 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Chilean Journal of Agricultural Research, Vol. 71, No. 1, 2011, pp. 52-62 RESEARCH Induction of resistance in chickpea (Cicer arietinum L.) against Ascochyta rabiei by applying chemicals and plant extracts Inducción de resistencia en garbanzo (Cicer arietinum L.) contra Ascochyta rabiei por la aplicación de productos químicos y extractos vegetales. M. Usman Ghazanfar1*, Waqas Wakil2, and Shahbaz Talib Sahi3
[1]
University of Sargodha, University College of
Agriculture, Sargodha, Pakistan. *Corresponding author (usmanghazanfar1073@yahoo.com). Received:
20 August 2010 Code Number: cj11007 ABSTRACT Since new fungicides are scarce in the market and because of environmental problems, researchers are now emphasizing other alternatives, such as the genetic potential of plants, resistance against pathogens, and the use of biotic and abiotic agents to develop induced or acquired resistance. We investigated the role of resistance-inducing substances (chemicals and plant extracts) in three chickpea cvs. C-44, Pb-91, and Bittle-98 in field experiments against Ascochyta blight disease. These cultivars were selected on the basis of better yield potential shown in previous experiments. Aqueous solution of salicylic acid (SA) at 0.5, 1.0, and 1.5 mM; acibenzolar-S-methyl (Bion®, ASM) at 0.4, 0.8, and 1.2 mM; and KOH at 25, 50, and 75 mM were applied, whereas Azadirachta indica A. Juss., Datura metel L., and Allium sativum L. plant extracts were applied at 5, 10, and 15%. Disease reduction data was recorded at different intervals from 4 to 14 d after being induced and inoculated with the pathogen. Overall results revealed that significant disease reduction (79.3%) was provided by ASM in the cv. C-44 at 1.2 mM compared with SA, whereas the least significant was KOH. Maximum disease reduction (43.5%) against disease in the plant extracts was observed by applying A. indica leaf extract, but D. metel and A. sativum extracts were not effective. Our findings suggest that enhancing resistance before infecting chickpea plants could be an innovative control method for Ascochyta blight of chickpea. Key words: Induced resistance, acibenzolar-S-methyl, salicylic acid, Azadirachta indica, Datura metel. RESUMEN Debido a la escasez de nuevos fungicidas en el mercado y problemas ambientales, los investigadores están ahora enfatizando otras alternativas, tales como potencial genético de las plantas, resistencia contra patógenos y uso de agentes bióticos y abióticos para el desarrollo de resistencia inducida y adquirida. Investigamos el rol de sustancias inductoras de resistencia (químicos y extractos vegetales) en tres cultivares de garbanzo ‘C-44’, ‘Pb-91’, ‘Bittle-98’, en experimentos de campo contra tizón por Ascochyta. Estos cultivares se seleccionaron basado en el mejor potencial de rendimiento mostrado en experimentos previos. Solución acuosa de ácido salicílico (SA) a 0,5; 1,0; y 1,5 mM, acibenzolar-S-metil (BionÒ) a 0,4; 0,8; y 1,2 mM y KOH a 25, 50, y 75 mM se aplicaron mientras los extractos vegetales de Azadirachta indica A. Juss., Datura metel L. y Allium sativum L. se aplicaron a 5, 10, y 15%. Datos de reducción en la enfermedad se registraron con diferentes intervalos desde 4 a 14 d después de inducción e inoculación con el patógeno. Los resultados revelaron que ASM 1,2 mM provocó una reducción significativa de la enfermedad (79,3%) en el cv. C-44 comparado con SA; sin embargo, la menor reducción fue observada para KOH. Entre los extractos vegetales, la máxima reducción de la enfermedad (43,5%) se observó con la aplicación de extracto foliar de A. indica y los extractos de D. metel y A. sativum no fueron efectivos para la reducción de la enfermedad. Los presentes hallazgos sugieren que el aumento de la resistencia antes de la infección de plantas de garbanzo pudo ser un método de control innovador para tizón del garbanzo por Ascochyta. Palabras clave: Resistencia inducida, acibenzolar-S-metil, ácido salicílico, Azadirachta indica, Datura metel. INTRODUCTION Chickpea blight caused by Ascochyta rabiei (Pass.) Labrousse is the most devastating chickpea (Cicer arietinum L.) foliar disease in many countries (Pande et al., 2005). This disease has caused considerable losses ranging from 5 to 100% around the world (Haware, 1998). In Pakistan, the number of epiphytotics has been reported with complete crop failure (Aslam, 1984). Chickpea breeders in Pakistan are focusing their attention on developing blight resistant cultivars (Iqbal et al., 1993; Ilyas et al., 2007). Different aspects of this disease, such as severity, lifecycle, pathogenicity, epidemiology, breeding for resistance, cultural, and chemical control measures have been successfully explored in different parts of the world, although this disease still poses a great threat to the chickpea crop (Mohamed et al., 2009; Trapero-Casas and Kaiser, 2009). This disease can be managed with different fungicides (Gan et al., 2006), but the most economical management strategy is using resistant cultivars (Türkkan and Dolar, 2009). However, breeding resistant chickpea cultivars is more difficult because of the great variation in A. rabiei isolate pathogenicity (Ali et al., 2009). Since the pathogen’s nature is constantly changing (Chaudhry et al., 2001), already resistant cultivars become susceptible. Therefore, what is needed to restrain or alleviate infection are recent management methods such as induction of resistance, which is a new technology for plant disease control based on activating the plant’s own defense system with the help of low molecular weight synthetic molecules (Cohen et al., 1999). Induced resistance is defined as an increase in the level of resistance without any changes in the basic genetic constitution (Baysal, 2001), which is associated with an enhanced ability of the plant to resist infection by an intense activation of defense responses. This method is the alternative to fungicides to protect plants since there is greater awareness about the harmful effect of fungicides on the natural ecosystem (Vimala and Suriachandraselvan, 2009). At the same time, development of pathogen resistance to pesticides, real and perceived health and environmental concerns increase the popularity of organic crops and sustainable agriculture (Kuć, 2001). Induced resistance can appear locally (i.e., in the organ where resistance is applied), or it may be systemic to the plant part which is spatially separated from the site of inducing agents (Walter, 2009). There are at least three types of induced resistance: systemic acquired resistance (SAR) developed by localized necrosis resulting in hypersensitive reaction (HSR), wound-induced resistance usually caused by insect feeding, and infection by virulent pathogen or by treatment with