 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Chilean Journal of Agricultural Research, Vol. 69, No. 4, Oct-Dec, 2009, pp. 477-485 Research Importance of ionic and osmotic components of salt stress on the germination of four quinua (Chenopodium quinoa Willd.) selections Importancia de los componentes iónico y osmótico del estrés salino sobre la germinación de cuatro selecciones de quinua (Chenopodium quinoa Willd.). José Delatorre-Herrera[1]*, and Manuel Pinto[2] [1]Universidad Arturo Prat, Departamento de Agricultura del Desierto y
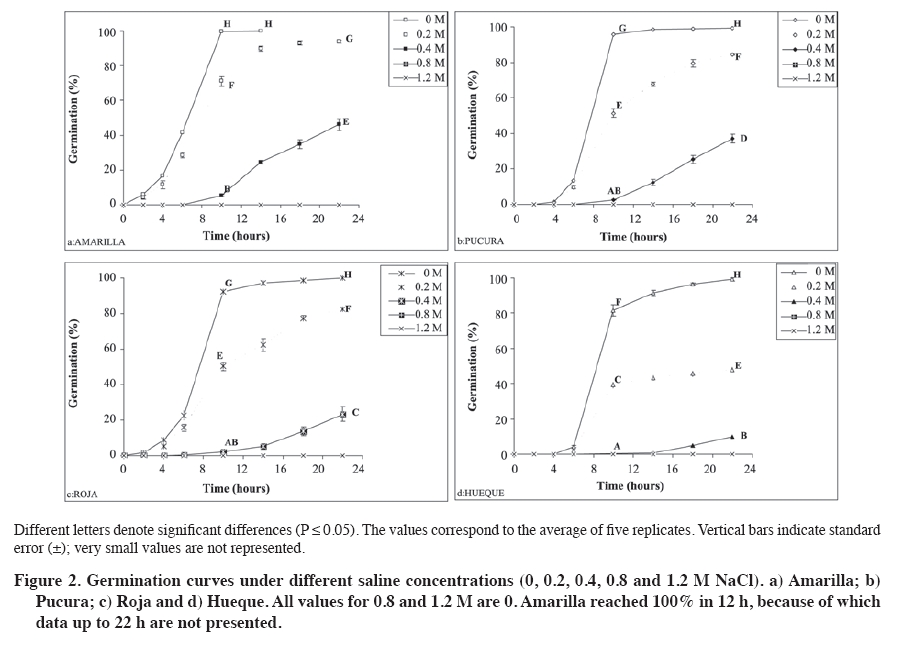
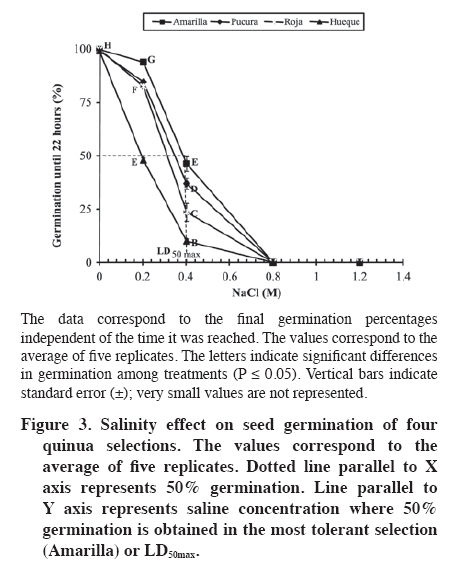
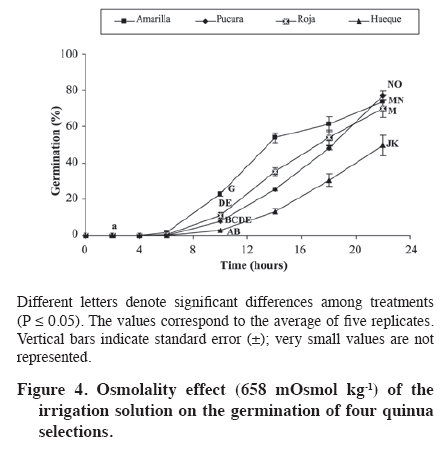
Biotecnología, Casilla 121, Iquique, Chile. *Corresponding author (jose.delatorre@unap.cl) Received: 05 May 2008. Code Number: cj09057 ABSTRACT The influence of the components of saline stress (osmotic and ionic factors) were evaluated during the germination of four quinua (Chenopodium quinoa Willd.) selections in Chile, two collected in the arid northern highland region with saline soils (Amarilla and Roja) and the other two from the rainy southern region with non-saline soils (Hueque and Pucura). The seeds were treated with different saline concentrations (0, 0.2, 0.4, 0.8 and 1.2 M NaCl). Germination kinetics shows that the control treatments of the four selections reach 100% germination. With application of 0.4 M NaCl (LD50max) germination was reduced by 53% in the Amarilla selection and 89.9% in Hueque. The germination rate was lower for the seeds from non-saline areas, which reached germination after 22 h, compared to 10 h for Amarilla. By separating the salinity stress factors (osmotic and ionic), it is evident that they have different degrees of influence on quinua germination, according to each selection. Thus, the germination of Amarilla selection from the arid area was the least influenced by the ionic factor (27%), Pucura and Roja were more affected by the ionic factor (40% and 46% respectively), and Hueque, from the rainy area, was more affected by the osmotic factor, in which caused a 50% loss in germination. Keywords: Chenopodium, salinity, imbibition, seeds, NaCl, altiplano, desert. RESUMEN La influencia de los componentes del estrés salino (factores osmótico y iónico) fue evaluada durante la germinación en cuatro selecciones de quinua (Chenopodium quinoa Willd.), dos cultivadas en la zona árida y salina del altiplano (Amarilla y Roja) y dos en la zona húmeda y suelos no salinos en Chile (Hueque y Pucura). Las semillas fueron tratadas con diferentes concentraciones salinas (NaCl 0; 0,2; 0,4; 0,8 y 1,2 M). Las cinéticas de germinación muestran que los tratamientos testigos de las cuatro selecciones alcanzan un 100% de germinación. Al aplicar NaCl 0,4 M (LD50max), la germinación se reduce en un 53% en la línea Amarilla y 89,9% en Hueque. La velocidad de germinación fue menor en las semillas provenientes de selecciones de zonas no salinas, las que demoran 22 h en alcanzar el 100% de germinación, en comparación con las 10 h de Amarilla. Al separar los factores estresantes de la salinidad (osmóticos y iónicos) es posible concluir que éstos tienen diferentes magnitudes de influencia en la germinación de quinua, según cada selección. Así la germinación en la selección Amarilla de la zona árida fue la menos influenciada por el factor iónico (27%), Pucura y Roja fueron más afectadas por el factor iónico (40% y 46%, respectivamente), en tanto que el factor osmótico influyó en un 50% en la pérdida de la germinación. en Hueque. Palabras clave: Chenopodium, salinidad, imbibición, semillas, NaCl, altiplano, desierto. INTRODUCTION Glycophyte plants have low levels of tolerance to saline stress. For example, an increase in salinity from 0 to 180 mM of NaCl decreases germination by 50% in species of the genus Phaseolus (Bayuelos et al., 2002). In contrast, halophyte plants are more tolerant to salinity. For