 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Chilean Journal of Agricultural Research, Vol. 69, No. 4, Oct-Dec, 2009, pp. 541-547 Evaluation of Metarhizium anisopliae var. Anisopliae Qu-M845 isolate to control Varroa destructor (Acari: Varroidae) in laboratory and field trials Evaluación del aislamiento Qu-M845 de Metarhizium anisopliae var. anisopliae para el control de Varroa destructor (Acari: Varroidae) en ensayos de laboratorio y terreno. Marta Rodríguez[1], Marcos Gerding1*, Andrés France1, and Ricardo Ceballos1 1Instituto de Investigaciones Agropecuarias INIA, Casilla 426, Chillán, Chile. *Corresponding author (mgerding@inia.cl). Received:
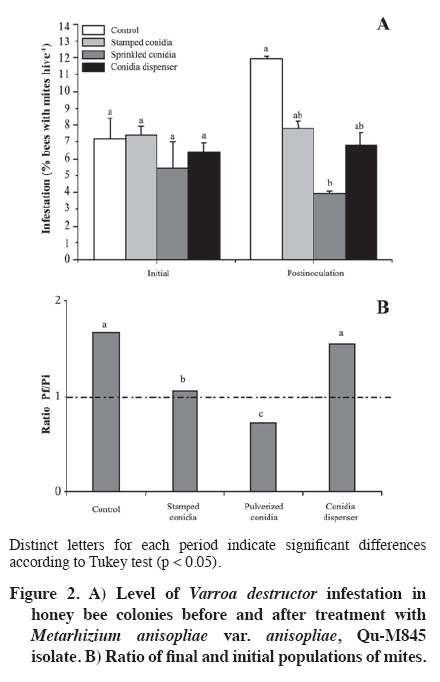
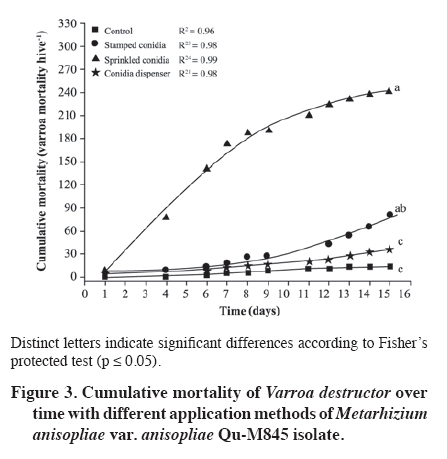
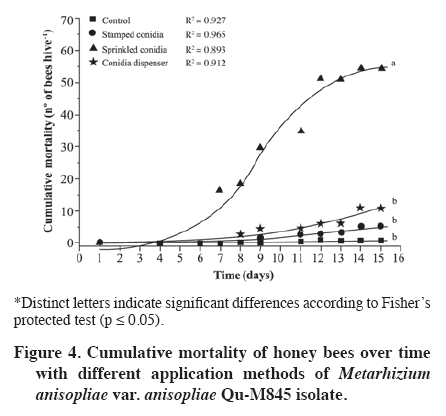
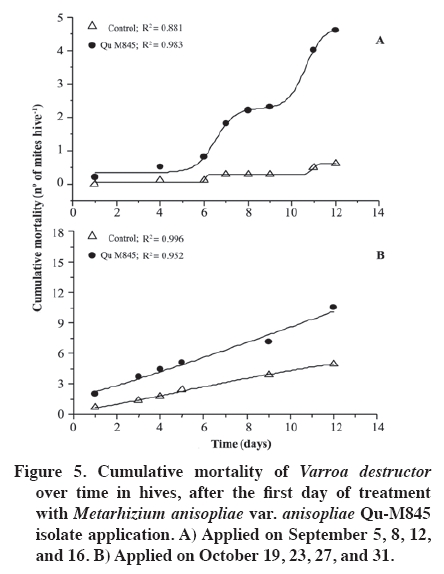
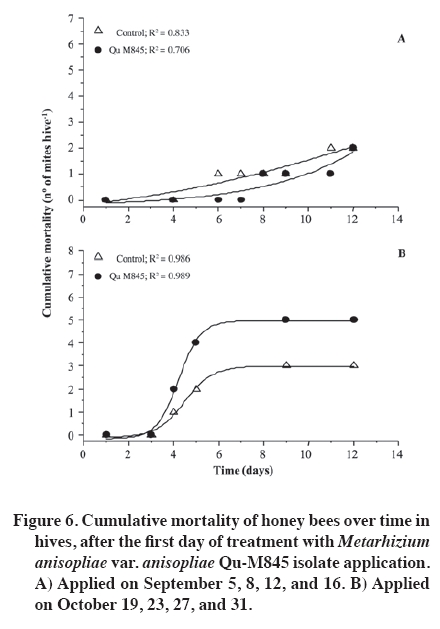
02 November 2007. Code Number: cj09065 ABSTRACT The effectiveness of the Metarhizium anisopliae (Metschinkoff) Qu-M845 isolate was evaluated in laboratory and field trials. It was previously selected for thermal resistance (at 30 and 35 ºC) and pathogenicity on Varroa destructor (Anderson and Trueman). In the laboratory, the first evaluations were carried out by spraying and increasing the concentration from zero to 108 conidia mL-1 on varroa adults. Lethal concentrations required for 50 and 90% mite mortality (LC50 and LC90) were 3.8 x 105 and 8 x 107 conidia mL-1, respectively (χ2 = 2.03). In the autumn field trials, three application methods (doses of 5 x 1010 conidia per hive) were evaluated. The treatments were: a) conidia stamped on filter paper, located on every second frame inside the hive; b) dry conidia sprinkled on and between frames; and c) dry conidia in a dispenser path at the entrance of the hive. Furthermore, untreated hives were included as controls. After 21 days of treatment, the dry conidia sprinkled on and between frames showed 67% less bees infested by the mite than the control (p < 0.05). Average bee mortality observed was significantly different between treatments (p < 0.05). The sprinkled dry conidia caused the highest mortality of bees. In spring, this treatment produced increased mite mortality and without bee mortality. These results demonstrated that it is possible to develop a biological insecticide based on this isolate to generate an alternative to control V. destructor. Keywords: biological control, entomopathogenic fungi, Varroa destructor, varroasis, insect pathology, Metarhizium anisopliae. RESUMEN La efectividad del aislamiento Qu-M845 de Metarhizium anisopliae (Metschinkoff), seleccionado previamente por su resistencia a temperaturas de 30 y 35 ºC, y patogenicidad sobre Varroa destructor Anderson y Trueman fue evaluada en laboratorio y en ensayos de terreno. Las primeras pruebas consistieron en pulverizar concentraciones crecientes de 0 a 108 conidias mL-1 sobre varroas adultas. La concentración letal para matar el 50 y 90% de la población (CL50 y CL90) fueron de 3,8 x 105 y 8 x 107 conidias mL-1, respectivamente (χ2 = 2,03). En otoño se evaluaron en terreno tres métodos de aplicación de una dosis de 5 x 1010 conidias por colmena. Los tratamientos fueron: a) conidias estampadas en papel filtro ubicado cada dos panales móviles al interior de la colmena; b) conidias espolvoreadas sobre y entre los panales; y c) dispensador de conidias ubicado en la piquera de las colmenas. Además se incluyeron colmenas sin tratar (testigo). Después de 21 días de la aplicación, el tratamiento de conidias espolvoreadas sobre y entre los panales presentó una disminución de 67% en el porcentaje de abejas infestadas, en relación al testigo (p < 0,05). Al evaluar la mortalidad de abejas se observaron diferencias entre tratamientos, siendo el espolvoreo de conidias el que causó mayor caída de abejas (p < 0,05). En primavera, este tratamiento produjo un aumento en la caída de ácaros sin registrar mortalidad de abejas. Estos resultados indican que es factible desarrollar un acaricida biológico en base a este aislamiento, generando, con ello, una alternativa de control para V. destructor. Palabras clave: control biológico, hongo entomopatógeno, Varroa destructor, varroasis, Metarhizium anisopliae. INTRODUCTION