 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Chilean Journal of Agricultural Research (formerly Agricultura Técnica), Vol. 69, No. 1, Jan-Mar, 2009, pp. 112-118 Scientific Note ENCAPSULATED SOMATIC EMBRYOS AND ZYGOTIC EMBRYOS FOR OBTAINING ARTIFICIAL SEEDS OF RAULI-BEECH (Nothofagus alpina (Poepp. & Endl.) Oerst.) Encapsulado de embriones somáticos y embriones cigóticos para obtención de semillas artificiales de raulí (Nothofagus alpina (Poepp. & Endl.) Oerst.). Priscila Cartes R.1*, Hermes Castellanos B.2, Darcy Ríos L.1, Katia Sáez C.3, Scarlette Spierccolli H.1, y Manuel Sánchez O.1 1 Universidad de Concepción, Facultad de Ciencias
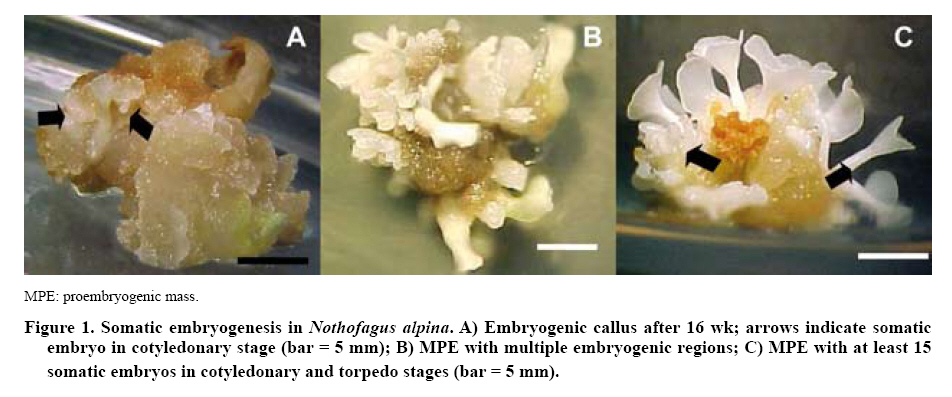
Forestales, Casilla 160-C, Concepción, Chile. E-mail: Received: 11 October 2007 Code Number: cj09014 ABSTRACT Somatic and zygotic embryos from mature seeds of rauli-beech, Nothofagus alpina (Poepp. & Endl.) Oerst., were encapsulated in different artificial endosperms in order to generate a cover that fulfills the function of nourishment and protection of the embryos, facilitating their later germination. The content of sodium alginate varied by 4%, 3%, and 2%, as did the immersion time in calcium chloride (CaCl2), which acts as complexing agent. The artificial endosperm components of the Murashige and Skoog medium (MS) were added, supplemented with 0.5 mg L-1 indolacetic acid (IAA), 0.5 mg L-1 naphthaleneacetic acid (NAA), 2 mg L-1 6-benzylaminopurine (BAP) and 30 g L-1 sucrose. The germinative behaviors of encapsulated somatic and zygotic embryos were evaluated after 4 wk. Comparing the percentages of germination reached by encapsulated somatic and zygotic embryos it was observed that they had similar germinative behavior according to the type of encapsulation applied. However, zygotic embryos substantially exceeded the germination levels reached by somatic embryos, 100% vs. 45% respectively. Key words: rauli-beech, somatic embryogenesis, synthetic seed, sodium alginate. RESUMEN Embriones somáticos y cigóticos provenientes de semillas maduras de raulí, Nothofagus alpina (Poepp. & Endl.) Oerst., se encapsularon en diferentes endospermas sintéticos con el fin de generar una cubierta que cumpla la función de nutrir y proteger al embrión para facilitar su posterior germinación. Se varió el contenido de alginato de sodio al 4%, 3% y 2% y el tiempo de inmersión en cloruro de calcio (CaCl2), el que actúa como agente acomplejante. Además, a la matriz artificial se adicionaron componentes del medio Murashige y Skoog (MS) suplementado con: 0,5 mg L-1 de indolacetic acid (IAA), 0,5 mg L-1 de ácido naftalenacético (NAA), 2 mg L-1 de 6-bencilaminopurina (BAP) y 30 g L-1 de sacarosa. Al cabo de 4 semanas el porcentaje de germinación de los embriones somáticos y cigóticos encapsulados tuvieron similar comportamiento germinativo según el tipo de encapsulado aplicado. Sin embargo, los embriones cigóticos superaron ampliamente los niveles de germinación alcanzados por los embriones somáticos, 100% vs. 45%, respectivamente. Palabras clave: raulí, embriogénesis somática, semilla sintética, alginato de sodio. INTRODUCTION The forestry sector has been one of the most dynamic in the Chilean economy since the 1970s, based mainly on plantating Pinus radita D. Don and Eucalyptus globulus Labill. This has led to a diminished importance of native forests in the national productive sector (INFOR-CORFO, 1997). However, the National Forest Recovery and Forestry Promotion Law has encouraged research, improvements in native species and the maintenance of forest ecosystems in order to strengthen this resource (CONAF, 2008). The raulí-beechis a native Chilean species that has been recovering its ecological and economic importance in recent