example, Cakile maritima Scop. can support a concentration of salts close to 400 mM of NaCl without showing effects in 80% of its germination. High concentrations of NaCl induce dormancy in the seeds of many halophyte species (Gul and Weber, 1998; Debez et al., 2004), while seeds of glycophyte species loss their viability under similar conditions. Nevertheless, germination in both halophyte and glyophyte plants is sensitive to salinity (Pujol et al., 2000). The velocity of germination is one aspect that is affected. For example, in C. maritime, germination begins 4 days later with treatments of 400 mM than with treatments of lower concentrations (Debez et al., 2004). Several authors have shown that the decrease in seed germination caused by salinity is the result of the joint action of two types of stress factors: water deficit produced by the osmotic effect of salts in the soil solution, also termed “osmotic dehydration”, and toxicity as a consequence of the excessive intake of ions such as Cl- and Na+ in tissues (Munns et al., 1995; Zhu, 2003). During germination, the joint action of the two stresses is manifested as a decrease in the percentage and velocity of germination (Huang and Redmann, 1995; Ungar, 1996; Kerepesi and Galiba, 2000). In halophyte plants the decrease in germination is mainly the consequence of lower osmotic potentials in the soil solution, given that ion intake by seeds can be limited or harmless, while in glycophytes, the excessive accumulation of ions in tissue causes nutritional imbalance, toxicity and finally death (Munns et al., 1995; Katambe et al., 1998; Dodd and Donovan, 1999). Thus, the differences in germination observed between the seeds of tolerant and sensitive plants could be the results of the activation of different mechanisms (Dodd and Donovan, 1999; Kerepesi and Galiba, 2000; Almansouri et al., 2001) of cells from the different parts of the embryonic axis in order to tolerate both osmotic and ionic stress during germination. The relative importance of the two stresses has not been well studied in species such as quinua, which is characterized by its adaptability to different edaphic conditions and tolerance to salinity (Jacobsen et al., 1999; Mujica et al., 2001). This species has been cultivated for centuries in the desert of northern Chile in soils with high salinity, with electrical conductivity (EC) that can reach values close to 10 dS m-1 (Delatorre et al., 1995). However, using a cultivar from the central-southern region of Chile, and therefore possibly adapted to soils with low EC, Karyotis et al. (2003) observed that an ED higher than 6.5 dS m-1 reduces germination in these accessions by 70%. The contrast in the response to salinity between selections from northern and southern Chile is probably the consequence of adaptations achieved over time by populations that developed in isolation from one another under very different environmental conditions. This has induced some degree of genetic distancing among these populations, segregating them into two discrete groupings. This segregation has broad agricultural implications, in particular in the adaptability that they have developed to their respective environments (Fuentes et al., 2005; 2008). Consequently, it is possible that the quinua populations from the north are closer to halophytes or facultative halophytes, as described by Bosque et al. (2003), while the selections from the south could be closer to glycophyte plants. In consideration of this background information, it can be argued that during quinua germination there are differences in levels of susceptibility to salinity between selections acclimatized to the arid and saline conditions of the altiplano and selections adapted to the conditions of high precipitation and non-saline soils of southern Chile. These differences could be expressed in the germination percentages and rates and the relative