 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
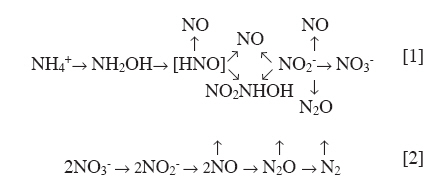
Chilean Journal of Agricultural Research, Vol. 70, No. 3, July-September, 2010, pp. 485-497 Review Greenhouse gas (CO2 and N2O) emissions from soils: A review Emisión de gases invernadero (CO2 y N2O) desde suelos: Una revisión. Cristina Muñoz1, Leandro Paulino1, Carlos Monreal2, Erick Zagal1 1 Universidad de Concepción, Facultad de Agronomía, P.O. Box 537, Chillán, Chile Correspondence Address:Erick Zagal, Universidad de Concepción, Facultad de Agronomía, P.O. Box 537, Chillán, Chile, ezagal@udec.cl Date of Submission: 31-Aug-2009 Code Number: cj10053 Abstract In agricultural activities, the main greenhouse gases (GHG) are those related to C and N global cycles. The impact of agriculture on GHG emissions has become a key issue, especially when considering that natural C and N cycles are influenced by agricultural development. This review focuses on CO2 and N2O soil emissions in terrestrial ecosystems, with emphasis in Chilean and similar agro-ecosystems around the world. The influence of land use and crop management practices on CO2 and N2O emissions is analyzed; some mitigation measures to reduce such emissions are also discussed here. More knowledge on the biological processes that promote of GHG emissions from soil will allow creating opportunities for agricultural development under friendly-environmental conditions, where soil can act as a reservoir and/or emitter of GHG, depending on the balance of inputs and outputs.Keywords: Global warming, carbon dioxide, nitrous oxide, soil biological processes, soil organic carbon, nitrification, denitrification Resumen En actividades agrícolas los principales gases de efecto invernadero (GHG) son los relacionados con los ciclos globales de C y N. El impacto de la agricultura sobre las emisiones GHG se ha convertido en una cuestión clave, especialmente si se considera que los ciclos naturales C y N se ven influidos por el desarrollo agrícola. Esta revisión se centra en emisiones de CO2 y N2 O del suelo en los ecosistemas terrestres, con énfasis en agro-ecosistemas de Chile y similares alrededor del mundo. Se analiza la influencia del uso del suelo y las prácticas de manejo del cultivo sobre emisiones de CO2 y N2 O, se discuten medidas de mitigación para reducir estas emisiones. Un mayor conocimiento sobre los procesos biológicos que promueven las emisiones GHG del suelo permitirá la creación de oportunidades para el desarrollo agrícola en condiciones ambientalmente amigables, donde el suelo puede actuar como un reservorio y/o emisor de GHG, dependiendo del balance de entradas y salidas. Palabras clave: calentamiento global, CO2 , N2 O, procesos biológicos del suelo, carbono orgánico del suelo, nitrificación, desnitrificación. Introduction Over the last few decades, climate change has been studied by researchers in different disciplines, who have predicted an increase in the temperature of the atmosphere and oceans mainly due to the emissions of greenhouse gases (GHG) such as carbon dioxide (CO2 ), methane (CH4 ), nitrous oxide (N2O), troposphere ozone (O 3 ) and chlorofluorocarbons (CFCs) (IPCC, 2007). For the past two centuries and especially after the Industrial Revolution, emissions of GHG have increased dramatically due to increased industrial and transportation activities associated with the use of fossil energy sources and land use changes (IPCC, 2007). In this sense, the Intergovernmental Panl on Climatic Change (IPCC, 2007) reported that the global atmospheric concentration of CO2 has increased from a pre-industrial value of about 280 to 379 ppm in 2005, exceeding the natural range over the last 650 000 yr (180 to 300 ppm, as determined from ice cores), and that the annual CO2 concentration growth-rate was larger during the last 10 yr (1995 to 2005 average: 1.9 ppm per year). The global atmospheric concentration of CH4 has increased from a pre-industrial value of about 715 to 1774 ppb in 2005, exceeding the natural range of the last 650 000 yr (320 to 790 ppb). In addition, the global atmospheric N2O concentration increased from a pre-industrial value of about 270 to 319 ppb in 2005, being the growth rate relatively constant since 1980 considering that more than a third of all N2O emissions are anthropogenic and primarily due to agriculture. Current GHG emissions generated by anthropogenic activities include land use and land use change in agricultural and forest systems, industrial development, and urban expansion, among other sources. All of them have contributed to disrupt the C and N cycles in terrestrial ecosystems (IPCC, 2007). The contribution of agricultural soils to CO2 , N2O and CH4 emissions depends on the biophysical processes, and the incorporation/decomposition of organic residues in the soil. Soil aerobic conditions produce CO2 , anaerobic conditions produce CH4 , and nitrification and denitrification processes of mineral-N result in N2O emission. Natural