 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
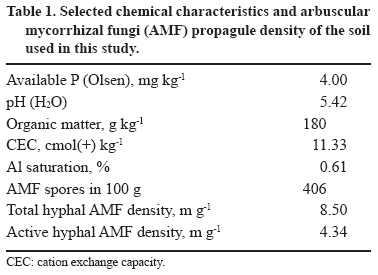
Chilean Journal of Agricultural Research, Vol. 69, No. 3, July-Sept, 2009, pp. 426-434 RESEARCH Mycorrhizal Propagule Persistence in a Succession of Cereals in a Disturbed and Undisturbed Andisol Fertilized with Two Nitrogen Sources Persistencia de Propágulos Micorrícicos en una Sucesión de Cereales en un Andisol Disturbado y no Disturbado, Fertilizado con Dos Fuentes de Nitrógeno. Pablo Cornejo1*, Rosa Rubio1, and Fernando Borie1 [1]Universidad de La Frontera, Departamento de Ciencias Químicas, Casilla 54-D, Temuco, Chile. *Corresponding author (pcornejo@ufro.cl). Received:
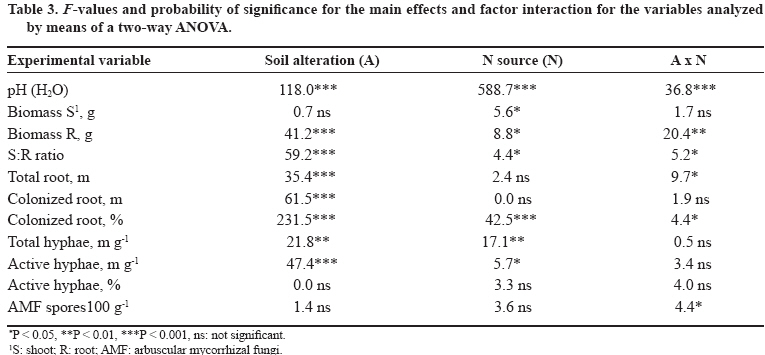
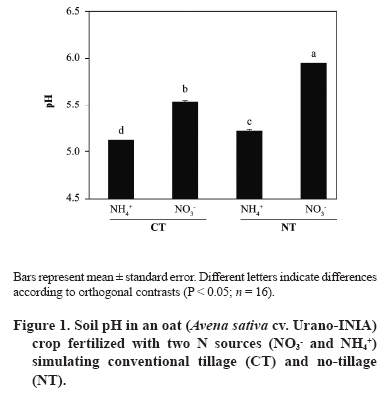
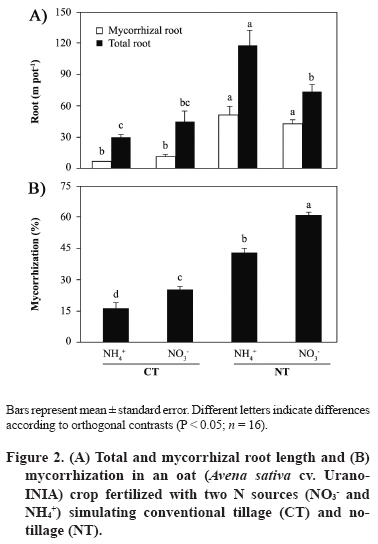
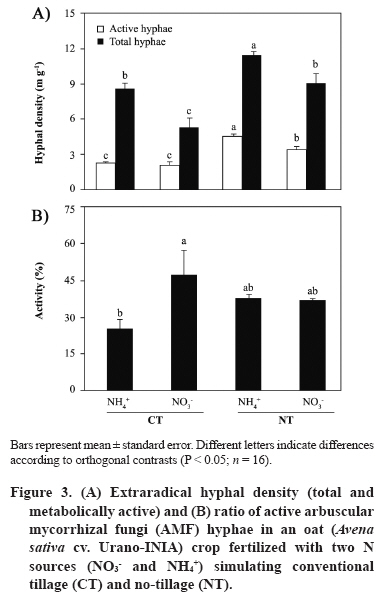
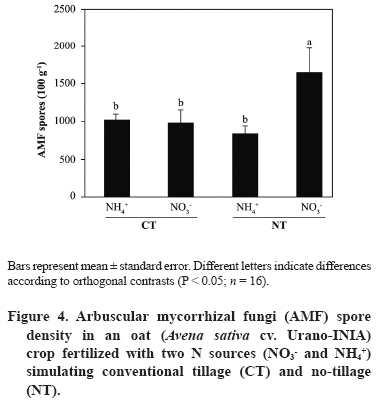
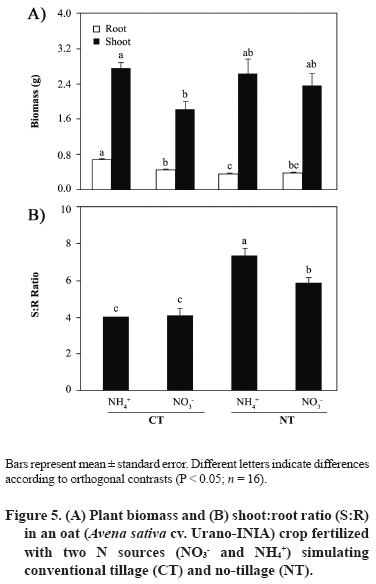
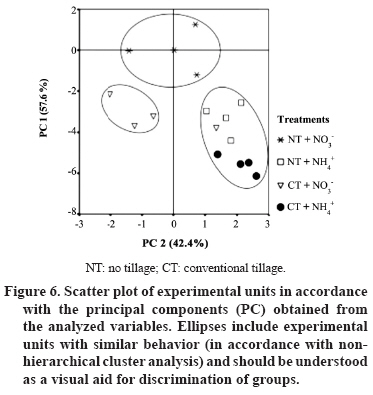
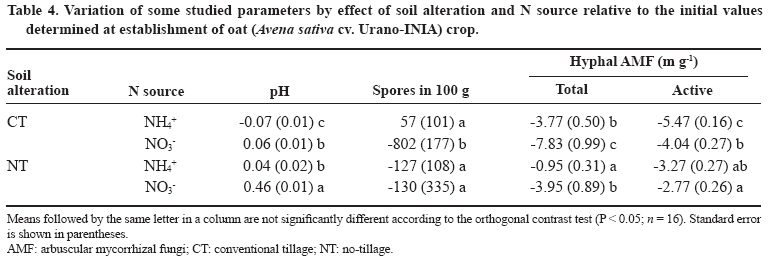
11 March 2008. Code Number: cj09051 ABSTRACT Arbuscular mycorrhizal fungi (AMF) form symbiosis with plant roots, improving its establishment, nutrition, and tolerance to adverse soil conditions. In annual crop rotations, some aspects such as the type of N fertilizer and tillage system used can affect the AMF propagule density and its functionality in the following crop. To analyze the effect of the agronomic practices previously mentioned on the persistence and density of AMF propagules, a study in a succession of cereals was carried out. For this, soil previously cropped with wheat (Triticum aestivum L.) and fertilized with urea (NH4+) or sodium nitrate (NO3-) was disturbed in each case by simulating conventional tillage (CT) or maintained without disturbing, simulating no-tillage (NT). It was then cultivated with oat (Avena sativa L.) using the same N sources. Higher densities of AMF active mycelium and colonized root length in the NT soil were observed (76% and 497% higher than in CT soil, respectively, P < 0.001). Moreover, NT + NO3- interaction produced a higher density of AMF spores (75% higher than in the rest of the treatments, P < 0.05), although showing a decrease in comparison with the previous wheat crop. These results suggest that N source and tillage system interact to influence persistence and density of AMF propagules, an important aspect to consider in acidic soils intended for annual crops. Key words: arbuscular mycorrhiza, mycorrhizal propagules, soil disturbing, soil pH, wheat-oat succession. RESUMEN Los hongos micorrícico-arbusculares (AMF) forman simbiosis con las raíces de las plantas, favoreciendo su establecimiento, nutrición y tolerancia a condiciones adversas del suelo. En rotaciones de cultivos anuales algunos aspectos como el tipo de fertilizante nitrogenado utilizado y el sistema de labranza pueden afectar la densidad de propágulos de AMF y su funcionalidad en el cultivo siguiente. Para analizar el efecto de las prácticas agronómicas previamente mencionadas sobre la persistencia y densidad de los propágulos de AMF, se realizó un estudio en microcosmos simulando una rotación de cereales. Para esto, suelo previamente cultivado con trigo (Triticum aestivum L.) y fertilizado con urea (NH4+) o nitrato de sodio (NO3-) fue, en cada caso, disturbado simulando una labranza convencional (CT) o mantenido sin disturbar simulando cero labranza (NT) y cultivado con avena (Avena sativa L.), utilizando las mismas fuentes de N. Se observaron mayores densidades de micelio activo de AMF y longitudes de raíz colonizada en el suelo NT (76% y 497% mayor que en suelo CT, respectivamente; P < 0,001). Por otra parte, la interacción NT + NO3- produjo una mayor densidad de esporas de AMF (75% mayor que en el resto de los tratamientos, P < 