importance of the two components involved in salinity (osmotic and ionic factors). The main objective of the present work was to establish the effect of saline stress on germination, as well as the relative importance of the component factors of this stress, ionic and osmotic stress, on four selections of quinua from two contrasting agro-ecological conditions, namely the northern high Andes and salt flats on the one hand, and the humid southern Chilean region with non-saline soils found at sea level. MATERIALS AND METHODS Seeds from four selections of quinua: Amarilla and Roja from the altiplano region of northern Chile (19º28´ S, 68º50´ W), and Hueque and Pucura from Temuco (38º43´ S, 72º32´ W) in southern Chile, were germinated on Whatman 42 filter paper in Petri dishes of 10 cm in diameter, embedded in 10 mL NaCl solutions with the following concentrations: 0; 0.2; 0.4; 0.8 and 1.2 M. There were five replicates of each treatment, each of which corresponded to a Petri dish with 100 seeds. The seeds were incubated for 22 h at 25 ºC in darkness in a germination chamber (Incubator 865, Precision Scientific, Chenai, India). Responses were measured in terms of the percentage of germination, which was considered as the presence of radicula of more than 0.5 mm. NaCl (Hach Company, Loveland, Colorado, USA) was used for preparation of the saline solutions at 99.9% of purity with de-ionized distilled water. Seed counts were made at 2, 4, 6, 10, 14, 18 and 22 h after seeding. The germination kinetics and germination velocity for each selection were obtained from this information, in accordance with the method of Khan and Ungar (1984). Based on the final germination values, the saline concentration was determined that reduced germination in each selection by 50%. This value is termed LD50. This range was established considering the requirement that the genotypes used express the effects of germination on salts at non-lethal concentrations. To separate the ionic and osmotic effects, the concentration was used that reduced germination by 50% in the most tolerant selection, termed LD50max, with the aim that this selection could express both the morphological and biochemical effects of salts. To determine the Na content, all the germinated seeds in each replicate were collected, dried at 70 ºC for 48 h and then milled. Na was measured according to the procedure described by Hunt (1982). A sample of 0.1 g was digested in 15 mL of HCl 0.5 M for 2 days. Na concentration was determined with a flame photometer (PFP7 Jenway, Gransmore, England). The osmotic potential (Ψs) of the seeds was determined by macerating 0.1 g of dry matter (DM) obtained from the complete seed previously congealed liquid N in 1 mL of nanopure water. The extract was centrifuged and an aliquot of supernatant was injected into an osmometer (3000 model, Advance, Norwood, Massachusetts, USA). The calculation of Ψs was obtained through the Van’t Hoff equation [1]: Ψs = C * T * R [1] where C corresponds to the concentration or osmolality (mOsmol kg-1 H2O) determined with an osmometer, T is the absolute temperature and R is the gas constant (0.00831 MPa mol-1 ºK-1). Determination of the ionic and osmotic effects According to Munns et al. (1995), the decrease in germination under saline conditions is the consequence of the combined effect of osmotic and ionic factors; consequently the total effect (TE) of salinity on germination can be defined as: TE = OE + IE [2] where it is deduced that the osmotic (OE) or ionic effects (IE) corresponds in turn to the subtraction of the complement corresponding to each one in relation to the total effect (TE). To resolve this equation, the osmotic components (EO) were separated, which were determined by germinating seeds in distilled water (zero osmolality) and in an isotonic solution of polietilenglycol (PEG 8000) with an osmolality equivalent to that of 0.4 M NaCl (LD50max) or 