 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Chilean Journal of Agricultural Research, Vol. 70, No. 1, Jan-Mar, 2010, pp. 26-33 Presence of a phytoplasma associated with witches’-broom disease in Ugni molinae Turcz. and Gaultheria phillyreifolia (Pers. ) sleumer determined by DAPI, PCR, and DNA sequencing Presencia de un fitoplasma asociado a la enfermedad de “escoba de bruja” en Ugni molinae Turcz. y Gaultheria phillyreifolia (Pers.) Sleumer determinado mediante DAPI, PCR y secuenciación de ADN. Nolberto Arismendi S.1, Nancy Andrade S.[1]*, Ricardo Riegel Sch.1, and Roberto Carrillo Ll.1 1Universidad Austral de Chile, Facultad de Ciencias Agrarias, Casilla 567, Valdivia, Chile. *Corresponding author (nandrade@uach.cl). Received:
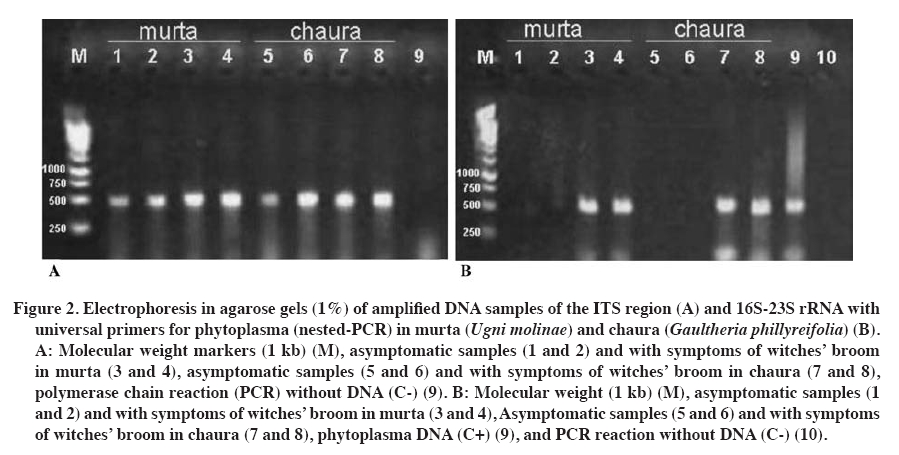
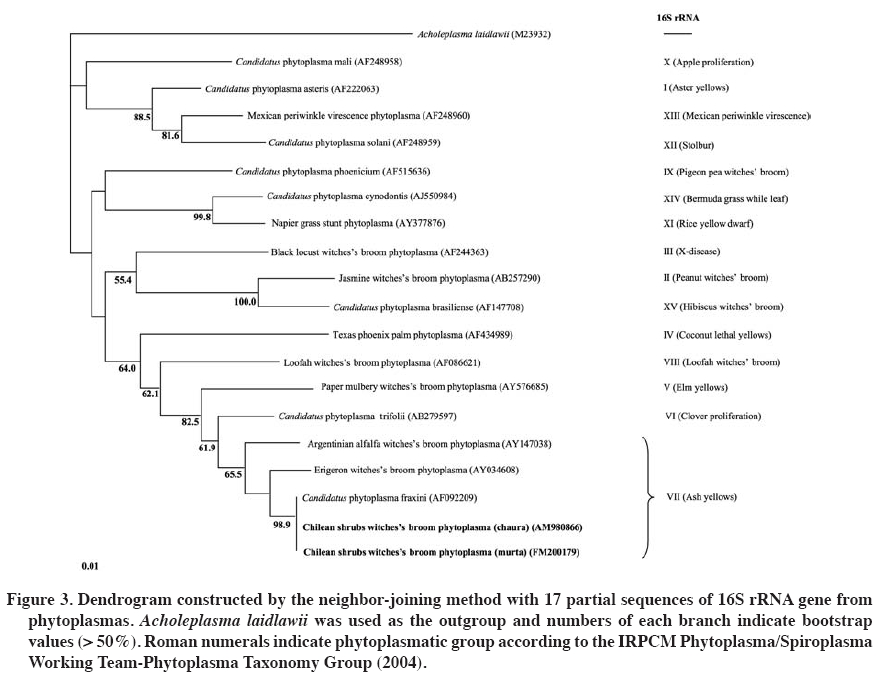
31 December 2008. Code Number: cj10003 ABSTRACT Murta (Ugni molinae Turcz.) and common chaura (Gaultheria phillyreifolia (Pers.) Sleumer) are native species of Chile. Plants of both species have shown over-branching like witches' broom. The causal agents of these symptoms in many plants are phytoplasma. To verify the presence of these microorganisms, DAPI (4',6-diamidino-2-phenylindole) staining analysis and polymerase chain reaction (PCR) were performed in symptomatic and asymptomatic plants. Positive PCR samples were sequenced to identify the pathogens involved. In individuals of both species with witches’ broom symptoms, DAPI staining showed fluorescent bodies in the phloem tissues, but not in asymptomatic plants. Verification by nested-PCR, phytoplasmatic DNA was amplified from diseased murta and chaura, but not in apparently healthy plants. Sequencing of amplified products allowed locating phytoplasma within the ash yellows group (16SrVII) and related to Candidatus phytoplasma fraxini. This is the first report of phytoplasma in Chilean native species. Considering the diversity of plant species infected by the ash yellows group suggests that G. phillyreifolia and U. molinae could be a phytoplasma reservoir for other economically important agricultural crops. Key words: native shrubs, witches’ broom, phytoplasma, microscopic and molecular tools. RESUMEN La murta(Ugni molinae Turcz.) y la chaura común (Gaultheria phillyreifolia (Pers.) Sleumer) son especies nativas de Chile. En plantas de ambas especies se ha observado una sobre-ramificación de tipo “escoba de bruja”. En muchas plantas los agentes causales de esta sintomatología son fitoplasmas. Para verificar la presencia de estos microorganismos se analizaron plantas con y sin síntomas mediante tinciones DAPI (4’,6-diamidino-2-fenilindol) y reacción en cadena de la polimerasa (PCR). Muestras positivas en la PCR fueron secuenciadas para identificar al fitopatógeno implicado. En individuos de ambas especies con síntomas de escoba de bruja, la tinción DAPI permitió observar cuerpos fluorescentes en los tejidos del floema, situación que no ocurrió en plantas asintomáticas. En la verificación mediante PCR-anidada, se logró amplificar ADN fitoplasmático en plantas de murta y chauraenfermas, pero no en plantas aparentemente sanas. La secuenciación de los productos amplificados permitió localizar al fitoplasma dentro del grupo “ash yellows” (16SrVII) y relacionado al “Candidatus Phytoplasma fraxini”. Éste es el primer reporte que caracteriza a un fitoplasma en especies nativas chilenas. Considerando la diversidad de especies de plantas infectadas por el grupo ash yellows, sugiere que G. phillyreifolia y U. molinae podrían constituir un reservorio de fitoplasmas para otros cultivos agrícolas de importancia económica. Palabras clave: arbustos nativos, escoba de bruja, fitoplasma, técnicas microscópicas y moleculares. INTRODUCTION Murta (Ugni molinae Turcz., Myrtaceae) and common chaura (Gaultheria phillyreifolia (Pers.) Sleumer, Ericaceae) are native species of Chile that share a similar environment and distribution (between 