0,05), aunque mostrando una disminución respecto del cultivo previo de trigo. Estos resultados sugieren que la fuente de N y el sistema de labranza influyen interactivamente sobre la persistencia y densidad de propágulos de AMF, aspecto importante de considerar en suelos ácidos destinados a cultivos anuales. Palabras clave: alteración del suelo, micorriza arbuscular, pH del suelo, propágulos micorrícicos, sucesión trigo-avena. INTRODUCTION The volcanic soils of Southern Chile, mainly Ultisols and Andisols, show a natural tendency to acidity due to its particular mineral composition with high allophane and high organic matter content (Zunino and Borie, 1985). These soils cover an area over 5 x 106 ha, mainly used in annual cereal and legume rotations. Conventional tillage in these soils with plowing and harrowing have been widely practiced, altering soil conditions, microorganism activity and growth, and nutrient dynamics (Alvear et al., 2005; Borie et al., 2006). It is well-known that soil microbiota plays a fundamental role improving productivity and stability of natural and agricultural ecosystems (Jeffries and Barea, 2001; Barea et al., 2005). Within this microbiota, stand out the arbuscular mycorrhizal fungi (AMF), which establish a mutualistic symbiosis called arbuscular mycorrhiza, with most of terrestrial plants. In this symbiosis, AMF benefit nutrient absorption and translocation to plant, particularly those less mobile in the soil, such as phosphate, ammonium, and numerous micronutrients, due to the extension of the radical system that represents the fungal hyphae in the soil (Clark et al., 1999; Barea, 2000; Cornejo et al., 2008a). In turn, the fungus receives carbonaceous compounds from plant photosynthesis (Pfeffer et al., 1999; Bago et al., 2002). It has been observed that plant production is significantly increased in mycorrhized plants compared to non-mycorrhized plants in acidic soils (Clark et al., 1999). By the above, adequate management of native AMF populations in these soils is an important aspect to consider in sustainable agricultural systems. Several studies have demonstrated that the functionality of mycorrhizal symbiosis in agrosystems is affected by practices such as the previous crop in the rotation, type of fertilizer used, and tillage system, influencing persistence and density of AMF propagules remaining in the soil (Borie and Rubio, 1999; Rubio et al., 2003; Borie et al., 2006; Cornejo et al., 2007; 2008b). However, among the most relevant aspects to be taken into account in acidic soils is the emphasis on the type of N fertilizer used, because the nitrate (NO3-) and ammonium (NH4+) ions have a considerable effect on the variations of rhizospheric pH, owing these ions are the most abundant, most absorbed, and most accumulated by the plant (Gerendás et al., 1997; Cornejo et al., 2008a). Moreover, the NH4+ nitrification process generates H+ among its products, so its use as a fertilizer can promote severe acidification processes, particularly in naturally acidic soils like those of volcanic origin in Southern Chile (Campillo and Rodríguez, 1984; Martens, 2001; Chu et al., 2005). Related to the aforementioned, it is known that AMF extraradical hyphae can absorb different N sources, both ionic and some organic forms, so that they can contribute significantly in the variations of mycorrhyzospheric pH (Bago et al., 1996; Bago and Azcón-Aguilar, 1997; Hawkins et al., 2000; López-Pedrosa et al., 2006; Cornejo et al., 2008b). In addition, the tillage system used can have a considerable effect on soil microbiota. In general terms, it has been observed that no-tillage or the use of conservation tillage systems benefit the size, diversity, and activity of soil microbial populations (Oehl et al., 2004; Alvear et al., 