658 mOsmol kg-1. Consequently, OE corresponds to the difference between the germination values obtained under pure water conditions (GH2O) and those obtained in the isotonic solution (GOs) OE = GH2O - GOs [3] An osmometer was used according to the procedure already described in order to determine the osmolality of each saline solution. The total effect (TE) of salinity, or the combined osmotic and ionic effect, is obtained by means of the difference between the germination values under non-saline conditions with nanopure water (GH2O) and germination obtained with the saline concentration LD50max. This germination is termed GLD50max, thus TE was determined through: TE = GH2O - GLD50max [4] Based on the values of TE and OE, the ionic effect (IE) was calculated: IE = TE – OE [5] Statistical analysis A factorial design in completely random blocks (Bi) was used in this study, with two factors: selections (Sk) (Amarilla, Roja, Pucura and Hueque) and salinity (Cj) (NaCl 0; 0.2; 0.4; 0.8 and 1.2 M). All the data were analyzed statistically used the variance test (ANOVA) with the INFOSTAT program (2004) 2007p version. The Duncan’s test was used for multiple comparisons, with a 95% level of significance. The percentages were transformed into an arcsine through the equation described by Dughetti and García (2004). The correlations were determined through a principle component analysis. RESULTS AND DISCUSSION Germination under non-saline conditions Under the non-saline conditions, all the selections present a sigmoid germination curve, with a first stage characterized by a low germination rate, which lasted until 6 h after seeding; an intermediate stage between 6 to 14 h with a maximum germination rate, and a final stage of an asintotic character in which the germination velocity increases minimally (Figure 1). All the selections reached 100% of germination, although at different times, which shows the good quality of the seeds used. The seeds of the selection Amarilla were the most rapid to germinate, presenting in the first stage a rate of 6.9 germinated seeds per hour (seed h-1), reaching 41.6% germination at 6 h after seeding. This germination velocity is in contrast to that presented by the selection Hueque. The latter presented only 0.7 seed h-1, reaching close to 3% of germinated seeds at the end of the same time (Figure 1). In this stage the selections Roja and Pucura presented intermediate values of 3.7 and 2.2 seed h-1, respectively. In the second stage of germination kinetics (between 6 and 14 h), the differentials among the germination rates observed in the first stage tended to be lower. However, owing to the marked differences presented in the first stage, the percentages of germination between Amarilla and Hueque were significantly different (P ≤ 0.05) at the end of the second stage (14 h). In effect, at the end of this stage the selection Amarilla reached 100% germination, while Hueque reached only 80%. Research on seeds of cv. Sajama from the Bolivian salt flats, which were irrigated with distilled water, showed that this variety reached the maximum germination at 10 h (Prado et al., 2000), very similar results to those described by Jacobsen and Bach (1998), who worked under non-saline conditions with the var. Olav originating from southern Chile. Because 100% of Amarilla seeds germinated by 10 h, the final phase of this selection was very short, while in Hueque it was prolonged to 22 h before reaching 100%. For their part, the selections Roja and Pucura presented an intermediate behavior, reaching 100% of germination by 18 h (Figure 1). As the concentration of salts in the saline solution increased (Figure 2), the percentages of germination decreased significantly in all the selections (P ≤ 0.05). Comparing the final germination percentages (hour 22) among the seeds irrigated with saline solutions and the control treatments (NaCl 0 M), it can be