sources of CO2 basically correspond to the respiration process of terrestrial and aquatic organisms. An estimated 60 Pg C yr -1 (1 Pg = 1 x 10 15 g) is emitted to the atmosphere by autotrophic respiration and a similar amount is emitted as a result of heterotrophic respiration (Reay and Grace, 2007), where the CO2 emission resulting from soil respiration is 10 to 15 times greater than the CO2 emission from fossil fuels (Raich and Schlesinger, 1992). The emission of CO2 due to volcanic activity is relatively minor on a global scale, accounting for 0.02 to 0.05 Pg C yr -1 . The impact of biomass burning events on atmospheric CO2 concentration is relatively short-term due to rapid uptake of CO2 during regrowth of vegetation (Reay and Grace, 2007). Stored C in terrestrial vegetation (afforestation and reforestation), CO2 absorption by the oceans (C dissolves as bicarbonate and carbonate ions), and C sequestration by the soil can potentially serve as CO2 sinks (Lal, 2006; Reay and Grace, 2007). Soils of the world can act as sinks or sources of GHG emissions, and the net balance between the absorbance of atmospheric C and its release determines the net temporal status of soils as either C absorbers (inputs) or releasers (outputs). Soils altogether contain an estimated 2500 Pg C being 3.3 times the atmospheric pool (Lal, 2006). Using the IPCC emission factors, Vergé et al. (2007) obtained projections of total emissions of GHG for South America of 757, 966 and 1207 Tg CO2 (1 Tg = 1 x 10 12 g) for the years 2000, 2015 and 2030, and with total GHG emissions per capita in the range of 2180; 2313 and 2558 kg CO2 equivalents in same period of time. Land use and land use change have an important role in the increase of N2O emissions, where the production of this gas from soils generally respond to natural or anthropogenic processes and to specific environmental conditions. The relevance of N2O to global warming is mainly attributed to two physico-chemical characteristics: (1) N2O is stable and remains in the atmosphere approximately 120 yr, and (2) N2O has a large global warming potential (GWP) that is 296 times greater than CO2 in a 100 yr period, which is related to the catalytic destruction of stratospheric ozone. The N2O is considered responsible for 4 to 5% of global warming (IAEA, 1992; Reay et al., 2007b), being the tropical and temperate non-cultivated soils the main sources and accounting for 63% of N2O total emissions (Reay et al., 2007b). When compared in terms of mass, GWP of methane (CH4 ) has been recently estimated at 23 times that of one molecule of CO2 in a 100 yr period (Reay et al., 2007a). As methane is produced under anaerobic conditions, seasonal or permanently flooded systems are important sources since they favor the development of methanogenic microorganisms (Reay et al., 2007a). The IPCC report (2001) indicates that about half of the global CH4 emissions are from natural sources, with estimates between 100 to 250 Tg CH4 yr -1 from wetlands, and 320 Tg CH4 yr -1 as a result of energy use, waste decomposition, ruminants, and rice crop, among others (Reay et al., 2007a). An extensive review about the biological processes and environments that promoted the CH4 emissions from soils was published by Le Mer and Roger (2001) and Lai (2009). This review focuses on soil components and processes that control CO2 and N2O emissions from terrestrial ecosystems by analyzing specific information available from Chilean ecosystems, or other ecosystems with similar edapho-climatic conditions, and identifying mitigation strategies. Sources of CO2 and N2O from Different Soils in Agricultural Systems Carbon dioxide emissions from soils In terrestrial ecosystems, natural processes of C transformation occur mainly in the soil, where biogeochemical activities and abiotic factors, such as climate, regulate the internal cycles and flows of the organic and inorganic forms of these elements (Monreal et al., 1997b). It is also important to indicate that net CO2 emissions result from the amount of atmospheric C, fixed through photosynthesis and stored in soil as organic matter, and the amount of soil C oxidized to CO2 during a given period. However, it is also necessary to consider that the principal source of net CO2 emissions in the atmosphere is from erosive and oxidative processes associated with agricultural crops (Follett et al., 2001). Thus, when discussing GHG emissions from agricultural land, both processes need to be considered. Land use (LU), land use change (LUC) and management practices alter the long-term steady-state level of soil organic C and N in soils. These activities, along with climatic variables, can potentionally increase or decrease soil biological activity associated with heterotrophic decomposition of soil organic matter (SOM). Bernoux and Volkoff (2006) estimated that about 5000 Tg C was lost from soils by past agricultural activities in Latin America, indicating that some ecoregions suffered important C