certain chemically induced systemic resistance (ISR) that developed due to plant growth colonization promoting rhizobacteria (PGPR).In systemic acquired resistance, an invading pathogen is not needed and various defense pathways are stimulated in the induced plant resulting in the production of diverse defense products, including lignin and pathogenesis-related proteins, some of which show chitinase or β-1,3-glucanase activity (van Loon, 1997; Neuhaus, 1999), phytoalexins, thionins, and defensins (Kessmann et al., 1994; Sticher et al., 1997; Kombrink and Schmelzer, 2001; Ziadi et al., 2001). Induced resistance is an age-old phenomenon for managing plant diseases that was first reported by Ray (1901) in rust diseases. Numerous studies have shown the appearance of defense responses against several important plant diseases (Abo-Elyousr and El-Hendawy, 2008; Jiang et al., 2008). Various chemicals, such as salicylic acid, isonicotinic acid (INA), benzothiadiazole (BTH), Rigel (salicylic acid analog), β-aminobutyric acid (BABA), NaClO3, HgCl2, probenazole, paraquat, polyacrylic acid, SiO2, Messenger (Harpin protein), and Phoenix (potassium phosphate) have been used to induce resistance against fungi, bacteria, and viruses (Schneider et al., 1996; Kuć, 2001; Percival et al., 2009). Extracts of various plants have also been explored as natural resistance inducers such as Azadirachta indica Juss. against Alternaria leaf spot on sesame (Guleria and Kumar, 2006), Datura metel L. against Rhizoctonia solani, Xanthomonas oryzae pv. oryzae, and Alternaria solani (Kagale et al., 2004; Latha et al., 2009). Few studies have been carried out on induction of resistance in Ascochyta blight of chickpea, except Chaudhry et al. (2001) who studied the induction mechanism in susceptible chickpea cv. C-727 by applying an aqueous solution of salicylic acid dipotassium hydrogen phosphate and cuprous chloride and found that all treatments showed significant results. The objective of our studies was to evaluate the reduction in Ascochyta blight disease as a result of induced resistance by applying simple chemicals and plant extracts. MATERIALS AND METHODS Chickpea cvs. C-44, Pb-91 and Bittle-98, which were susceptible but had shown good yield character in a previous experiment, seeds were cultivated in small plots of 6.006 m2 in the experimental area of the Department of Plant Pathology, University of Agriculture, Faisalabad, Pakistan during the 2007-2008 season. The experiment was laid out under factorial arrangement of randomized complete block design with three replicates of 39 plots. Eighteen plots were induced and inoculated, three plots served as controls while the remaining 18 plots were induced but not inoculated. Each plot had six rows of 15 chickpea plants. Extracts of neem (Azadirachta indica), Datura (Datura metel L.), and garlic (Allium sativum L.) were prepared in the phytobacteriology laboratory of the Department of Plant Pathology, University of Agriculture, Faisalabad. Neem and datura leaves were collected from the agronomic research area and square N° 32, University of Agriculture, Faisalabad while garlic cloves were purchased from a local market. Neem and datura leaves were thoroughly washed under tap water, materials were surface-sterilized with 1% sodium hypochlorite solution, and then washed with sterilized water. Leaves were homogenized (Yellow line DI-25 Basic, GmbH