observed that with 0.2 M NaCl, the most affected selection was Hueque, which decreased germination by 51.2%, while the decrease in Amarilla was only 6%. With 0.4 M NaCl, germination was more drastically reduced in all the selections (P ≤ 0.05), Amarilla decreased by 53.8%, Pucura by 63%, Roja by 76.6% and Hueque, which was the most affected, by 90.2%. Concentrations of NaCl above 0.8 M inhibited germination in all the selections. Salinity inhibited germination in the four selections of evaluated quinua. The selection Amarilla was the most tolerant, given that its germination was reduced by 53.8% in solutions of 0.4 M of NaCl (LD50max), unlike Hueque, in which germination decreased by 89.4% (Figure 2). This reaffirms that of the four evaluated selections, the most tolerant to salinity is Amarilla from northern Chile and Andean salt flats. The most sensitive to salinity is Hueque, from the rainy non-saline region of southern Chile. Based on the final germination data and considering 22 h after seeding as a limit, a period in which Hueque reached 100% germination in the control treatment, LD50 was estimated for each selection (Figure 3). According to these data, Hueque seeds decreased germination by 50% when 0.2 M NaCl was applied. For its part, Amarilla did the same when 0.4 M NaCl was applied. Germination in Pucura and Roja decreased by 50% with intermediate saline concentrations of 0.36 and 0.32 M NaCl, respectively. These results confirm that the selection Hueque was the most sensitive to the total effect of salts (osmotic and ionic), with a low tolerance to salinity (LD50 = NaCl 0,2 M). For the accession that was most tolerant to salts, Amarilla, LD50 was at 0.4 M NaCl and consequently this salt concentration was used as LD50max. This concentration has been used empirically by other authors to induce saline stress in quinua (Prado et al., 2000), so that these results confirm this doses as a sub-lethal concentration that significantly affects germination of quinua seeds. In this respect, all the selections submitted to LD50max (NaCl 0,4 M) showed a considerable delay in germination. The selection Amarilla, for example, did not exceed 3.3 seeds h-1, while Hueque reached rates of only 0.7 seeds h-1. Separation of the component stressful factors of salinity Determination of the osmotic effect (EO) on the germination of quinua seeds. The data on germination using an iso osmotic solution equivalent to 0.4 M NaCl (658 mOsmol kg-1 or -1.4 MPa) is presented in Figure 4. These results show that the selections Pucura, Amarilla and Roja presented very similar final germination percentages: 77.0; 73.8 and 69.8%, respectively. On the other hand, the selection Hueque was again the most affected by the osmotic potential of the solution (-1.4 MPa), in that its germination was reduced by 49.6%, which differed significantly from the other selections (P ≤ 0.05). However, this data alone are not sufficient to determine if OE is the preponderant effect in the deterioration of germination of this selection. These results also show that all the selections under conditions of osmotic stress present a delay of 6 h in the velocity of germination compared to the control treatment. Nevertheless, this process advance germination by 4 h compared to that with 0.4 M NaCl (Figure 2), which shows the ionic effect as the more influential as the osmotic in deteriorating germination. This is supported by data from Almansouri et al. (2001), who reported that the application in wheat of an iso-osmotic solution (1.57 MPa) of polietilenglicol (PEG) had a greater effect on germination than NaCl as a result of the reduced osmotic potential of the irrigation solution. Influence of ionic effect on quinua seeds subjected to saline stress: To separate the osmotic and ionic effects, the percentages of germination obtained with nanopure water were used, with NaCl 0.4 M (LD50max) and 658 mOsmol kg-1 of the