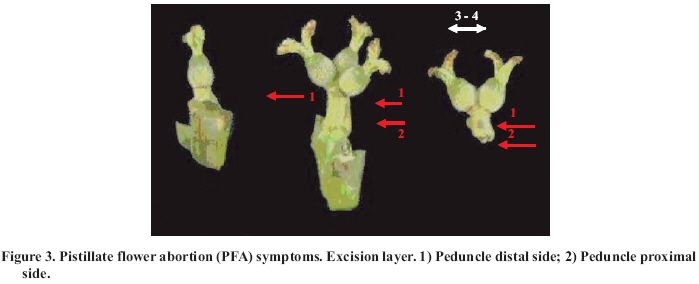
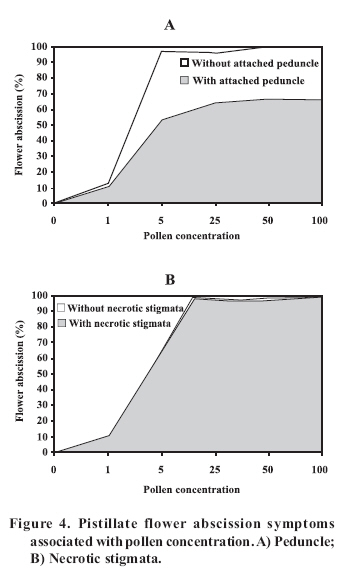
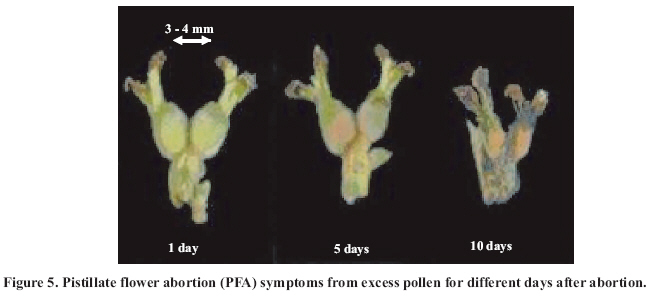
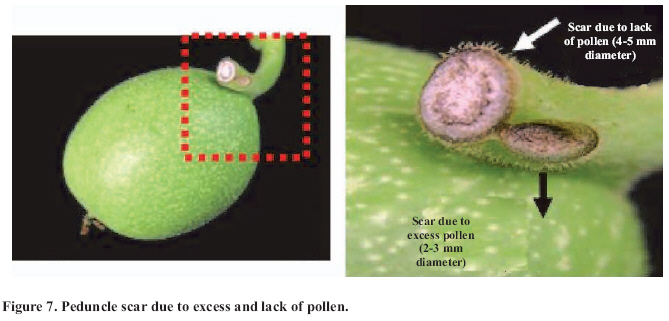
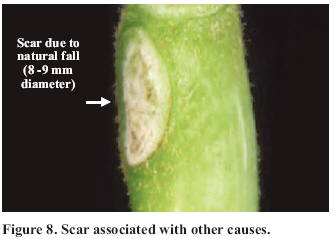
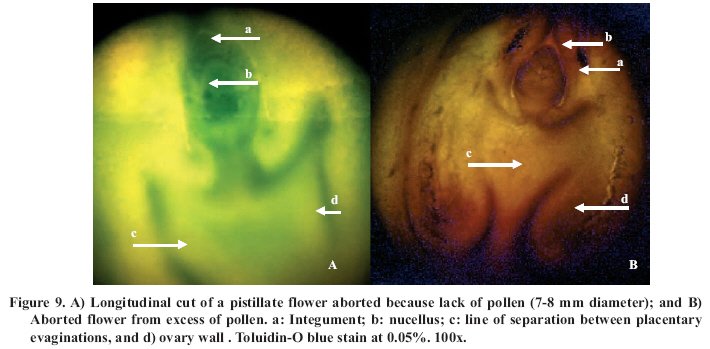
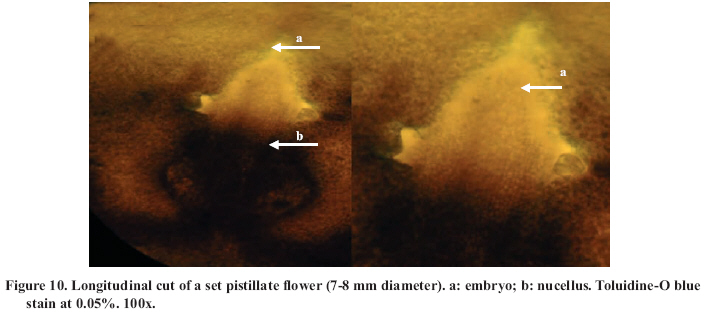
Abscission of the pistillate flower in two states of development at different pollen concentrations. Abscission from excess pollen appeared early in the season, in effect, 3 to 4 weeks following the beginning of receptivity, around the 20th of October. On the other hand, abscission from lack of pollen was evident 1 to 2 weeks after abscission from excess pollen was observed. This concurs with what has been described by Catlin et al. (1987); McGranahan et al. (1994) and Polito (1998). Increasing abscission of flowers was observed as the concentration of pollen was increased. At 5% of pollen (10 grains mm-2 of applied surface) 96.9% of abscission was reached, and the maximum setting was presented with 1% de pollen (5 grains mm-2) (Figure 2). This result confirms that cv. Serr requires a low concentration of pollen for setting to occur without floral abscission. Significant differences were observed in abscission from excess pollen of between 0 and 1%, according to the multiple range Tukey test (P ≤ 0.05). There was also a difference between 0% and any concentration equal to or greater than 5% (Figure 2). Upon analyzing the effect of 5, 25, 50 and 100% of pollen, no differences in the magnitude of abscission were observed, from which it can be concluded that for cv. Serr applications of between 0 and 5% of pollen favor setting, without provoking floral abscission from excess pollen. The different pollinated floral stages did not present significant differences, which suggest that the intensity of abscission is independent of these periods or states of pollination. Given this, any monitoring of the quantity of pollen grains should be done before and during the periods of receptivity of the pistillate flower. Symptomology caused by excess pollen It was observed that the flower aborted from excess pollen stopped growing and fell, having a diameter of 3 to 4 mm (Figure 3). This concurs with what was described by McGranahan et al. (1994), Polito (1998), Krueger (2000) and Lemus et al. (2001). As well, those authors indicated that the pistil grows to a diameter of 0.3 to 0.5 mm. As well, two separation areas were observed: the first in forming, and not described in the literature, was found in the distal area of the peduncle, and the second in the proximal area of the peduncle (Figure 3). The distal separation area of the peduncle was present in 36% of the cases, causing flower drop in absence of the peduncle. This can be confusing if the absence of the peduncle is attributed to abscission from lack of pollen. The remaining 64% presented the attached peduncle, by the effect of proximal separation area (Figure 4). Of the total aborted flowers from excess pollen, 98.6% presented stigmatic necrosis (Figure 4), which concurs with what was described by Catlin et al. (1987), McGranahan et al. (1994) and Polito (1998), for flowers that have received an excess of pollen (PFA). A loss of color and brightness was observed in flowers aborted from excess pollen. At ten days after dropping, the flowers are dehydrated, with a ligneous texture. This is the situation normally