2005), and in the case of AMF, it would be related to greater root colonization, propagule density, and communities diversity of this type of fungi (Borie et al., 2000; 2006; Jansa et al., 2002). However, the joint effect that the type of N source and tillage system has on AMF propagule densities in annual rotation systems in volcanic soils from Southern Chile has not been studied. Therefore, the aim of this study was to analyze, under controlled greenhouse conditions, the effect of contrasting N sources applied as fertilizers and soil alteration on persistence and density of AMF propagules in a succession of cereals, simulating a crop rotation in an Andisol. Assuming that arbuscular mycorrhizal symbiosis plays an important role in plant growth and nutrition, it is necessary to know the basis for selecting agronomic management alternatives that increase persistence, density and activity of AMF propagules naturally present in volcanic soils from Southern Chile. MATERIALS AND METHODS Experimental design A factorial design 2 x 2 full randomized was used, including two N sources (NO3- and NH4+), each of which simulated tillage systems with (conventional tillage, CT) and without (zero tillage, NT) soil sieving in the experimental units. Hereafter, this simulation factor of the different tillage systems will be referred as soil alteration. In each case, four replicates were used for each established combination (n = 16). Soil characteristics The soil used was obtained in a field intended for annual crops in the commune of Vilcún (38º41’ S; 72º24’ W; 180 m.a.s.l.), La Araucanía Region, Chile. Some initial characteristics of this Andisol are shown in Table 1 (Temuco family, Vilcún series; mesic, Entyc Dystrandept). Soil was collected from a of 5-25 cm depth, air-dried, sieved at 5 mm, fertilized with 0.063 g P kg-1 as pulverized triple super phosphate and 0.063 g K kg-1 as KCl in solution, equivalent to a 100 kg ha-1 dose of each nutrient, similar to those used by farmers for cereal crops in these same soils. Sixteen 1 L pots with 800 g of this soil were prepared. Subsequently, wheat (Triticum aestivum L., cv. Otto-Baer) plants were established and fertilized with 0.126 g N kg-1 (30% in Zadocks 11 and 70% in Zadocks 31; Zadocks et al., 1974), equivalent to a 200 kg N ha-1 dose, using CO(NH2)2 as a source of NH4+ in half of the experimental units, and NaNO3 as a source of NO3- in the rest. At the end of this crop (dry grain, Zadocks 99), wheat plants were harvested and after 3 months, the soil of four experimental units fertilized with each N source was disturbed by sieving at 5 mm, simulating CT. Then pH and AMF propagule density (spores, total and active hyphae, and colonized root length) were determined in the sieved experimental units in order to know the initial value of the soil parameters at the time of establishing the second cereal in the designed rotation simulation (Table 2). Plant material Oat (Avena sativa L. cv. Urano-INIA) was used as a second AMF host in the simulated cereal rotation. Oat seeds were sterilized with a 2% w/v Chloramine-T (CH3C6H4SO2N(Cl)Na x H2O) solution for 3 min and were rinsed thoroughly with dH2O. Seeds were pre-germinated on sterile tissue paper moistened with dH2O and after 7 days, two seedlings of each experimental unit were established with homogeneous growth of 5-6 cm leaf length and 4-5 cm radical length. Crop conditions Plants, both wheat and oat, grew under greenhouse conditions at 25 ± 3/15 ± 3 ºC day/night temperatures, 16:8 h light:dark photoperiod, and relative humidity of 80-90%. Pots were watered manually with distilled water according to plant needs during the experiment (determined by pot weight). Every 2 weeks, 10 mL of nutritive solution were added (Johnson et al., 1996) without P and