years after having suffered severe deterioration owing to indiscriminate extraction of the best individuals of forests, resulting in a decrease in the quality of the species today (Lara et al., 1996; Sánchez-Olate et al., 2000). The propagation of N. alpina by direct seeding raises difficulties because of the low tolerance of the species, which only grows successfully where mineral soils are exposed to light (Loewe et al., 1997; Donoso and Lara, 1999). As well, the low viability of the seeds, periodicity in seed production (high production in short cycles of 2 to 3 years) and insect attacks affect the development of this species (Burschel et al., 1976). Techniques of micropropagationor in vitro cultivation have emerged as alternatives for species that do not have the property of producing viable seeds, that is, species that cannot germinate and develop adequately in their natural environment (González et al., 2004). Among the micropropagation techniques is somatic embryogenesis or the regeneration of embryos based on the vegetative tissue of N. alpina as an efficient technique that allows for the mass propagation of selected genotypes, for productive and conservation purposes (Castellanos et al.,2004). The success of this technique depends on the development of a series of processes that influence the genotype of the mother or donating explant and the concentration of exogenous growth regulators, which in adequate combinations would allow for obtaining a determinant embryogenic response for the production of somatic embryos (SE) (Guerra et al., 2001). The SE can be immersed in a protective matrix constituting an artificial or synthetic seed, providing a convenient method for the propagation by cloning of elite plant varieties or species that are difficult to propagate in their natural environment (Fuji et al., 1987). The first indications of artificial seed propagation were reported in annual crops such as alfalfa (Medicago sativa L.) and sugar cane (Saccharum spp.). Currently, systems of artificial seed production have progressed substantially in this area, the most advanced being in seeding under ex vitro or field conditions, obtaining high percentages of conversion to plants (Fuji et al., 1987; Nieves et al., 2003). However, the germination rates and the subsequent conversion to plants of SE of different woody species are still low, owing mainly to deficient and asynchronic maturation of the embryonic pole, which makes the terminal stages of the process difficult (Tapia et al., 1999; Castellanos et al., 2004). Several researchers suggest that to control growth and facilitate the germination of SE, the synthetic endosperm can simulate an endosperm of sexual origin, containing one or several compounds such as: nutrients, growth regulators, anti-pathogens, herbicides, bio-controllers and bio-fertilizers, among others, with the aim of ensuring the conversion of the plant and its development in the field (Castillo et al., 1998; Kumar et al., 2004; Malabadi and Van Staden, 2005). The composition of the protective matrix should allow for the growth of the encapsulated embryo, providing mechanical resistance according to the available energy of the embryo, given that an excessively hard endosperm results in energy loss and weak or nil growth of the encapsulated ES (Jiménez and Quiala, 1998; González et al., 2004). The literature notes the use of different substances (agar, gelrite) to encapsulate ES, with the manipulation of the concentration of sodium alginate and the exposure time to the complexing agent calcium chloride reporting the best results of germination and conversion to plants in woody species (Patel et al., 2000; Maruyama et al., 2003; Utomo et al., 2008). This study seeks to define