losses constitute a potential future sink if the tendency can be reversed, like the case of Central Brazilian and Central American regions. The C loss in South Chile was estimated lesser than other regions, with 64 Tg C in agricultural areas and 12 Tg C in mosaic of native and agricultural areas. The CO2 emitted from soils is a useful indicator to determine energy flow patterns, specifically the mineralization of nutrients and the rate of organic material decomposition. The CO2 flow from soils is the result of root respiration and physiological processes of the microorganisms involved in the decomposition of organic material. Emissions of CO2 from soils appear to be highly variable in heterogeneous soil micro-sites, and they are influenced by the activity of roots, microbial processes, crop residue and litter content, microclimate and catalytic properties of clay colloids (Matteucci et al., 2000). In [Table - 1] is possible to observe the range of CO2 emission in different countries, with an extensive range of CO2 emission from different soil use and management, e.g. 0 to 31 Gg CO2 -C ha -1 yr -1 (1 Gg = 1 x 10 9 g) for different cropping systems, shrub land/natural landscape and pastures systems. Chile presented in 1999 the First National Communication to the Conference of the Parties to the United Nations Framework Convention on Climate Change (FCCC) and showed the production of GHG (expressed as equivalent of CO2 ) mainly from agriculture, change in land use and forestry. In this sense, the national inventory of GHG for Chilean agriculture (Novoa et al., 2000; DICTUC, 2004; González et al., 2009a) is carried out through empirical estimations based on factors established by international experts associated with the IPCC. The last national GHG inventory of Chile (González et al., 2009b) reports that the Agriculture Sector produces GHG emissions equal to 13.3 Gg of CO2 equivalent (CO2 e) yr -1 for 2003, showing an increase in GHG emissions compared to 1984 (10.1 Gg of CO2 equivalent, CO2 e, yr -1 ), with a global increase of about 32%. This information is a relevant estimation of the contribution of agriculture to GHG emissions, but there is a clear need to determine emission factors for different regions and edapho-climatic and management systems throughout the country and reduce the uncertainty in order to improve future national reports. In this context, it is also important to note that the great extension of Chile and its different edapho-climatic conditions make difficult the determination of emission factors under all conditions, being necessary to focus on the areas with the highest production of GHG emissions. For example, in the [Figure - 1] is possible to observe that approximately 53% of the productive soil surface in Chile correspond to natural and improved grasslands, and the agricultural soils account for almost 10% of soil surface, being concentrated in Central zone of Chile (INE, 2007). In spite of the small surface of agricultural soils respect to the total area of the country, high amounts of inorganic fertilizer are applied; in this regard FAO (2009) indicates that the use of fertilizer including N, P and K, in temporary and permanent cropland of Chile increased from 246 to 343 kg ha -1 yr -1 between 2002 and 2007, respectively. Because of this, the agricultural soils could be considered as a potential area of high GHG fluxes. However, measurements in situ of CO2 emission for agricultural soils from Chile are lacking. [Table - 2] shows the range of CO2 emissions from different soils in Chile under laboratory experiments. Here, it is possible to observe great differences in CO2 emissions among different soils and managements, with a range of 2 to 194 μg CO2 -C g -1 d -1 from some different forest, agroforestry and cropping systems. Scientists have already acknowledged the importance of have appropriate inventories and uncertainty estimates. However, there are few studies on the uncertainties in GHG inventories. Uncertainty estimations in national GHG inventories have become part of the Intergovernmental Panel on Climatic Change (IPCC) Good Practice Guidance (IPCC, 2000). Winiwarter and Rypdal (2001) indicated an overall GHG emission uncertainty between 10.5% and 12% for CO2 , CH4 and N2O in Austria, depending on the year. They showed that much of the uncertainty derived from lack of understanding of the biological processes associated with GHG emissions from soils. Gregorich et al. (2005) determined discrepancies among the emission factors proposed by the IPCC under some given conditions in Canada. Thus, it is essential to get a reliable database with GHG emission factors for each ecological region that will enable regional and national GHG inventories decrease their emission uncertainties. Process of nitrous oxide emissions from soils Nitrous oxide is a product of microbial metabolism during the process of nitrification (Equation 1) and denitrification (Equation 2) (Robertson and Groffman, 2007).