34º and 54º S lat) given that murta has been reported from the Maule Region to Aysén del General Carlos Ibáñez del Campo Region (Seguel and Torralbo, 2004) and chaura from the Maule Region to the Magallanes Region and the Chilean Antarctic (Donoso and Ramírez, 1997). Both species grow in varied conditions, invading deforested, marginal lands with low fertility, as part of the forest and prairie undergrowth and ecotones (Donoso and Ramírez, 1997; Seguel and Torralbo, 2004). Murta is considered a species with high agronomic and pharmaceutical potential, principally for its organoleptic, antioxidant, and analgesic attributes (Seguel and Torralbo, 2004; Delporte et al., 2007; Suwalsky et al., 2007). On the other hand, chaura is an ornamental species with little impact on the national market. There are few records of pests and diseases that can affect the development of both species. However, in murta and chaura plants, a strong reduction has been observed in the growth of shoots, causing leaves to develop abnormally, giving rise to over-branching that later dies. This type of symptomatology is known as witches’ broom (Figure 1). The causal agents of this symptomatology can be varied (biotic and abiotic), but in the majority of cases, they have been related to phytoplasmas (Ghosh et al., 1999; Montano et al., 2001; Abou-Jawdah et al., 2002; Barros et al., 2002; Khan et al., 2002; Marcone et al., 2004; Al-Zadjalt et al., 2007). Since their detection in the 1960s, a large number of diseases caused by phytoplasmas have been pointed out that affect hundreds of species worldwide (Lee et al., 2000; Bertaccini, 2007). The problem is not new in Chile since there are reports of some economically important crops (Hepp et al., 1998; Hepp and Vargas, 2002; Herrera and Madariaga, 2003; Fiore et al., 2007; Matus et al., 2008). Phytoplasmas are obligate parasites that develop and reproduce asexually in the cytoplasma of their host cells (insects and plants) (Lee et al., 2000; Weintraub and Beanland, 2006). The fact that it is impossible to cultivate these microorganisms in vitro has limited their characterization or identification (Schneider et al., 1997). Although microscopic techniques are still important for detecting phytoplasmas (Chapman et al., 2001; Fránová et al., 2007), molecular techniques are indispensable for determining taxonomic and phylogenetic relationships between phytoplasmas and other microorganisms (Lee et al., 1998; 2000). Universal primers, but specific for phytoplasmas, based on the regions of the 16S rRNA gene, intergenic region, and 23S rRNA gene are used widely for detecting these prokaryotes (Smart et al., 1996; Heinrich et al., 2001). Furthermore, sequencing of polymerase chain reaction (PCR) products and restriction fragment length polymorphism by PCR (PCR-RFLP) are elementary techniques in the identification and characterization of phytoplasmatic groups (Lee at al., 1998; 2000). However, the majority of phytoplasma reports in Chile are based on electronic microscopy and DNA amplification with PCR using universal primers (Hepp et al., 1998; Villagra, 2001; Hepp and Vargas, 2002; Herrera and Madariaga, 2003). Only recent work has involved PCR-RFLP and DNA sequencing for a better identification of these prokaryotes (Fiore et al., 2007; Matus et al., 2008). The aim of this research was to determine the presence of phytoplasmas in U. molinae and G. phillyreifolia plants with symptoms of witches’ broom. Microscopic techniques such as DAPI staining (4',6-diamidino-2-phenylindol) and molecular techniques such as specific amplification of phytoplasmatic DNA with PCR and subsequent sequencing were used. MATERIALS AND METHODS Samples of stems and petioles of young U. molinae and G. phillyreifolia plants with and without symptoms of witches’ broom disease were taken from localities near the city of Valdivia (39°49' S; 73°12' W), Chile. DAPI staining was based on the Romero (2001) protocol: 1 cm long pieces of symptomatic and asymptomatic stems and petioles were cut and put in 5% glutaraldehyde (5 mL glutaraldehyde and 95 mL 0.1 M buffer phosphate pH 6.8) for 15 min. Samples were then washed in 0.1 M buffer phosphate pH 6.9 for 5 min. Immediately after, longitudinal cuts of 10 to 15 microns were made on the samples to be stained with DAPI staining (0.1 mg of 4',6-diamidino-2-phenylindole in 100 mL of sterile water) for 20 min. Finally, the preparations were observed with a fluorescent microscope (BP 450-490, FT 510 and LP 520 filters; Carl Zeiss Axiolab, New York, USA) with a UV light excitation longitude of 450-490 nm with an oil immersion lens 100X and ocular lenses 10X to achieve a maximum magnification of 1000X. For genomic DNA extraction (plant and phytoplasma DNA), the protocol prepared by Lodhi et al. (1994) was followed along with the following modifications: 200 mg of each sample were ground in the presence of liquid nitrogen and dissolved in 500 µL of CTAB-PVP 2% (Tris-HCl 100 mM pH 8.0; NaCl 1.4 M; EDTA 20 mM pH 8.0; 2% CTAB w/v; 0.2% 2-mercaptoethanol v/v and 2% polyvinylpyrrolidone PVP40-50G w/v) extraction buffer (pre-heated at 60 ºC) and incubated at 65 ºC for 30 min. Then, 500 µL of cold (-20 ºC) chloroform-octanol (24:1) was added and centrifuged for 5 min at 6000 g. Next, the top phase was extracted to which was added 350 µL of chloroform-octanol (24:1). Centrifugation was repeated and once again the top phase was extracted to which 175 µL of ammonium acetate 7.5 M and 370 µL of cold isopropanol were added. These reagents were mixed by inversion and left to precipitate all night at 4 ºC. Subsequently, they were centrifuged for 30 min at 13600 g and the DNA precipitate was washed with 500 µL of cold ethanol, followed by another centrifugation (13600 g; 5 min) with the ethanol being eliminated after each washing. Finally, the precipitate formed was left to dry at room temperature for 30 min and was resuspended in 100 µL of sterile distilled water. DNA quantification was carried out with a spectrophotometer (Nanodrop ND-1000, Wilmington, Delaware, USA) and its quality was verified by PCR for the ITS regions (present in the plants), using the ITS3 and ITS4 primers (Van der Stappen et al., 1998). To determine the presence of phytoplasmatic DNA, a preamplification was performed with a pair of universal primers for phytoplasmas (PA2F: 5’gccccggctaactatgtgc 3’; PA2R: 5’ ttggtgggcctaaatggactc 3’) (Heinrich et al., 2001). A nested-PCR was carried out from this first reaction using a new pair of primers with greater specificity and sensitivity (NPA2F: 5’ atgacctgggctacaaacgtga 3’; NPA2R: 5’ ggtgggcctaaatggactcg 3’) that magnified a region of 485 bp which corresponds to position 1182 of the 16S rRNA gene and 1667 of the intergenic region between the 16S-23S rRNA genes (Heinrich et al., 2001). Preamplification was performed in a volume of 25 μL per sample, divided into: 2.5 μL of PCR 10x buffer; 1.0 μL of MgCl2 (25 mM); 0.5 μL dNTP (2 mM); 0.5 μL of each primer (10 pmol μL-1); 0.2 taq ADN polymerase (5 U μL-1, Fermentas Inc., Glen Burnie, Maryland, USA), 18.8 μL sterile distilled water with 1.5% PVP, and 1 μL of DNA (25 ng μL-1). The thermal profile of the 35 cycles of the PCR (MJ Research, PTC-100 Thermal Cycler) consisted in denaturalization at 94 ºC for 1 min (3 min for the first cycle), alignment at 60 °C for 1 h 15 min, and elongation at 72 ºC for 1 h 30 min (10 min for the last cycle). For the nested-PCR, basically the same preamplification conditions were used, only changing the primers and eliminating PVP. For this magnification, 40 cycles were carried out at 94 ºC for 1 min (3 min for the first cycle), 60 ºC for 50 s, and 72 ºC for 1 h 30 min (7 min for the final elongation). Phytoplasmatic DNA was used as a positive control (ACLR-AY isolate provided by N. Fiore, Universidad de Chile). Products of the PCR reaction were run in 1% agarose gels