and CO, Germany) in sterile distilled water at 1:1 (w/v) and filtered through muslin cloth to produce a 100% crude plant extract. Garlic extract was prepared from fresh samples; the dry outer peel of the cloves was first removed, surface-sterilized for 2 min in 70% ethanol, and washed in three changes of sterile distilled water. Cloves were crushed into a pulp in a sterile porcelain mortar with a pestle; pulp was suspended in 100 mL water in a 250-mL Erlenmeyer flask, and filtered through muslin cloth. Plant extracts were heated to 40 °C for 10 min to avoid contamination (Jaganathan and Narasimhan, 1988) and diluted to the required concentration (5%, 10%, and 15%) with sterile distilled water (v/v). Salicylic acid (Sigma Aldrich, Germany) and KOH (Sigma Aldrich, Germany), purchased from the market, and Bion® (acibenzolar-S-methyl), provided by Syngenta Crop Protection, Germany, were the chemicals used to induce resistance. Aqueous solution of salicylic acid (SA) 0.5, 1.0, and 1.5 mM; acibenzolar-S-methyl (ASM) 0.4, 0.8, and 1.2 mM; and KOH 25, 50, and 75 mM were applied. However, all plant extracts were applied at 5%, 10%, and 15%, respectively. At the early flowering stage, all the resistance inducing agents (chemicals and plant extracts) were sprayed onto the plants until runoff while control plants were sprayed with distilled water. Mass preparation of already isolated and preserved A. rabiei inoculums was carried out by the method of Ilyas and Khan (1986). Conidial counts were adjusted with a haemocytometer. Normal agronomic practices were followed throughout the experiment. Four days after induction of resistance, plants were challenged with spore suspension of A. rabiei (1 x 105 spores L-1) after adding three drops Tween 80 per liter as a wetting agent with a lady hand sprayer until runoff in the evening since temperatures are lower at night helping better conidia germination; this was continued for 3 d to ensure maximum plant infection. Plants were sprinkled with water to maintain the humidity level for the conidial germination of the fungus. Data on disease reduction percentage was calculated at a 4, 6, 7, 11, and 14 d interval by a mixed quantitative and qualitative 0-9 rating scale and converted to disease percentage reduction by the slightly modified formula from Tivoli et al. (2006):
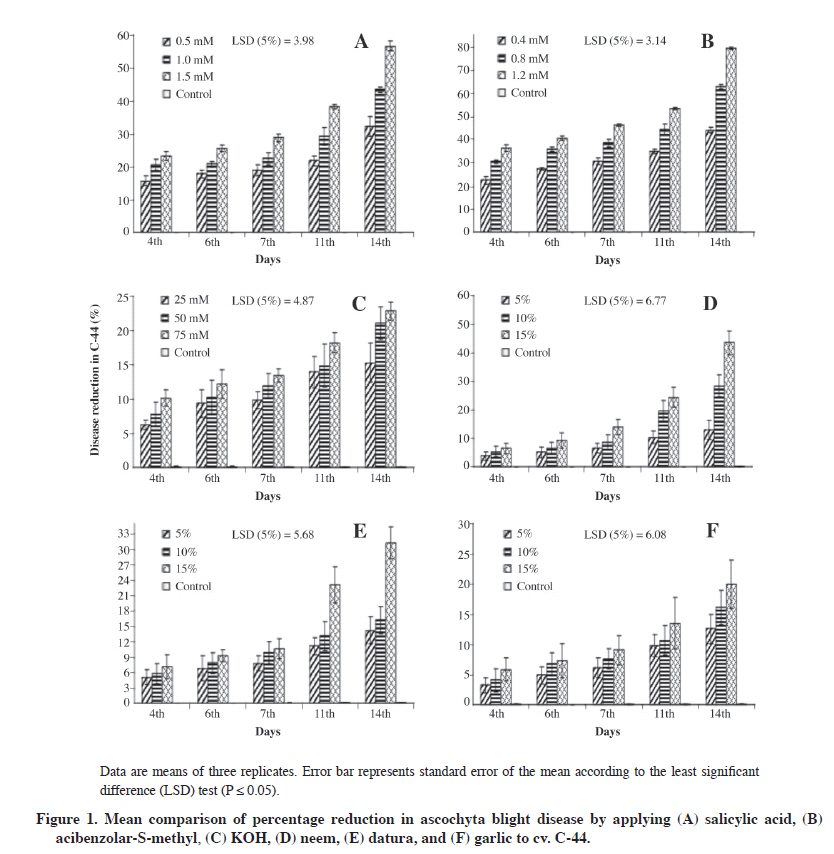
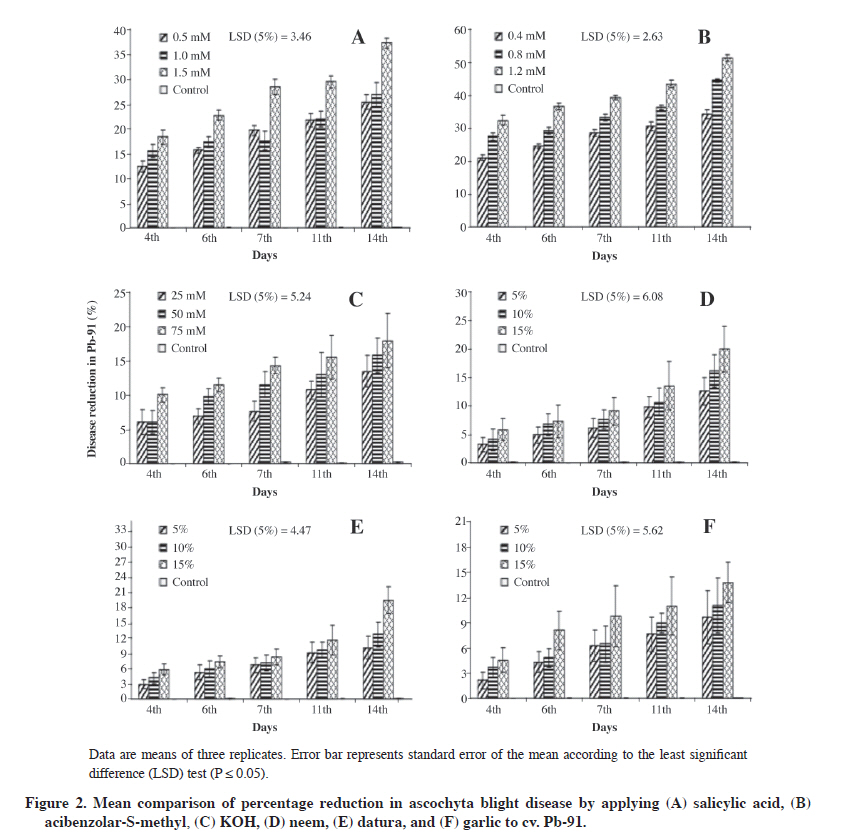
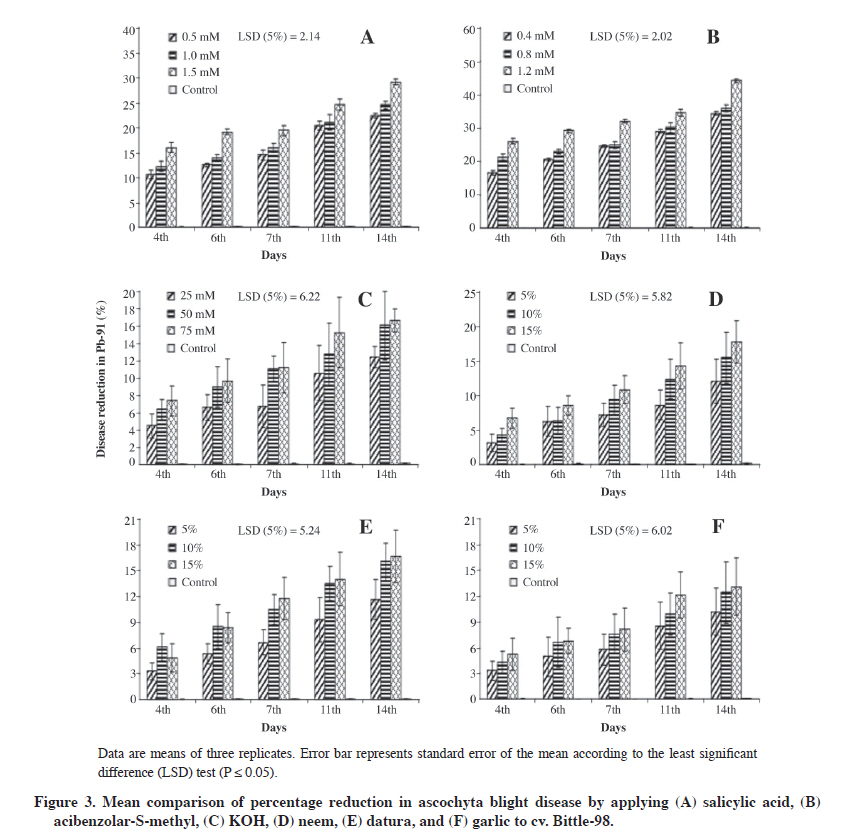
Data were subjected to ANOVA and the significance of treatments was determined by Duncan’s multiple range test (P ≤ 0.05) (Gomez and Gomez, 1984; Steel et al., 1997). RESULTS Percentage disease reduction by applying chemicals and plant extracts to ‘C-44’ Disease reduction was highly significant in Ascochyta rabiei (P ≤ 0.05) and observed through pretreatment with different chemical and plant extract doses in cv. C-44 (Figure 1). Maximum efficacy after inoculation with the pathogen was in the aqueous solution of ASM (79.3%) while the lowest reduction was exhibited by KOH (22.8%). Applying the aqueous solution of SA revealed a 56.8% disease reduction. Among the various tested concentrations of SA, ASM, and KOH, the highest dose rates, (1.5, 1.2, and 75 mM respectively) of these resistance inducing substances were found to be effective against pathogen development, and effectiveness persisted for 14 d after the induction and inoculation with the pathogen