equivalent iso osmotic solution (-1.4 MPa). Based on these data, it is possible to determine that osmotic potential significantly influences (P ≤ 0.05) the selection Hueque, provoking a 50% reduction in germination (Table 1). This could indicate that Hueque is a susceptible species to water deficit and that the osmotic effect in salinity stress has a major influence on its germination. Nevertheless, the ionic effect is not less important, given that it represents 40% of the decrease in germination of this selection. For their part, germination in Amarilla, Pucura and Roja is more affected by the ionic potential, with reductions in germination of between 51.3 and 64.5%. Table 1. Determination of the influence of osmotic and ionic factors of saline stress on seed germination in four quinua selections submitted to 0.4 M of NaCl.
Different letters denote significant differences among treatments (P ≤ 0.05). The values correspond to the averages of five replicates. Notably, these results confirm that Hueque is susceptible to salinity, with both factors (osmotic and ionic) being equally important. On the other hand, the same analysis determines that Pucura and Roja are more influenced by the ionic factor (40% and 46%, respectively). Amarilla was shown to be the most tolerant given the low influence of the osmotic (26.2%) and ionic factors (27.6%) on germination. The osmotic effect reduced germination in Hueque by 50%, while the ionic effect reduced it by 40%. This indicates that in this case germination is affected by the availability of water than by the ionic effect. The glycophyte character of this selection might be a consequence of its acclimatization to non-saline environments with high precipitation. Calculating the relative importance of the two stresses (osmotic and ionic), it is possible to determine that the two stressful factors have a proportional participation in the deterioration of germination in Amarilla and Hueque that is manifested as a ratio of 1.1 and 0.8, respectively, differentiating them in the magnitude of the effect. For their part, germination of Pucura and Roja are more influenced by the ionic effect. Based on the principle component analysis (PC) of the percentages of germination, we can infer that there is a correlation between ionic (EI) and osmotic effects (OE) in each selection. As there are only two variables (IE and OE) and the PCs are lineal combinations of these variables, it can be expected that PC explains 100% of the variation caused by the two effects. Under this statistical procedure, the principle component 1 (PC1) explains 59.6% of the behavior and allows for determining that the selection Hueque is the most affected by saline stress of all the selections. Pucura and Roja presented an intermediate effect, while Amarilla was the most tolerant to salts. Nevertheless, the principle component 2 (PC2), which explains 40% of total variability, allows for differentiating the most susceptible and tolerant varieties in relation to the two stress factors. PC2 indicates that Hueque correlates positively with osmotic stress, while Amarilla is less influenced by ionic stress (IE). Quinua germination was affected by both OE and IE independent of the accessions, which is in agreement with what was indicated by Poljakoff-Mayber et al. (1994). The analysis by osmotic and ionic factors (Table 1) shows that OE causes a 22 to 30.2% decrease in germination of in Amarilla, Pucura and Roja. This indicates that these selections can behave as pseudo-halophytes, consistent with what was found by Bosque et al. (2003). OE causes a decrease in water availability, affecting water intake and consequently germination, which is why the ionic phase is not manifested. This occurs in seeds of glycophyte plants that are affected by salts, such as wheat (Almansouri et al., 2001) and is similar to the situation of the accession Hueque, which comes form a non-saline