The Varroa destructor (Acari: Varroidae) mite is originally from Asia where it parasitizes bees Apis cerana Fabr. (Hymenoptera: Apidae). Due to seasonal migration of livestock, this parasite was extended to Apis mellifera Linnaeus, causing serious worldwide damage to the population of this species (Chandler et al., 2001). Since its detection in Chile in 1992, in the Province of San Fernando, Libertador General Bernardo O’Higgins Region, it has spread to almost the whole country causing serious economic losses to beekeepers (Hinojosa and González, 2004; Neira et al., 2004). Varroasis, the disease caused by this mite, starts when the fertilized V. destructor female abandons the adult bee that it has parasitized and penetrates in a bee breeding cell about to be sealed. Mite oviposition starts two days later, the female lays between 3 and 12 eggs, after 48 h the nymphs are born, start to feed on the hemolymph of the forming bee, and are converted into adults within 5 to 8 days. The mating of the mites occurs in the cell before the bee emerges (Harbo and Harris, 1999; Llorente, 2003). The V. destructor female looks for the soft zones of the adult bees to perforate and suck their hemolymph, causing physical damage by decreasing protein content, and infectious toxin due to the transmission of microorganisms causing viral and bacterial diseases (Chandler et al., 2001; Nordström, 2003). Methods to control varroasis are based on the use of synthetic acaricides such as tau-fluvalinate, flumetrin, and coumaphos. However, it is possible to find resistance to these products (Harbo and Harris, 1999) and their residues in honey and wax (Calderone, 1999; Pérez et al., 2000). Another alternative control is the use of oils and organic acids such as formic acid, oxalic acid, and thymol, which have been intensively studied in Europe and Asia. In spite of its effectiveness in controlling varroa, damaging effects have been reported on bees, such as alteration in the recognition of the queen’s oviposition, bee mortality, and caustic effects on the skin of the handler (Calderone, 1999; Kanga et al., 2003; Floris et al., 2004). An alternative control method is the use of biological control agents, a practice which has aroused increasing interest, given the concern to count on new non-contaminating ways of pest control, and because more is known about the biology and controlled production of these entomopathogenic organisms (Tanada and Kaya, 1993; Dorta and Arcas, 1996). Background information on the use of entomopathogenic fungi (EPF) point out that it is possible to control mites in all of their stages, including eggs, with fungi such as Beauveria bassiana (Balsamo) Vuillemin and Metarhizium anisopliae var. anisopliae (Metschinkoff) (Chandler et al., 2000). Field trials carried out in beehives in the U.S. have demonstrated the capacity of M. anisopliae to control V. destructor (Kanga et al., 2003). Preliminary trials carried out by the Centro Tecnológico de Control Biológico of the Instituto de Investigaciones Agropecuarias INIA Quilamapu with entomopathogenic fungi has made it possible to obtain the M. anisopliae Qu-M845 isolate, selected for its thermal tolerance and high pathogenicity on V. destructor (Rodríguez et al., 2009). In accordance with the above-mentioned, the purpose of this study was to evaluate the mortality caused by this isolate on Varroa destructor, determine the lethal concentration (LC) and the lethal time (LT) for the concentration close to CL90 in laboratory conditions, as well as to study different application methods and effectiveness in the field. MATERIALS AND METHODS Laboratory trials Trials were carried out in the laboratories of the Centro Tecnológico de Control Biológico of the Instituto de Investigaciones Agropecuarias INIA, Centro Regional de Investigación Quilamapu and in the apiary of the Campo Experimental Santa Rosa, Chillán, Chile. Isolate multiplication We used a strain of M. anisopliae var. anisopliae Qu-M845 sown on Petri dishes with potato dextrose agar (PDA) and incubated at 25 ºC until sporulation. Conidia were harvested from the surface of the culture carried out in a sterile distilled water suspension plus 0.1%. Tween 80 (Sigma®, Steinheim, Germany). Conidia concentration was determined with a Neubauer counting chamber (BOECO, Germany) (Rodríguez et al., 2006). Lethal concentration test Female V. destructor adults were collected from infested beehives and transferred to plastic chambers with blotting paper at the bottom. Then suspensions were applied: 105, 106, 107, and 108 conidia mL-1, plus Tween 80 on V. destructoradults, with a Potter pulverization tower system. Adults treated with distilled water plus Tween 80 made up the control. Inoculated varroa were immediately placed on drone pupas maintained in 1 mL plastic tubes with cotton wool at the top and incubated between 25 and 30 ºC in the dark (Rodríguez et al., 2009). Varroa mortality with the different inoculum concentrations were recorded daily and the dead mites were incubated in humid chambers at 25 ºC to observe symptoms of mycosis. Microscopic preparations were carried out with the dead bodies to identify the fungus and check if it coincided with the initial isolate. The experimental design was completely random with five treatments and five replicates, each experimental unit consisting of 10 mites. Varroa mortality curves were expressed as a percentage of mortality in distinct concentrations and were adjusted to sigmoid curves from which CL50 and CL90 were calculated. Mortality of distinct concentrations were compared on day 7 post inoculation with analysis of variance and mean separation by the Tukey test (p < 0.05) (Gomez and Gomez, 1984). TL50 was calculated from the mortality observations accumulated over time with CL90. Field trials Evaluation of application methods An apiary, with 25 Langstroth-type hives of A. mellifera, was established in the Campo Experimental Santa Rosa (36°31' S, 71°54' W) of INIA-Quilamapu in October 2005. In May 2006, doses of 5 x 1010 conidia (10% humidity) were prepared for each hive from the M. anisopliae Qu-M845 isolate obtained from precooked sterile rice. Treatments consisted in three application methods: a) conidia stamped on filter paper located on every second moveable frame inside the hive; b) conidia sprinkled on and between the frames; and c) conidia dispenser path located at the one way entrance of the hives. Furthermore, a control treatment was included that consisted in hives without any treatment. Sampling was carried out before the trial to determine the initial percentage of varroa infestation. The experimental design was completely random with four treatments and three replicates. The randomized experimental unit was one beehive. Evaluation of bee and varroa mortality after the fungus application was carried out with an open cardboard with Vaseline that was placed on the floor of each hive and evaluated every 48 h over a period of 15 d. The individuals collected were incubated in a humid chamber to detect signs of mycosis. Results of mortality over time were adjusted to sigmoid curves and comparison of cumulative mortality means on day 15 post inoculation by Fisher’s protected test (p < 0.05). After 21 d post-treatment, beehives were evaluated to determine V. destructor infestation levels and 200 bees were collected from each hive and placed in a jar with water and detergent. They were subsequently shaken to separate the varroa from the bodies of the bees and the number of mites per bee was counted (SAG, 2002). Results were compared among treatments to identify the percentage of parasitized bees and the relationships between the final and initial populations (Pf/Pi), both submitted to analysis of variance and mean separation by the Tukey test (p < 0.05). Spring evaluation During the spring, four M. anisopliae (Qu-M845) applications were carried out every 4 d between September 1 and 16, and afterwards,, another four between October 16 and 31. All the doses were 5 x 1010 conidia per hive, applied between the frames of the breeding chamber. The control was beehives without any applications. Following the methodology described above, the effect of the fungus on the bees and mites during 12 d was evaluated by taking out a cardboard with Vaseline every 48 h. The experimental design was completely random with a beehive as an experimental unit, two treatments, and five replicates. The varroa