As shown in Equation 1, nitrification is defined as an ammonia oxidation process in soil, which may be relevant on the production of N2O in the second step of this reaction Equation 2. Singh and Tyagi (2009) extensively described the mechanisms and related microorganisms that promote nitrification in soil, indicating that the autotrophic and heterotrophic microorganisms may act as protagonists of this process. The groups of bacteria that transform the ammonium to nitrate are Nitrosomonas, Nitrosolobus, Nitrosovibrio, Nitrosopira and Nitrosococcus genus, where the overall nitrification process is controlled by ammonium and oxygen concentrations (Singh and Tyagi, 2009). Forest soils usually provide acidic conditions which may select microorganisms to different processes according to the tolerance of pH ranges. Brierley and Wood (2001) proved that heterotrophic bacteria and fungi promote nitrification in acid soils of coniferous forests in Western Europe, where bacteria of Arthrobacter sp. seem to be the most highly adapted to generate heterotrophic nitrification. Microbial denitrification occurs when nitrate is present in anaerobic microsites, where the oxygen demand exceeds its supply, under water saturation or where the local O 2 demand is very elevated (FAO, 2001). The positive relationship between nitrification and denitrification with the production of N2O has been proven (Saggar et al., 2004; Singh and Tyagi, 2009), and it has also been established that spatial and seasonal variability, structural conditions of soil microsites, soil water content, C and N availability and soil temperature are the most important variables in the production of N2O from volcanic soils of agricultural and forested ecosystems. In many terrestrial ecosystems, the microbial activity and abiotic factors are responsible for the efficiency of the N cycle, especially where the N availability in ecosystems is limited. In pristine ecosystems, such as the forests of South-Central Chile, the efficiency pattern of the N cycle still reflects the natural processes developed along the Holocene (Perakis and Hedin, 2002) and the physical-chemical characteristics of volcanic soils present in these environments (Chorover, 2002), resulting in low contributions of soil N gases to the atmosphere (Godoy et al., 2003). The paradigm of the N conservation in pristine soils is based on the NH4 + consumption by microorganisms and vascular plants by net primary production. This N conservation mechanism is supported by the scarce production of NO3 - in these soils, where the nitrification would require an extra-consumption of energy at the ecosystem level (Huygens et al., 2007). As result of this N conservation mechanism, elevated quantities of N, immobilized by microbial action and adsorption of inorganic forms of N onto clay colloid surfaces, have been observed (Bengtsson and Bergwall, 2000; Huygens et al., 2008). Anthropogenic activities associated with agriculture and forestry disrupts the closed N cycle causing lower cycle efficiencies in the ecosystem. Land use change from native forest to forest plantations (e.g. Pinus radiata) and grassland significantly increased N mineralization and nitrification in soils of New Zealand (Parfitt et al., 2003). Burger and Jackson (2003) indicated that soil N transformation rates were significantly higher in agricultural land with greater availability of organic substrates than in soils with lower substrate availability. In addition, it has been stated that soil management practices such as tillage (Chirinda et al., 2010), fertilization (Allen et al., 2010; Cardenas et al., 2010) and lime (Galbally et al., 2010; Zaman and Nguyen, 2010), legume crop (Huth et al., 2010), or grazing (Saggar et al., 2004; Luo et al., 2010) affect N2O production. In the [Table - 3] is shown N2O emission from different cropping systems with similar characteristics to Chile, where the values of emissions are in the range of 0 to 44 kg N2O-N ha -1 yr -1 . Special attention should be paid to "hot spots", which are places or events that are characterized by potentially high N2O emissions, where the combined effect of urine, dung, compaction and other impacts on the soil structure, favoring high nitrification/denitrification rates, should be considered. In this context, is possible to observe a range of 0 to 156 kg N2O-N ha -1 yr -1 in pasture systems with applications of animal