the composition of synthetic endosperm, varying the sodium alginate content and exposure time to the complexing agent, with the goal of generating a synthetic endosperm that allows for the normal germination of encapsulated embryos. As well, the germinative behavior of somatic embryos and encapsulated zygotic of N. alpina are compared to determine the influence of the type of embryo on the germinative response. MATERIALS AND METHODS Place of study The study was carried out in a vegetal tissue cultivation laboratory of the Forestry Sciences Faculty and the Biotechnology Center of the University of Concepción during 2006 and 2007 a period that covers the preparation of the initial material until the encapsulation and germination of the SE. The initial material was SE of N. alpina cultivated under axenic conditions obtained from proembriogenic mass (MPE) from the RaC01embryonic line (Raulí-beech, coyledonary explant, Clon 01). In accordance with established protocols for the species, based on embryogenic callus (Figure 1A) the continuous regeneration of MPE was generated MPEs (Figure 1B), from which the somatic embryos are isolated to encapsulate in a torpedo-cotyledonary state of development, with an approximate length of 3 to 4 mm (Figure 1C) (Castellanos et al., 2005). Components of the artificial matrix The synthetic endosperm or artificial matrix was composed of minerals and vitamins from the MS culture medium (Murashige and Skoog, 1962) supplemented by 0.5 mg L-1 of indolacetic acid (IAA), 0.5 mg L-1 of naphthalene acetic acid (NAA), 2 mg L-1 of 6-benzyl aminopurine (BAP), 2 mg L-1 of Fe-EDTA and 30 g L-1 of sucrose. Depending on the encapsulation treatment applied, agar is replaced by sodium alginate at 4% (E1), 3% (E2) and 2% (E3), according to Prewein and Wilhelm (2002), Ipekei and Gozukirmizi (2003) and Maruyama et al. (2003), respectively. The complexing agent of the capsules is applied through immersion in calcium chloride (CaCl2) at different concentrations and determined time intervals (Table 1). Finally, they were washed in sterile water for 40 min.
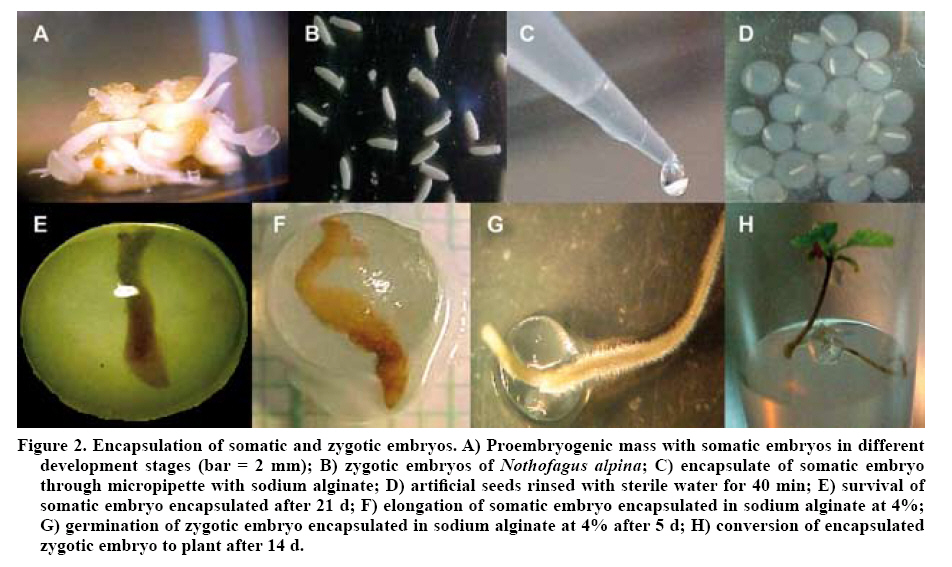
E1: type 1 encapsulate, sodium alginate at 4% and immersion in calcium chloride 15 g L-1 for 10 min; E2: type 2 encapsulate, sodium at 3% and immersion in calcium chloride 5,5 g L-1 for 30 min; E3: type 3 encapsulate, sodium at 2% and immersion in calcium chloride 14 g L-1 for 30 min. Encapsulation of somatic embryos The somatic embryos isolated from the mass (Figure 2A) were submerged in a solution of sodium alginate (gel), according to the type of encapsulation applied, and subsequently suctioned through a micropipette (Figure 2C) to provide a protective capsule. In order to seal the capsules, they were then submerged in a complexing solution of de CaCl2 for a determined period of time (Table 1) followed by washing in sterile water for 40 min (Figure 2D). This process was carried out under aseptic conditions in a laminar flow chamber, laminar with prior sterilization of the material and culture medium (matrix). Finally, the artificial seeds were cultivated in a germination medium in Petri dishes with macro and micronutrient from the MS medium supplemented with 30 g L-1 of sucrose and 7 g L-1 of agar-agar. They were then left in the culture chamber at a temperature of 25 ± 1 ºC in complete darkness. Encapsulation of zygotic embryos The seeds of N. alpina were soaked for 48 h in running water. The seed surfaces were then disinfected by submerging the seeds in a solution of ethanol at 70% (v/v) for 5 min, followed by washing in sterile water for 3 min. Subsequently the seeds were placed in a commercial solution of sodium hypochlorite diluted to 50% (v/v), following by three washings in