observed in seasons with high levels of pollen (Figure 5). Symptomology caused by lack of pollen In relation to the non-pollinated flowers, 94.1% fell with an approximate size of 7-8 mm in diameter. Some 98.7% of the flowers did not have the peduncle attached and distal separation area of the peduncle was not necrotic, unlike the flowers aborted from excess pollen (Figure 6). Consequently, it is the case that with inflorescence a flower drops because lack of pollen and another from excess, an aspect that can be seen with or without the presence of necrosis in the separation area. The stigmas of flowers dropped from lack of pollen are dry, but maintain their pale green color and the pistil is completely expanded. The set fruit had completely dry and brown stigmas (Figure 6). This concurs with the description by McGranahan et al. (1994), Polito (1998), Krueger (2000) and Lemus et al. (2001). The fruit dropped from lack of pollen reached a uniform size, a characteristic that facilitates diagnosis in the field. The contact surface, size and color of the scar in the peduncle allowed for differentiating both types of abscission. The scar caused by excess pollen has an irregular surface, is 2 to 3 mm in diameter and is brown. On the other hand, the scar caused by lack of pollen is chalky, has a smooth surface and is 4 to 5 mm in diameter. Both types of scar can be observed in Figure 7. Another scar was later observed produced by the fruit drop owing to other causes. This was similar in terms of the contact surface and color to that observed for drop from lack of pollen. However, it was 8 to 9 mm in diameter, and clearly the dropped fruit were of a larger diameter to those dropped from lack of pollen. This type of scar is often found in years of high load (Figure 8). Study of the pistil The presence of placental evaginations was observed in the two types of aborted flowers, but without necrosis in those with symptoms of abscission owing to excess pollen (Figure 9). This does not concur with what was found by Catlin et al. (1989) and Lemus et al. (2001), who reported that necrosis of placental evaginations allows for differentiating abscission from excess pollen from that caused by lack of pollen. The placental evaginations can be considered temporal storage structures, comparable to the placenta of the tomato (Lycopersicon esculentum Mill.), which disappear once the embryo is extended, a situation that also applies to the female flowers of the walnut. Nevertheless, the impossibility of observing necrotic zones within the flower could be due to excessively thick tissue. Consequently, other techniques are recommended that could allow for confirming this result. It was also observed that some flowers with symptoms of setting (7-8 mm) presented embryonic growth (Figure 10). Consequently, completely dry and brown stigmas are symptoms that allow for differentiating set flowers from unpollinated ones. CONCLUSIONS Under the conditions of the present study, it is possible to conclude that:
LITERATURE CITED
Copyright 2008 - Instituto de Investigaciones Agropecuarias, INIA (Chile). The following images related to this document are available:Photo images[cj08018f7.jpg] [cj08018f6.jpg] [cj08018f5.jpg] [cj08018f1.jpg] [cj08018f4.jpg] [cj08018f8.jpg] [cj08018f10.jpg] [cj08018f2.jpg] [cj08018f3.jpg] [cj08018f9.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}