waste in different countries [Table - 3]. Mitigation of GHG Emissions Mechanisms of C sink by soils At present, among the processes associated with the C capture in soils, it is important to consider the potential role of many agricultural soils as C sinks (Matus et al., 2006; Muñoz et al., 2007a; Martínez et al., 2008). Atmospheric C may be allocated into SOM reservoirs with different mean residence times from months or few years in labile C pools to hundred years in recalcitrant C pools (Monreal et al., 1997a). Several management practices and technologies help mitigate GHG emissions from soils. Soil conservation practices decrease CO2 emissions through the reduction of cultivation, protecting the soil surface with crop residues, and increasing the N use efficiency by crops (Hobbs et al., 2008). Diverse evidence shows that no-till system increases the C content in the soil, being an effective strategy for CO2 sequestration. Bayer et al. (2006) evaluated different tillage systems in sites with more than 10 yr of antiquity, and compared conventional tillage, reduced tillage and no tillage in two Brazilian Oxisols. They concluded that the C stocks in no-till systems produced a mean rate of C accumulation of 0.35 Mg ha−1 yr−1 (1 Mg = 1 x 10 6 g) higher than the other evaluated systems in tropical soils. The conversion from conventional till to no till restored some of the depleted SOC and N pools in a long-term tillage experiments in different soils of Ohio, USA (Mishra et al., 2010). In experiments of conventional tillage, rotation is an important factor. For example, in a long-term experiment in an Andisol of South-Central Chile, Zagal and Córdova (2005) and Sandoval et al. (2007) reported increases in the content of SOC and microbial biomass and its activity. This was the result of a lower intensity of agricultural land use that included an 8 yr crop rotation (3 yr of annual crops and 5 yr hay), compared to a conventional 4 yr crop rotation (continuous annual crops). Rossel and Galantini (2006) estimated the potential of C sequestration in soils of the western mountain ridges and desert of Argentina indicating that SOC content can improve by converting degraded lands to forest or permanent grasses, with a potential increase of 420 ± 140 Tg (1 Tg = 1 x 10 12 g) of SOC in a total of 140 Mha. In this sense, Lal (2006) show relevant information about estimations and alternatives for increase the soil carbon sequestration in different biomass of Latin America. Many soil properties are related to soil structure, where the macro- and micro-aggregates confer resistance to the microbial and enzymatic decomposition of C. This involves the occlusion of the organic material in structural units of the soil, preventing enzyme attacks or microorganism access to the area of the organic material, and a reduced diffusion of oxygen to the interior of the aggregates, which reduces the microbial activity (Christensen, 2001). The change in LU and management practices affected the stability of aggregates, C content and its dynamics in Andisols of South-Central Chile (40º S lat) (Huygens et al., 2005) and the implementation of a long-term pasture, as part of crop rotations in volcanic soils of Chile (Sandoval et al., 2007), favored the accumulation of specific carbonaceous molecular species that increased the proportion of stable macro-aggregates and soil C sequestration. Tisdall and Oades (1982) proposed the concept of soil aggregate hierarchy, in which the primary particles of soil aggregates link together and are stabilized by persistent agents such as humic substances or complexes with polyvalent cations that form microaggregates (less than 250 μm), and through temporary linking agents, such as polysaccharides, fungal hyphae and plant roots that contribute to the formation of macroaggregates (upper than 250 μm). Among the persisting linking agents, calcium (Ca +2 ) in the soil promotes the formation of clay-cation polyvalent-organic material complexes (Clough and Skjemstad, 2000). Furthermore, it indirectly influences the formation of macroaggregates through the stimulation of microbial activity in acidic soils (Chan and Heenan, 1999). Additions of Ca +2 (as carbonaceous compounds) in field studies increased the aggregation level for approximately 10% (Chan and Heenan, 1998; 1999). However, a temporary decrease (1 to 3%) in aggregate stability has been observed in soils