sterile water for 3, 4 and 5 min, respectively. This process was carried out under constant agitation in a laminar flow chamber. The protective tissue was then removed from the seeds, under a stereoscopic magnifier and with the aid of pincers and a sterilized scalpel. In this manner bare embryos were obtained ready to proceed to encapsulation (Figure 2B). The encapsulation process was the same as that applied to the somatic embryos (Table 1). Finally, the artificial seeds were cultured in a germination medium in Petri dishes with macro and micronutrients from the MS supplemented with 30 g L-1 of sucrose and 7 g L-1 of agar-agar, which were left in the culture chamber at a temperature of 25 ± 1 ºC in darkness. Once the synthetic seeds germinated, they were transferred to the light, with a photoperiod of 16:8 (light:darkness) in the same growth chamber. After 3 weeks in darkness, an evaluation was made of the percentage of encapsulated of somatic embryos and that presented elongation of the hypocotyl-root axis and of the survival rate. Subsequently, at 4 weeks the percentage of germination of encapsulated somatic embryos was evaluated. The criterion used to evaluate the germination of the encapsulated embryos was the emergence of the root from the artificial capsule. The survival percentage of the zygotic embryos was evaluated and every four days the percentage of germination reached was evaluated, taking the emergence of the root from the capsule as a criterion. The experimental design was oriented to determine the best germinative behavior of encapsulated somatic embryos in different artificial matrices. To do this three encapsulation procedures were evaluated (E1, E2 and E3), with three repetitions of each. The experimental unit was a Petri dishes with six artificial seeds, generating 18 seeds per encapsulate type. The data obtained in percentages was submitted to arcsine transformation of the square root prior to the analysis. The effects of the treatments were evaluated using ANDEVA with a level of significance (α) of 0.05. When there was a significant difference among treatments a minimum significant difference of means test (MSD) was carried out, for which the statistical program SAS (SAS Institute, 2002) was used. To determine the behavior of the encapsulated zygotic and somatic embryos a completely random experimental design was used, with three treatments and four replications for each treatment. The experimental unit (Petri dish) was composed of six capsules generating a total of 24 artificial seeds per treatment. RESULTS AND DISCUSSION The levels of elongation of the hypocotyl-root axis and of survival of encapsulated somatic embryos at 21 days (Figure 2E and F) were higher by 70 and 90%, respectively, in all the treatments applied (Table 2). As well, there were no significant differences in the different percentages of sodium alginate used and complexing time in CaCl2 among the aforementioned variables. However, a response was presented similar to what was obtained by authors such as Castillo et al. (1998), who observed a higher percentage of germination of encapsulated somatic embryos with increased sodium alginate content and reduced exposure time to CaCl2 (Prewein and Wilhelm, 2002; Malabadi and Van Staden, 2005). This is because high concentrations or excessive exposure of the embryos to the complexing agent (CaCl2) results in more absorption and penetration of CaCl2 to the embryo, which can generate growth inhibition that is reflected in decrease in the germinative response and subsequent development in the field (Redenbaugh et al., 1986; Malabadi and Van Staden, 2005). Table 2. Effect of different treatments of encapsulating applied to somatic embryos of Nothofagus alpina.