region. Katambe et al. (1998) indicate that high concentrations of NaCl applied to seeds of halophyte plants, such as Atriplex prostrata, are more inhibiting of germination than iso-osmotic applications of PEG. In this case, osmotic and ionic effects act together. Nevertheless, in this last point there are discrepancies, given that Song et al. (2005) indicates that both Na+ and Cl- contribute to reducing the inhibiting effect of the osmotic potential of seeds of halophyte plants, raising the percentage of germination under saline conditions, as occurs in the selections Amarilla, Roja and Pucura. Khan and Ungar (1996) indicate, as well that the seeds of halophyte plants that do not germinate often enter dormancy. In this respect, Table 3 shows the percentage of seeds that germinate after having been submitted to different saline treatments, ratifying that Amarilla is more tolerant to salts, given that it recovered 23.1% of seeds that did not germinate owing to saline stress. Table 3 also shows that quinua seeds entered saline dormancy. Analyzing the Na content, the main cause of the ionic effect is made evident by the ratio: final/initial Na concentrations (Table 2), in which the selections Amarilla and Roja, from the slat flats of the high Andes, show an increase in this ratio when they are submitted to LD50max (0.4 M NaCl). Table 2. Ratio between sodium absorbed at 22 h vs. initial sodium content in seeds germinated with 0 and 0.4 M NaCl
T: measurements at 0 and 22 h. Different letters denote significant differences among treatments (P ≤ 0.05). The values correspond to averages of five replicates. Table 3. Recovery of seed germination after salt stress
* Mean ± standard deviation. Lower case letters indicate significant differences according to Duncan test (P ≤ 0.05). Thus, both Amarilla and Roja increased Na intake toward the end of the germination period (22 h) by 8.9 and 10.5 times, respectively, compared to its initial concentration. Nevertheless, Roja is more sensitive to salts than Amarilla, which is in agreement with what was indicated by Cramer et al. (1994), who found that higher intake of Na+ in corn (Zea mays L.) does not correlate with salt tolerance, a situation that is observed with all the evaluated accessions. Management of Na in the tissues of tolerant species is controlled by the existence of mechanisms that allow for effective handling salts, such as the mobilization of ions to maintain ionic and osmotic homeostasis in the cells (Flowers et al., 1977; Hasegawa et al., 2000; Blumwald, 2000; Zhu, 2003). CONCLUSIONS Salinity affects the germination process in quinua seeds. However, the evaluated selections have different levels of tolerance to salts (NaCl). The selection Hueque is considered the most sensitive, presenting a 50% seed loss (LD50) with 0.2 M NaCl, while Amarilla was the most tolerant, with an LD50 of 0.4 M NaCl. The components of salinity (osmotic and ionic effects) affect the evaluated quinua selections differently. Hueque was affected more proportionally by the osmotic effect, while Pucura and Roja are more affected by the ionic effect. The selection Amarilla, which was considered as the most tolerant, presented a low influence of both factors, in particular of the ionic effect. ACKNOWLEDGEMENTS The authors are grateful for the support provided by the Fundación para la Innovación Agraria (FIA) through the Project FIA-PI-C-2004-1-A-079, by FONDECYT through the Project Nº 1060281, as well as the Department of Desert Agriculture and Biotechnology of the Universidad Arturo Prat, with special thanks to Alejandra Acevedo of the Physiology and Vegetal Nutrition Laboratory. LITERATURE CITED
Copyright 2009 - Chilean Journal of Agricultural Research The following images related to this document are available:Photo images[cj09057f4.jpg] [cj09057f1.jpg] [cj09057f2.jpg] [cj09057f3.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}