although at all the applied chemical dose rates, significant protection started after day 11 and reached the maximum after 2 wk. The three plant extracts were evaluated at 5, 10, and 15% dose rates and neem leaf extract efficiently suppressed the disease (43.5% reduction) while the datura (31.4%) and garlic (26.7%) extracts also demonstrated protection, although lower than neem (Figure 1). Furthermore, this protection persisted for 2 wk and the highest concentration was efficient in reducing the disease. Percentage disease reduction by applying chemicals and plant extracts to ‘Pb-91’ Applied plant extracts (5, 10, and 15%) in cv. Pb-91 significantly reduced disease development (P ≤ 0.05). All extracts equally provided reduction 14 d after inoculation against Ascochyta rabiei under field conditions with neem (20.0%), datura (19.5%), and garlic (13.8%) (Figure 2). Results for chemicals were significantly different from plant extracts although low disease reduction occurred with SA and ASM (37.2% and 51.2%) compared with cv. C-44, which was quite promising for controlling chickpea blight. The lowest disease reduction in KOH (17.9%) was similar to plant extracts. Percentage disease reduction by applying chemicals and plant extracts to ‘Bittle-98’ The percentage of disease reduction in cv. Bittle-98 by applying chemical and plant extracts was significant (P ≤ 0.05). Applying chemicals to induce resistance was more efficient than with plant extracts. The highest level of disease reduction was ASM (44.1%) after 14 d with a 1.2 mM concentration (Figure 3). KOH (16.6%) had the lowest disease reduction while SA effectively reduced ascochyta blight (29.1%). Neem (17.7%), datura (16.6%), and garlic (13.1%) exhibited disease reduction. The effect of all these plant extracts and KOH is similar, but ASM and SA were superior in protecting chickpea against A. rabiei. DISCUSSION Induced resistance has attracted the attention of researchers worldwide as a possible strategy for integrated plant disease management (Sundar et al., 2001). The curative property of SA and its derivatives has been known since Hippocrates described the presence of salicylates to provide relief during childbirth (Weissman, 1991). Significant knowledge advances have been made in the last 20 yr about the metabolism and signaling mechanism in plant defense activation when applying SA (Durner et al., 1997). The role of SA is well-known at the beginning of defense responses against different abiotic and biotic stress (Catinot et al., 2008). Results have proved that SA can be used to control chickpea blight. Similar reductions (49.5%) against A. rabiei were achieved (Chaudhry et al., 2001) by applying SA at 1 mM in the susceptible cv. C-727. The effect of SA to induce resistance in groundnut plants against Alternaria alternata was investigated by Chitra et al. (2008). Foliar application of SA at 1 mM significantly reduced leaf blight disease intensity and increased pod yield under glasshouse conditions. Foliage spray of 2 mM SA significantly reduced (40.3%) leaf blight of onion caused by Stemphylium vesicarium under greenhouse conditions 15 d after inoculation (Abo-Elyousr et al., 2009). The ability of the four chemicals SA, sodium salt of SA, isonicotinic acid, and DL-β-amino-n-butyric acid, as well as the yeast antagonist Cryptococcus flavescens were evaluated against Fusarium head blight (FHB) of wheat