with variable charge, due to an increase in the positive charges of the surfaces exposed to mineral soils that produce the dominance of repulsive forces and cause clay dispersion. Continuous and long-term applications of Ca +2 favor the formation of stable soil aggregates (Chan and Heenan, 1998; 1999). It is also important to consider that sesquioxides influence the stability of soil organic matter (SOM) and aggregation in different soils. Barral et al. (1998) indicated that aluminium (Al 3+ ) and iron (Fe 3+ ) control aggregation in acidic soils with low clay and SOC contents, such as Oxisols. Barthès et al. (2008) suggested that, in some tropical soils, aluminium containing sesquioxides were major determinants of macroaggregate and OM stability. Finally, Matus et al. (2006) after a compilation of 225 soil pedons from two nationwide datasets of South-central Chile indicate that aluminium is the principal factor for immobilization of SOC in acid volcanic soils. Mitigation of N2O emissions from soils Different and complementary strategies may be used to reduce N2O emissions from agricultural soils. Their successful implementation by farmers depends on costs, soils, crops, markets and climate conditions. Saggar et al. (2009) indicate that current strategies to reduce N2O are soil management, optimum N application rate, timing of application and use of nitrification inhibitors, among other measures. For example, management of organic or inorganic fertilizers directly influences N2O emissions. Saggar et al. (2009) compiled information from several countries and determined that the use of animal urine and dung or grazed grasslands imply losses from 0 to 4% of N input as N2O, but it was found that in poorly drained clay soils of England losses can be higher (11 to 14% of N input). On the contrary, the use of N fertilizers in grassland soils can produce 0 to 2% of N input as N2O, but poorly drained sandy soils of Scotland have reported losses of up to 12% of N input. An extensive review about management options to reduce N2O emissions from grazed pastures is available in Luo et al. (2010). Nevertheless, Rochette et al. (2008) reported that fertilization of silage maize with dairy cattle manure resulted in greater or equal N2O emissions when compared with the use of synthetic N. Different results have been reported regarding the influence of tillage managements on N2O flux. Rochette (2008) compiled information from 25 studies (approximately 45 site-years of data) with same-site comparisons of no-till and tilled soils, showed that no-till generally increased N2O emissions in poorly-aerated soils in regions with a humid climate, but was neutral in soils with good and medium aeration. On average, N2O emissions under no-till were 0.06 kg N ha -1 lower, 0.12 kg N ha -1 higher and 2 kg N ha -1 higher than under tilled soils with good, medium and poor aeration, respectively. In addition, Ball et al. (2008) found that fluxes of N2O were 1.5 to 35 times higher from no-till than from ploughed soils, indicating that the N2O emission were strongly influenced by the soil water-filled pore space. In addition, Bhatia et al. (2010) determined that the temporal emissions of N2O-N were higher in no-till versus conventional till system since soils under no-tillage were generally more moist and had organic matter concentrated near the soil surface, what favored N2O production. Another strategy for limiting N2O emissions from fertilizers is the use of chemical inhibitors of nitrification that reduce the oxidation of NH4+ to NO3 - , or inhibitors of urease activity that inhibit or slow down the release of urea-N. Bremner and Douglas (1971) studied the effects of more than 100 compounds (organic and inorganic) on soil urease activity and concluded that 2,5-dimethyl-p-benzoquinone, 2,5-dichloro-p-benzoquinone, and 2,6-dichloro-p-benzoquinone were the most effective organic compounds to slow down urea hydrolysis in soils and, consequently, reduce agronomic and environmental problems caused by the fast hydrolysis of urea. At commercial level, it is possible to find nitrification inhibitors such as nitrapyrin (2-chloro-6-trichloromethylpyridine; NP), dicyandiamide (2-cyanoguanidine; DCD) and 3,4-dimethylpyrazole phosphate (DMPP) (Carrasco and Villar, 2001) with diverse results. For example, incorporation of NP with N fertilizers (i.e., urea) in cereal crop productions increased the