and bee mortality curves for each inoculated hive were calculated twice: at the start of spring and 45 d later, and results were expressed as mortality over time by adjusting sigmoid curves. Cumulative mortality was compared on day 15 post inoculation with analysis of variance and mean separation with Fisher’s protected test (p < 0.05) (Gomez and Gomez, 1984). RESULTS AND DISCUSSION Laboratory trials The mites that died with the EPF treatment were incubated in humid chambers and showed symptoms of mycosis of the inoculated fungus, which was demonstrated by the presence of mycelium and spores in the legs and abdomen, whereas the control treatment showed no symptoms. The M. anisopliae Qu-M845 isolate produced 98% varroa mortality on day 7 post inoculation with a concentration of 108 conidia mL-1. The treatment with 107 conidia mL-1 produced a different value, 72% mortality. The 106 and 105 conidia mL-1 concentrations the varroa mortality were similar, 46 and 48%, respectively (p ≤ 0.05). During this period, mortality did not occur in the V. destructor control (Figure 1). CL50 and CL90, with 3.8 x 105 and 8 x 107 conidia mL-1, respectively (χ2 = 2.03) were obtained from the sigmoid formula (Table 1). Kanga et al. (2002) obtained CL90 for V. destructor of 1.37 x 109 conidia mL-1 of an M. anisopliae isolate on day 7 post inoculation, which was higher than that obtained in this trial. For other mite species like Tetranychus evansi, Wekesa et al. (2005) point out a lethal concentration between 0.7 x 107 and 2.3 x 107 conidia mL-1 to achieve the 50% mortality with diverse M. anisopliae isolates, values that are again higher than those found in our trials. Lethal time to kill 50% of the population (TL50) was calculated with the 1 x 107 concentration, value corresponding to the closest calculation of CL90. Results indicated TL50 of 1.35 d. Meikle et al. (2006) observed a lethal time of 4 to 7 d for varroa with a dose of 107 conidia mL-1 of M. anisopliae with an inoculation technique and maintenance of the mites similar to the one described. These values were higher than those that we found. Chandler et al. (2000) point out that death of the entomopathogenic fungi generally occurs in a period of 3 to 10 d after the infection, which is produced by water loss, nutrient deprivation, mechanical damage, and toxin action. Evaluation of application methods After the applications, treatments with fungi presented a V. destructor infestation significantly different and lower than the control. The treatment with conidia sprinkled on the frames presented a 67% decrease in the percentage of infested bees with respect to the mite population in the control (Figure 2A). Concerning to the variation of the populations (Pf/Pi), the control increased 1.7 times the initial population, treatments with stamped conidia and conidia dispenser maintained a relationship close to one which indicates that the varroa population did not grow. Lastly, the treatment with sprinkled conidia decreased 0.72 times the initial population (Figure 2B). Cumulative mite mortality during this period showed similar results when directly applying the conidia on and between the frames, and the conidia adhered to filter paper, but were statistically different to the results obtained with the other treatments (p = 0.0093) (Figure 3). Despite these results, an infestation level less than or equal to 3% was not obtained, level recommended as tolerable in the hive (Neira et al., 2004). However, these results were obtained at 21 d, and could be expected to continue decreasing as the Metarhizium inoculant managed to multiply itself in the dead varroa. Kanga et al. (2006) obtained similar results when evaluating plastic strips covered with M. anisopliae spores to control V. destructor in bee colonies in autumn. When evaluating cumulative bee mortality for each treatment, differences in the death of bees over time were observed, hives in which conidia were applied on and between the frames had greater bee mortality than the rest of the treatments; furthermore, they were similar (p < 0.05) to the control beehives (Figure 4). Out of 154 dead bees in the hives treated with conidia sprinkled on the frames, 35% showed mycosis. Though