under greenhouse conditions showing that NaSA and 2,6-dichloroisonicotinic acid (INA) at 10 mM significantly reduced FHB severity compared with the non-treated disease control 3 d prior to challenge with the pathogen (Zhang et al., 2007). The significant trait of the disease resistance inducer is that it persists longer in the host by increasing its biochemical constituents as in this study where SA is still effective against A. rabiei on day 14. Applying a higher concentration of SA at 8 g L-1 caused more vigorous growth than in the control (Quintanilla and Brishammar, 1998) indicating that increased resistance was obtained in the tuber injected with a higher concentration of SA. After being infected with TMV, a higher accumulation of SA at the infection site was also studied, which may be attributable to transcriptional activation of PR genes in the inoculated and uninoculated leaves (Durner et al., 1997), but this increase was not significant in the uninfected systemic tissue. Kone et al. (2009) studied the induction of disease resistance to Phytophthora capsici on squash by applying DL-3-aminobutyric acid (BABA), 2,6-dichloro-isonicotinic acid (INA), Saver (ai: salicylic acid), Syrup (nutrient supplement), and ASM on the mycelial growth in vitro studies and found that only Saver significantly reduced mycelial growth and sporangium production at concentrations of 100 mg mL-1 or higher. Bion (acibenzolar-S-methyl) was originally marketed to manage powdery mildew of wheat and barley in Europe (Görlach et al., 1996). ASM and INA are considered the best chemical elicitors available to induce resistance. They are considered functional analogs of SA and bring out a systemic form of induced resistance across a broad range of plant pathogens (Friedrich et al., 1996; Maleck et al., 2000). These chemicals did not exhibit any direct antimicrobial activity; however, some instances of antimicrobial activity have been reported in association with high elicitor concentrations (Tosi and Zazzerini, 2000; Rohilla et al., 2002). In our studies, ASM efficiently reduced A. rabiei disease and the reduction lasted for 2 wk using the highest concentration. Similar results were obtained by Dann et al. (1998) who studied field and greenhouse experiments from 1993 to 1996 and found that a 20 to 70% reduction in severity was recorded for three or four applications of 2,6-dichloroisonicotinic acid (DCINA) in soybean cvs. Elgin 87 and Williams 82, which were highly susceptible to Sclerotinia sclerotiorum. This effect was not evident in cvs. Corsoy 79 and NKS19-90, which were resistant to white mold disease. Yield also increased, particularly in susceptible cultivars under high disease pressure. Comparable results were also reported by Sundar et al. (2001) as significantly reducing red rot lesion development in sugarcane pretreatment with CGA-245704 at 100 µg mL-1 applied as soil drench to the potted plants, and effectiveness persisted for up to 30 d after the treatment. Persistent resistance for longer periods was also reported by Ziadi et al. (2001) against downy mildew of cauliflower. Dann and Deverall (1996) demonstrated in two consecutive years of field trials that applying 2,6-dichloro-isonicotinic acid (INA) to 16- to 20-d-old bean seedlings (cv. Redlands Pioneer) were protected against rust disease caused by inoculation with Uromyces appendiculatus. This protection persisted