efficiency of N use from 38 to 74% on oat, and from 59 to 92% under corn crops in volcanic soils of Chile (Longeri et al., 1988), but NP and thiourea applied to spring wheat resulted in a reduction in grain production and harvest index (Vidal and Longeri, 1988). The use of DCD in pastures has shown reductions greater than 80% in N2O emissions (Mosier et al., 1998). On the contrary, the same inhibitor applied to very acidic soils (pH 4) was not effective in reducing N2O emissions, probably due to the inactivation by association with SOM (Mosier et al., 1998). Acetylene has shown to be a strong inhibitor, but it is difficult to apply and maintain adequate concentrations in the soil (Freney et al., 2000). In recent reports, Bhatia et al. (2010) applied new nitrification inhibitors, S-benzylisothiouronium butanoate (SBT-butanoate) and S-benzylisothiouronium furoate (SBT-furoate), and obtained reductions of the N2O emission of 15.6 and 19.2% in a no till system. An alternative strategy is the use of the allopathic actions of some plant species on nitrifying and denitrifying microorganisms. Many authors (e.g. Bremner and Douglas, 1971; Wardle et al., 1997; Lata et al., 2004) have studied plant species from a wide range of ecosystems and have indicated that the inhibitory effect of the nitrification and/or denitrification processes is an adaptive process of certain plant species to low N content in the soil, as a mechanism of nutrient conservation. In this sense, Lata et al. (2004) indicate two possible mechanisms involved in allelopatic effect of Hyparrhenia diplandra Hack. (Poaceae) on nitrifying bacteria in a wet savanna ecosystem in Western Africa, first through the root exudation of phenolic acids and tannins compounds and the competition for the available ammonium. In conifer forests of the Northern Hemisphere, adapted to occurrence of natural fires, the removal of Ericaceae species, that compose an intermediate phases of post fire succession, promotes a significant increase in soil nitrification, due to a decrease of the amount of phenols and terpenes in the soil (Wardle et al., 1997). By conducting a pot experiment, Kiran and Patra (2003) determined that applications of two doses (100 and 200 mg kg -1 ) of urea, coating with dried plant materials of Mentha spicata L. and Artemisia annua L. increased the essential oil yields of menthol mint, indicating that these natural products could be potential nitrification inhibitors, Conclusions Agricultural and forestry activities are potential sources of GHG emissions to the atmosphere produced as result of soil biological processes. More knowledge on the biological processes that promote GHG emissions from soil, as well as on their relation with different types of soil management and use, will allow to the creation of new opportunities for agricultural development under friendly environmental conditions. Soil ecosystems act as absorbers, reservoirs and emitters of GHG, depending on the balance of inputs and outputs, which are conditioned by different processes that influence GHG emissions such as soil biological respiration, rate of nitrification and other oxidative process as soil erosion and land use change. Mitigation measures to reduce net GHG emissions include increasing soil C sequestration by reduced tillage, extended rotations, increased crop use efficiency of fertilizer-N, use of chemical or natural inhibitors of nitrification. Adequate management of soils and crops results in a reduction of GHG emissions by storing atmospheric C as soil organic matter. Acknowledgements The authors thank the Bicentenary Program of Science and Technology, CONICYT-World Bank-Universidad de Concepción "Inserción de Investigadores a la Academia (PSD-12)", FONDECYT Project N° 1080076, DIUC N° 208.123.010-1.0, and Livestock Emissions & Abatement Research Network (LEARN) Post-doctoral Fellowships for their support that make possible our research. [95] References
Copyright 2010 - Chilean Journal of Agricultural Research The following images related to this document are available:Photo images[cj10053f1.jpg] [cj10053t2.jpg] [cj10053t3.jpg] [cj10053t1.jpg] [cj10053f2.jpg] |
| |||||||||

![[Table - 1]](/showimage?cj/photo/cj10053t1.jpg){kind=link}
![[Figure - 1]](/showimage?cj/photo/cj10053f1.jpg){kind=link}
![[Table - 2]](/showimage?cj/photo/cj10053t2.jpg){kind=link}
![[Table - 3]](/showimage?cj/photo/cj10053t3.jpg){kind=link}