with 0.1 μL mL-1 of ethyl bromide in an SB buffer (Brody and Kern, 2004) at 100 V for 30 min. Subsequently, the amplicons were observed and photographed on a UV light transilluminator. The DNA magnified by PCR was purified and sequenced (Macrogen, Seoul, South Korea) using the modified Tint primer (Smart et al., 1996) (5’ aggcgtgtgctctaac 3’) to avoid sequencing of contaminating DNA. Phytoplasma sequences obtained from chaura and murta were recorded in the database of the European Bioinformatics Institute (EBI, Cambridge, UK) as the accession numbers AM980866 and FM200179, respectively. These sequences were compared and identified (FASTA-EBI) with others in the EBI database. The dendrogram was constructed with the ClustalX program by the neighbor-joining method with 1000 bootstraps to relate 17 phytoplasmatic sequences involving 15 groups classified on the basis of the 16S rRNA gene according to IRPCM Phytoplasma/Spiroplasma Working Team-Phytoplasma Taxonomy Group (2004). To achieve a valid comparison, the 17 sequences were reduced to the longitude of the isolated sequences in this study. Acholeplasma laidlawii (Mollicutes) was used as the dendrogram root. RESULTS AND DISCUSSION DAPI staining U. molinae and G. phillyreifolia plant samples presenting symptoms of witches’ broom showed a high concentration of fluorescent bodies in the phloem cells (Figure 1). Small points and aggregations near the cell wall in the cells of the conductor tissue were observed. On the contrary, tissues of plants without symptoms of the disease did not present fluorescence in the phloematic cells. DAPI is a low cost and rapid method for detecting phytoplasmatic bodies in different plant tissues, but is not a specific tool since it can stain DNA of other microorganisms or organelles such as mitochondria and chloroplasts (Fránová et al., 2007). Therefore, this technique has only been used as a preliminary diagnostic tool and is mainly complemented by PCR (Jarausch et al., 1998; Cordova et al., 2003; Bricker and Stutz, 2004; Canik and Ertunc, 2007). PCR Amplification of the ITS region allowed satisfactorily copying the plant DNA fragment (500 bp) in all the U. molinae and G. phillyreifolia samples evaluated (Figure 2A). This control was essential to verify the absence of inhibitors of the PCR reaction such as polysaccharides and/or phenolic compounds, habitual resistance components in plants suffering some kind of stress due to pathogens (Lherminier et al., 2003). No phytoplasmatic DNA amplicons were achieved in preamplification (first PCR reaction) in the murta, chaura, and positive control samples evaluated. However, carrying out a nested-PCR (second PCR reaction) allowed obtaining DNA bands of an approximate length of 485 bp (Figure 2B) for the samples that presented symptoms of witches’ broom. A fragment of similar size was observed for the positive control of phytoplasmas. On the other hand, no DNA bands were visualized in the asymptomatic tissue samples (Figure 2B). The fact that the detection of phytoplasmatic DNA was achieved only with a nested-PCR is attributable to its low concentrations as compared to plant DNA and the possible presence of remains of the PCR inhibitor compounds (Green et al., 1999; Khan et al., 2004). In the second PCR reaction, conditions are optimized and the sensitivity of the technique is increased (Berges et al., 2000) facilitating visualization of the amplicons. Sequencing and identification DNA sequences between 392 and 416 bp (accession numbersFM200179 in U. molinae and AM980866 in G. phillyreifolia) were obtained with the sequencing of the amplicons generated from the diseased tissue of both species. Both sequences showed a 100% homology one with the other, as well as high homologies with other phytoplasma sequences. Alignment and construction of the dendrogram with sequences from the 16S rRNA gene and part of the 16S-23S rRNA intergenic region of 17 phytoplasmas allowed