it has been reported that M. anisopliae can infect A. mellifera in laboratory trials, at this time, it has not been reported to cause epizootics between bees (Chandler et al., 2001). More recently, Metarhizium has been considered for controlling other bee pests such as the Aethina tumida (Coleoptera: Nitidulidae) parasite in South Africa from isolated strains from this insect (Muerrle et al., 2006). Gliñski and Buczek (2003) point out that the hard, waterproof cuticle, biochemical activity of the juice of the middle intestine, and its peritrophic membranes, along with the tracheal system form a physiological mechanism and barriers that effectively protect the cavity of the body against invasion by fungi in bees. Furthermore, the antimicrobial substances such as phytocides (volatile organic alelochemical antimicrobial compounds), and volatile essences present in the ingested food can also destroy bacteria and invasive fungi. However, the infections with high spore doses or infections caused by highly pathogenic fungi species can destroy the anatomy and physiology of these immunological barriers Gliñski and Buczek (2003). On the average, in the control beehives, 71 dead bees were recorded, but only 1.5% presented parasitism by Metarhizium. This can be explained by the transport of the conidia by the wind when they were applied, by drones taking the conidia from one hive to another, or by bees drifting between hives (Kanga et al., 2003), as well as for the proximity between treatments. Indeed, Meikle et al. (2007) found that the percentage of varroa infected with B. bassiana also increased in the control, suggesting conidia movement probably due to drifting bees. Kanga et al. (2003) indicate that conidia can be easily applied in an apiary, factor that can be beneficial for beekeepers. In accordance with this, and to the results obtained, it is concluded that the most effective application method is the pulverization of conidia on the frames. Spring evaluation When evaluating the effect of the fungus on the beehives in spring, significant differences were observed between the hives treated with the Qu-M845 isolate and the control hives (Figure 5). The mite mortality curve over time was adjusted to a sigmoid for both evaluation dates. On the two dates, the hives treated with fungi produced greater mite mortality (R2 = 0.98 and R2 = 0.95) for the first and second observation date, respectively. The percentage differences between treated and untreated hives was 87 and 52.4% (Figure 5 A and B). These results concur with those obtained by Meikle et al. (2007), who observed an increase in the percentage of infected mites in the hives treated with a B. bassiana isolate. No significant differences were observed in bee mortality among treatments for both application dates (September and October) (p ≤ 0.05) (Figure 6A and 6B). Neither were symptoms of mycosis observed in the collected bees, ruling out mortality caused by fungus. The lack of pathogenesis of M. anisopliae for other animals, including mammals, as well as the results obtained, indicate that this species is a promising alternative for the biological control of varroa. However, before recommending its use as a biological control agent of V. destructor or of other bee pests, more field evaluations are necessary to define the effects on beehives (Muerrle et al., 2006). CONCLUSIONS In laboratory conditions, the Metarhizium anisopliae var. anisopliae Qu-M845 native isolate was pathogenic for Varroa destructor. The values of the lethal concentrations were lower than in other studies. Conidia pulverized on and between the frames at a dose 600 times greater than CL90 considerablyreduced mite infestation in autumn and spring. These results lead us to consider M. anisopliae as a promising tool to control V. destructor. ACKNOWLEDGEMENTS The authors thank Sady Muena for his valuable collaboration in developing this research. LITERATURE CITED
Copyright 2009 - Chilean Journal of Agricultural Research The following images related to this document are available:Photo images[cj09065f3.jpg] [cj09065f1.jpg] [cj09065f4.jpg] [cj09065t1.jpg] [cj09065f5.jpg] [cj09065f6.jpg] [cj09065f2.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}