N, to compensate the possible microelement deficiencies due to limited soil volume. Nitrogen fertilizer was applied in two portions: at the establishment of the oat plants (30% in Zadocks 11) and at the half tillering stage (70% in Zadocks 22-24), to complete 0.063 g N kg-1, which represents a dose of 100 kg N ha-1. The N fertilizers were the same used for the previous wheat crop. Measurements Evaluations were carried out at the beginning of anthesis (Zadocks 60, 150 days after sowing). Roots and shoots were dried at 65 ºC for 48 h in a forced-air oven (UIM 400, Memmert GMBH + Co., Schwabach, Germany), and weighed. Before drying the roots, a portion was separated to determine the proportion of AMF colonized root (Giovanetti and Mosse, 1980) after clearing with a 10% w/v KOH solution and staining with trypan blue (Phillips and Hayman, 1970). Root length was measured by gridline intersect method (Tennant, 1975). Total extraradical mycorrhizal hyphae were determined by the method described by Borie et al. (2000) and the metabolically active hyphae by detecting their dehydrogenase activity (Kabir et al., 1997). Total density of both types of hyphae was quantified by the Newman (1966) line intersect method after collection in a nitrocellulose filter with 0.45 mm pore size (White Gridded HAWG, Millipore, Bedford,Massachusetts, USA). The AMF spores were separated from the soil by wet sieving and decanting in a 70% w/v sucrose solution (Gerdemann and Nicholson, 1963), and were quantified with a stereo magnifying glass with a 30-50 magnification. Soil pH was measured in a soil:dH2O mix (2:5 w/v). Statistical analysis The effect of soil alteration, N source, and their interaction were determined by means of a two-way ANOVA. Means were compared by the orthogonal contrast test. Data not meeting assumption for ANOVA were transformed, but the results are shown in their original measurement scale. Principal component analysis (PCA) was carried out to establish the relationship between distinct response variables, the extracted principal components (PC), and the established treatments. A non-hierarchical cluster analysis with the farthest-neighbor method based on the factors obtained was used to determine the similarity between the different experimental units. In all cases the SPSS software v. 10.0 (SPSS Inc., Chicago, Illinois, USA) was used. Statistical significance was determined at P < 0.05. RESULTS AND DISCUSSION Both soil alteration and N source separately showed a strong influence on most of the studied variables (Table 3). Nevertheless, the interaction of these factors only influenced some variables (pH, root biomass, and AMF spore density). In particular, soil alteration showed a greater influence on changes on AMF propagules, especially root colonization and hyphae density (total and active), as well as on the root biomass production, the shoot/root ratio (S/R), and root length. On the other hand, the N source showed a strong influence on the variation of soil pH which reached a difference of 0.8 units between CT + NH4+ and NT + NO3- treatments (Figure 1). The use of NO3- as N source generated a pH increase in this study, which would be due to the extrusion of OH- to the rhyzosphere associated with the absorption of NO3- to maintain the radical electroneutrality. On the other hand, in treatments where NH4+ was used, lower pH values were observed which would be produced by H+ extrusion in the rhyzosphere (Marschner, 1995; Logan et al., 1999). In addition, the predictable rhizosphere acidification due to nitrification of NH4+, that can represent an important source of acidity in volcanic soils, must be consider (Campillo and Rodríguez, 1984; Martens, 2001; Cornejo et al., 2007). This aspect is especially relevant when choising the type of N