relating the derived phytoplasma sequences of murta and chaura (from now on referred to as Chilean shrub witches’ broom phytoplasma) in the ash yellows group (16SrVII) (Figure 3). These partial sequences were highly similar (99.8%) to Candidatus Phytoplasma fraxini, and to a lesser degree with Erigeron witches' broom phytoplasma (97.3%) (accession number AY034608), and Argentinean alfalfa witches'-broom phytoplasma (95.1%) (accession number AY147038); all of which are from the ash yellows group. These results confirm the preliminary evaluations carried out with DAPI, indicating that the observation of fluorescence in the phloematic tissues coincides with the presence of phytoplasmas, similar to another works made on other species (Cordova et al., 2003; Bricker and Stutz, 2004; Canik and Ertunc, 2007; Fránová et al., 2007). This is one of the first reports of the detection and identification of native plant phytoplasmas in Chile. Previously, with the help of electronic microscopy and PCR, witches’ broom disease had been associated to phytoplasmas in U. molinae, but without identifying them (Villagra, 2001). In G. phillyreifolia, no report had been presented indicating the nature of the causal agent in the witches’ broom symptomatology. Both chaura and murta are shrubs that grow together in southern Chile, situation benefiting the mixed infection of the phytoplasma since recent studies have demonstrated that the Carelmapu ramosi Linnovuori & DeLong(Cicadellidae) insect is related as the principal vector of the phytoplasma associated with witches’ broom disease in G. phillyreifolia (Arismendi et al., 2008). The same species of insect had already been identified as one of the possible vectors of phytoplasma in murta (Miño, 2003), circumstance favoring the plant-phytoplasma-insect interaction. Phytoplasmas of the ash yellows group (16SrVII) in America have been detected in different hosts and mainly related to diseases of the witches’ broom type (Griffiths et al., 1999; Feeley et al., 2001; Barros et al., 2002; Conci et al., 2005). Within the ash yellows group, phytoplasmas reported in Argentina (Argentinean witches’-broom phytoplasma) and Brazil (Erigeron witches’ broom phytoplasma) are associated with the 16SrVII-C and 16SrVII-B subgroups, respectively (Barros et al., 2002; Conci et al., 2005). The high bootstrap (98.9%) generated in the dendrogram suggests that Chilean shrub witches’ broom phytoplasma is related more to the Candidatus Phytoplasma fraxini (16SrVII-A) than to the two previously mentioned subgroups (Figure 3). Recent studies in Chile have reported the presence of phytoplasmas of the 16SrVII-A subgroup associated with yellow grapevine disease (Fiore et al., 2007). However, it is still not possible to determine with the currently available information if native species such as chaura and murta could act as important phytoplasma reservoirs for other economically important crops. Furthermore, future research should establish the aspects of temporal and spatial dispersion of this type of phytoplasma, aspect associated with the feeding habit and distribution of the insect vector(s). CONCLUSIONS Microscopic evidence (DAPI staining) and molecular evidence (PCR and DNA sequencing) indicate that witches’ broom disease in U. molinae and en G. phillyreifolia is associated with a phytoplasma within the ash yellows group (16SrVII), relating that sequences more with the 16SrVII-A subgroup than to the other two subgroups reported on. ACKNOWLEDGEMENTS This research was financed by Project DID S-20066 of the Universidad Austral de Chile. We are grateful to Nicola Fiore of the Universidad de Chile for providing the phytoplasmatic positive control (C+). LITERATURE CITED
Copyright © 2010 - Universidade de Santa Cruz do Sul The following images related to this document are available:Photo images[cj10003f1.jpg] [cj10003f2.jpg] [cj10003f3.jpg] |
| |||||||||

{kind=link}
{kind=link}
{kind=link}