for 5 wk and the spread of the disease was curtailed to the upper uninoculated leaves so that there was no need to apply further booster doses of INA. Sometimes plants exposed to higher concentrations have also shown phytotoxicity (Louws et al., 2001), but independently of induced resistance. Lopez and Lucas (2002) studied the ability of different resistance inducing chemicals to provide protection against anthracnose of cashew caused by Colletotrichum gloeosporioides. Maximum disease reduction in detached leaves with no phytotoxic effects was obtained with 0.07 mM a.i. ASM and DCINA, 5 mM SA, and 50 mM K2HPO4 with a time interval of at least 72 h between activator applications and inoculation with the pathogen. In the case of attached leaves, foliar spray was more efficient than the soil drench treatment. Efficacy of ASM and INA has also been reported in monocots powdery mildew, Septoria leaf spot, and rust of wheat in field trials and solanaceous crops. Effectiveness of BTH applied at 35 g a.i. ha-1 at a 7 to 10 d interval gave promising results to control Xanthomonas axonopodis pv. vesicatoria and Pseudomonas syringae pv. tomato on tomato foliage and fruits, leguminous crops, and white mold of soybean can be controlled by repeated applications of INA, on fruit trees, cotton, and spinach (Vallad and Goodman, 2004). Contrary results of dose-dependent response in ASM were studied by Ajay and Baby (2009) against blister blight disease of tea and showed that applying ASM at a 0.14% dose rate exhibited 25% disease protection compared with a 0.18% (not significant). Although a substantial number of studies have dealt with phosphate-induced resistance in plants, the primary mode of phosphate action is unknown. It is believed that activation of SAR by locally applying phosphate strongly suggests that it could result either from a translocation of the inducer itself or from signals which initiate an oxidative burst that are generated in phosphate-treated tissues. Applying KOH in this study was not significant except in cv. C-44 with 23% disease reduction, which is not considered as satisfactory in any disease control program. Different types of phosphate salt viz dibasic and tribasic sodium or potassium phosphates were highly effective in SAR induction with a pH value greater than 7 (Gottstein and Kuć, 1989). Foliar application of these phosphate salts induced systemic resistance (ISR) in cucumber (Descalzo et al., 1990), broad bean (Walters and Murray, 1992), grapevine (Reuveni and Reuveni, 1995), maize (Reuveni et al., 1994), pepper (Reuveni and Reuveni, 1995; Reuveni and Reuveni, 1998), and rice (Manandhar et al., 1998) against diversified pathogens belonging to various taxonomic groups. However, all the results are contradictory with our findings and there might be a difference in the selection of K salts or pH may be less than 7. On the other hand, applying 50 mM dipotassium hydrogen phosphate provides 51.5% protection against A. rabiei (Chaudhry et al., 2001). The efficacy of various plant extracts to induce resistance was studied in these experiments; overall results showed that only neem leaf extract at the highest dose rates can provide protection against chickpea blight in cv. C-44 while the effectiveness of datura extracts and garlic is not significant. This lower disease reduction might be due to the lower dose of these plant extracts and perhaps a booster