fertilizer to be used, particularly in soils that naturally tend to be acidic. Moreover, NT use also produced a pH increase, observing an additive effect of both factors (N source and soil alteration) in the NT + NO3- combination. Observed alkalinization with NO3-, though significantly different in CT treatments, was less than that observed in NT treatments. In particular, the ratio and absolute length of colonized root were 2 to 4 times higher in the NT treatments than in the CT (Figure 2), suggesting a strong influence of established arbuscular mycorrhiza in the rhizospheric pH modification. In addition, pH also increased with NO3- as N source in disturbed and undisturbed soils, which could be due to the greater AMF spore density at the end of the preceding wheat crop in treatments with NO3- as N-source (Table 2). These would have functioned as an increased source of mycorrhizal inoculum, allowing an earlier and more intense colonization in the oat plants fertilized with NO3- as N source. This fact can be especially relevant in annual rotation systems like those commonly used in extensive areas of the Central-Southern Chile, since the N source, besides directly influencing the number of propagules remaining in the soil at the end of the crop, would determine inoculum potential of AMF present in the soil for the next crop in the rotation (Cornejo et al., 2007; 2008b). In a similar study by de Varennes and Goss (2007), it was concluded that the greatest efficiency of tripartite symbiosis among native AMF, rhizobia, and legumes was due to the greater AMF colonization rates which were reached more prematurely in undisturbed than in disturbed soils. Regarding the aforementioned, numerous studies suggest that conventional tillage, disturbing the soil, generates a decrease in the infectiveness of the AMF mycelium by destroying the hyphae network in the soil (see Kabir, 2005 for a review), which supposedly occurred in this study when simulating conventional tillage by sieving. The use of NO3- as N source would generate adequate conditions for greater growth of the fungal structures, especially in soils with acidic characteristics. In this sense, Cornejo et al. (2007) studied the effect of NO3- and NH4+ on the development of Glomus etunicatum in a wheat crop growing in an Andisol under greenhouse conditions, observing that the use of NO3- sequentially promoted greater root colonization in vegetative growth stages of the crop, greater density of metabolically active hyphae at harvest time, and greater density of fungal spores 3 months after harvest. In this study, during the growth stage of oat Zadocks 60, similar results to those obtained by Cornejo et al. (2007) were observed in the same growth stage for wheat, both in mycorrhizal colonization and production of extraradical hyphae (Figure 3). In turn, results obtained in this study suggest that variations in pH and mycorrhization were produced by the addition and/or feedback of the effects of a greater AM functionality (or at least a greater colonization intensity), and the effect of NO3- as N source in the rhizospheric alkalinization by absorption both by the root and fungal hyphae (Bago et al., 1996; Bago and Azcón-Aguilar, 1997). Therefore, this effect would be dependent on both the N source applied, the mycorrhizal status of the plant, and the magnitude of its mycorrhizal colonization (Vaast and Zasoski 1992; Bago et al., 1996; Bago and Azcón-Aguilar, 1997). Density of AMF extraradical hyphae was greater in NT than in CT treatments, particularly the active hyphae density, although this difference was not of the magnitude observed in mycorrhization or mycorrhizal root length (Figure 3). These results agree with those found by Borie et al. (2006) who studied the effect of distinct tillage