E1:
type 1 encapsulate, sodium alginate at 4% and immersion in calcium chloride 15 g L-1 for 10 min; E2: type 2 encapsulate, sodium al 3% and immersion in calcium
chloride 5,5 g L-1 for 30 min; E3: type 3 encapsulate, sodium al 2%
and immersion in calcium chloride 14 g L-1 for 30 min. The importance of the high percentage of survival and germination obtained in the encapsulated zygotic embryos should be noted (Table 3). From these results it is clear that if the embryo reaches necessary vigor during the maturation phase, can break the mechanical resistance and lack of oxygen with increased sodium alginate content in the matrix, and thus the germinative response is not affected (Jiménez and Quiala, 1998). This can be extrapolated to somatic embryos, refining the maturation protocols to obtain better percentages of artificial seed germination (Nieves et al., 2001). Table 3. Effect of different treatments of encapsulating applied to zygotic embryos of Nothofagus alpina.
E1:
type 1 encapsulate, sodium alginate at 4% and immersion in calcium chloride 15 g L-1 for 10 min; E2: type 2 encapsulate, sodium at 3% and immersion in calcium chloride
5.5 g L-1 for 30 min; E3: type 3 encapsulate, sodium at 2% and
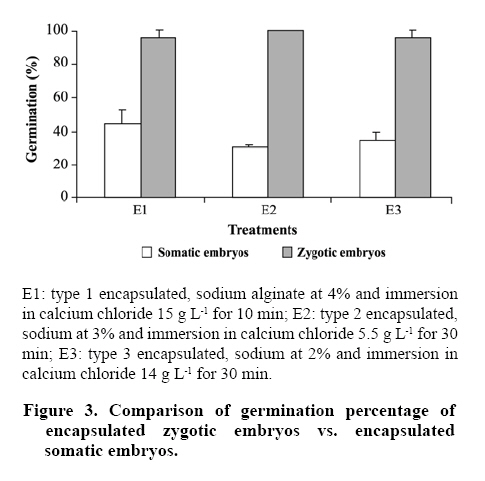
immersion in calcium chloride 14 g L-1 for 30 min. Comparing the germinative behavior of encapsulated somatic and zygotic embryos it can be observed that both types of embryos have similar behavior according to the encapsulation treatment applied (E1, E2 and E3). However, the zygotic embryos strongly surpassed the germination levels (100% for E2) reached by the somatic embryos (44% for E1) (Figure 3). Some authors suggest that the germination of ES is affected by the degree of vigor or maturity of the embryos at the moment of being encapsulated (Gómez, 1998; Nieves et al., 2001), given that the mechanical resistance that an excessively hard encapsulation can result in a major part of the available energy being utilized in breaking the synthetic endosperm (Jiménez and Quiala, 1998). It is important to point out that the zygotic embryos germinated at 4 days of cultivation (Figure 2G), achieving the conversion to plants (Figure 2H), while the somatic embryos germinated at approximately 4 weeks and in an irregular manner, manifesting a lack of maturity and asynchrony in the development of these embryos (Castellanos et al., 2004). However, refining the encapsulation process for somatic embryos, with the goal of improving the percentages of germination and subsequent conversion to plants, can mean the generation of a cloning product with the potential to outperform seeds of sexual origin (Tapia et al., 1999; Nieves et al., 2001). CONCLUSIONS The encapsulation procedure can be applied to both types of embryos of N. alpina, somatic and zygotic, allowing for the normal germination of embryos. There were significant differences in the percentage of germination reached for somatic embryos and zygotes according to the percentage of sodium alginate (2%, 3% and 4%) and the exposure time to the calcium chloride agent. The higher levels of germination were reached by the zygotic embryos in comparison to the somatic embryos, owing mainly to the lack of maturity and asynchrony of the embryoids. LITERATURE CITED
Copyright 2009 - Instituto de Investigaciones Agropecuarias, INIA (Chile). The following images related to this document are available:Photo images[cj09014f2.jpg] [cj09014f1.jpg] [cj09014f3.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}