dose was needed but not applied so that these results are from a single application.. The role of plant extracts to induce resistance against plant pathogens has been reported by different researchers. Guleria and Kumar (2006) found that aqueous leaf extract of neem controlled Alternaria leaf spot of sesame. The efficacy of neem leaf extract has also been reported by Paul and Sharma (2002) and it had the same effect as bavistin in managing leaf stripe pathogen of barley. Antimicrobial activity and induction of systemic resistance by Datura metal leaf extract against Rhizoctonia solani and Xanthomonas oryzae pv. oryzae in rice has been investigated (Kagale et al., 2004) and it was found that foliar application of leaf extract effectively reduced the incidence of sheath blight and bacterial blight of rice. Satya et al. (2007) applied aqueous leaf extract of zimmu (Allium sativum L. x Allium cepa L.) to first and second leaves cotton plants that induced systemic resistance in third and fourth leaves and reduced the number of lesions by up to 73% after challenged infection with Xanthomonas campestris pv. malvacearum compared with water-treated control plants. This might be due to the fact that active ingredients in these plants can enhance biochemical substances when applied as allicin extract from A. sativum and oxalate as active components of spinach and rhubarb leaves, which can reduce plant diseases by accumulated SA (Doubrava et al., 1988; Slusarenko et al., 2008). It has been effectively used against different plant pathogens, such as Alternaria solani, Pseudomonas syringae pv. tomato (Pst), Xanthomonas vesicatoria (Xv), and Clavibacter michiganensis sub-sp. michiganensis (Cmm) of tomato and induce disease resistance (Balestra et al., 2009; Latha et al., 2009). Roth et al. (2000) studied the effects of an extract of Lychnis viscaria L. seeds that contains brassinosteroids, an aqueous application enhanced by 36% resistance to tobacco, cucumber, and tomato against viral and fungal pathogens by increasing PR-proteins, peroxidase, chitinase, and β-1,3-glucanase. The disease-controlling ability of Datura metel against plant pathogens and as an inducer of resistance is well documented in the literature as A. rabiei (Shafique and Shafique, 2008), Pennisetum glaucum, and Sclerospora graminicola in pearl Millet (Devaiah et al., 2009). CONCLUSION We concluded that resistance inducing chemicals have the potential to control chickpea blight when applied at the proper time and dose rate. These chemicals can be integrated with suitable fungicides since they have a synergistic effect with fungicides and persist longer; it would help reduce the number of fungicide applications by enhancing the natural resistance of chickpea cultivars even if they were susceptible to A. rabiei. Plant extracts can be used, but at higher dose rates and combined with resistance inducing chemicals to control chickpea blight. ACKNOWLEDGEMENTS The researcher is highly obliged to Dr. Shakeel Ahmad, Head of Research and Development Syngenta, Pakistan for providing Bion and his assistance to import it from Germany. LITERATURE CITED
Copyright 2011 - Chilean Journal of Agricultural Research The following images related to this document are available:Photo images[cj11007f2.jpg] [cj11007f3.jpg] [cj11007f1.jpg] |
| |||||||||

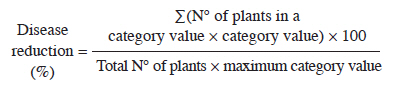
{kind=link}
{kind=link}
{kind=link}