systems in Ultisols in the Central-Southern Chile, finding greater densities of AMF mycelium and radical colonization in crops cultivated with conservation systems (no-tillage and reduced tillage) than those cultivated in a conventional way. On the other hand, the use of NH4+ as N source was associated with a greater density of total AMF hyphae than with NO3-, suggesting the presence of a compensation mechanism by AMF to capture this ion that is less mobile in the soil, but for which high-affinity transporters have been described in this type of fungi (López-Pedrosa et al., 2006). In the particular case of the NT + NO3- treatment the greater density of active hyphae would be related to a greater level of radical colonization, a preliminary step before extraradical production of hyphae. Furthermore, disturbing the soil would reduce the ratio of metabolically active hyphae in the soil (Figure 3). This is particularly evident in the CT + NH4+ treatment respect to the NT treatments that jointly showed 10% more activity. Relative to the AMF spore production, the NT + NO3- treatment showed a density 62 to 99% greater than the other treatments (Figure 4). This result shows that both NO3- as N source and the tillage systems that disturb less the soil can promote the density of an important biotic component in view of maintaining a long-term presence of AMF propagules. This spore density increase is related to pH increase, situation already observed in previous studies under similar conditions (Rubio et al., 2003; Cornejo et al., 2007; 2008b). In this study, differences in spore density were also found in intermediate growth stages (Z60) which differs from the observed by Cornejo et al. (2007), where these differences, with contrasting N sources, were found in wheat postharvest. This corroborates the manifestation of an additive effect between NO3- as N source and NT on the density of AMF propagules, since the fungus growth and symbiotic development would be accelerated, and be expressed in earlier stages of crops growing in acidic soils. Other observations by the authors of this study have shown direct and significant relationships between plant biomass production and spore density in the soil at postharvest (Cornejo et al., 2008b). Nevertheless, the biomass levels obtained in this study were similar (Figure 5), so the increase of spore density in the NT + NO3- treatment would basically be due to the improvement of the soil characteristics with NO3-, and the greater density of functional AMF propagules. Shoot biomass production did not show any differences except in the CT + NH4+ treatment when compared to the CT + NO3- treatment. Differences of a greater magnitude were observed in root biomass production, showing CT treatments greater values than NT treatments. Compared to the root length (Figure 2), it can be concluded that this greater radical biomass production was associated with root swelling in CT treatment, probably related with the lower observed colonization. These results generate a greater S:R ratio in NT treatments, showing greater biomass production efficiency in the shoot of these treatments, precisely those that showed more colonization and greater active hyphae density. Related to these results, previous studies show that mycorrhized plants demonstrate inhibition of radical growth due to the AMF hyphae, as a whole, acts as a complementary radical system compensating many root functions (Barea, 2000). Principal components analysis, followed by a non-hierarchical cluster analysis allowed the formation of highly homogeneous groups (Figure 6). The experimental design was strongly adjusted to this type of analysis, explaining PC 1 a 57.6% and PC 2 a 42.4% of the total variance, respectively. Principal component 1 was significantly correlated with pH, total and colonized root length, and biomass production, while PC 2 was correlated with the density of total and active hyphae. Among the groups formed after the non-hierarchical cluster analysis, highlight the NT + NO3- treatment with a high incidence in PC 1 and intermediate values in PC 2, representing a group with high biomass production, high mycorrhizal propagule density, and a considerable pH increase. Treatments which used NH4+ as N source formed another group that showed a greater hyphal density associated with a high incidence in PC 2, but heterogeneous with respect to the other studied variables. The CT + NO3- treatment showed intermediate values on PC 1 associated with an intermediate biomass production and AMF propagule densities that would represent, in this particular case, the effect of NO3-, but without the added beneficial effect of NT. It is worth noting that variations in the values of the different studied variables, compared to their initial values (after wheat postharvest, Table 1), show the additive effect of NO3- used as N source and undisturbed soil, especially emphasized in the pH, that registered an increase of 0.46 units in the NT + NO3- treatment (Table 4). This suggests a positive interaction relative to the use of ammonium N sources and conventional tillage systems that could be evident in rotation programs in the short- or medium-term. In the case of AMF hyphae and spores, virtually in all the analyzed situations, a decrease was observed, probably due to the early use of these propagules in the first colonization stages (spore germination and radical colonization), before the stages of structure resistance formation, temporarily close to plant senescence (Cornejo et al., 2007). In spite of the aforementioned, it can be observed that the decrease related to the initial values was more moderate in NT than in CT treatments, especially in the cases in which NO3- was used. This greater propagule persistence and density is of special agroecological interest when the available alternatives to manage AMF populations naturally found in volcanic soils in Southern Chile are considered. CONCLUSIONS It can be concluded that the N source used as fertilizer and the application of practices which disturbing or non-disturbing the soil in crop successions of mycorrhizable species, such as those commonly established in volcanic soils in Southern Chile, have a strong effect on the persistence and density of AMF propagules, and pH changes in the soil. In particular, the use of NO3- is associated to the widely studied effect of rhizospheric alkalinization, as well as to a greater density of AMF propagules that would facilitate early mycorrhizal colonization in the plant species of the next crop in the succession. On the other hand, conservation tillage systems, such as the one simulated in this study, generate a considerable increase in mycorrhizal colonization and AMF propagule density, as well as increased pH in the soil, with an additional effect to the one observed with NO3- as N source. These aspects can be of a special agronomic relevance since fertilization and tillage system represent two aspects of crop management with the greatest impact on the productivity and stability of agroecosystems. ACKNOWLEDGEMENTS This study was financed by FONDECYT Project 1990756, from Comisión Nacional de Investigación Científica y Tecnológica, CONICYT, Chile. LITERATURE CITED
Copyright 2009 - Chilean Journal of Agricultural Research The following images related to this document are available:Photo images[cj09051t1.jpg] [cj09051t2.jpg] [cj09051f6.jpg] [cj09051t4.jpg] [cj09051f4.jpg] [cj09051f1.jpg] [cj09051f2.jpg] [cj09051f5.jpg] [cj09051t3.jpg] [cj09051f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}