 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Chilean Journal of Agricultural Research, Vol. 71, No. 1, 2011, pp. 63-72 RESEARCH Symptom severity and viral protein or RNA accumulation in lettuce affected by big-vein disease Severidad de síntomas y acumulación de proteínas o ARN virales en lechugas afectadas por la enfermedad de las venas grandes Carolina Araya1, Elizabeth Peña1, Erika Salazar1, Lisset Román1, Claudia Medina1, Roxana Mora1, Agustín Aljaro1, and Inés-Marlene Rosales1 1Instituto de Investigaciones
Agropecuarias INIA, Casilla 439-3, Santiago, Chile Received:
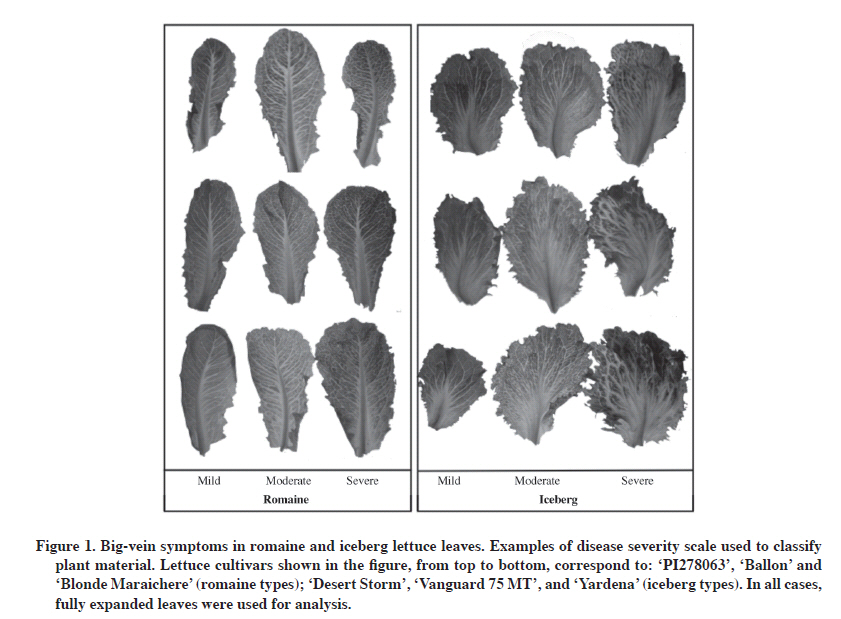
07 December 2009 Code Number: cj11008 ABSTRACT Big-vein disease (BVD) is a widespread and economically damaging disease in lettuce (Lactuca sativa L.). Typical symptoms are chlorotic clearing around leaf veins, leaf deformations, and impaired head development. In this research, we studied the relationship between symptom intensity and protein and viral RNA accumulation in infected plants. Naturally infected lettuce plants, from the field or greenhouse, were classified according to their symptomatology: mild, moderate, severe, and symptomless. Coat protein accumulation was evaluated by a double antibody sandwich/enzyme-linked immunosorbent assay (DAS-ELISA), and RNA levels were studied by semi-quantitative reverse transcription polymerase chain reaction (RT-PCR) and quantitative RT-PCR. Virus coat protein accumulation did not differ for the two viruses associated with this disease among lettuce plants showing different symptom severity. Similarly, abundance of Mirafiori lettuce big-vein virus (MLBVV)-RNA3 or Lettuce big-vein associated virus (LBVaV)-RNA-2 were not different (P > 0.05) for diverse big vein disease severity rating scales. This suggests that symptom severity expressed by big-vein diseased lettuce plants did not necessarily reflect the accumulation of viruses associated with this disease in the host. Therefore, lettuce plants showing mild symptoms of BVD do not necessarily present lower virus levels than plants showing more severe symptomatology. Key words: Lettuce big-vein disease, symptom expression, Ophiovirus, Varicosavirus, Lactuca sativa. RESUMEN La enfermedad de las venas grandes de la lechuga (Lactuca sativa L.) es de origen viral, está ampliamente distribuida en el mundo, y provoca graves daños económicos en este cultivo. Los síntomas típicos de la enfermedad son clorosis alrededor de las venas, deformación de hojas y ausencia de formación de cabezas. En este trabajo se estudió la relación entre la intensidad de síntomas y la acumulación de proteínas y ARNs de origen viral en plantas afectadas por esta enfermedad. Lechugas infectadas naturalmente, provenientes de campo y de invernadero fueron clasificadas con sintomatología leve, intermedia, severa, y asintomáticas. La acumulación de proteínas de cubiertas virales fue evaluada utilizando DAS-ELISA (sandwich con doble anticuerpo-ensayo inmunoabsorbente ligado a enzima), y los niveles de ARN viral se estudiaron por medio de transcripción reversa-reacción en cadena de la polimerasa (RT-PCR) semicuantitativa y RT-PCR cuantitativa. No hubo diferencia en la acumulación de proteínas virales para los dos virus asociados a esta enfermedad, al comparar plantas de lechugas con distintos niveles de severidad de síntomas. De forma similar, cuando se analizó la acumulación de ARNs virales en los diferentes niveles de la escala de severidad de síntomas utilizada, no hubo diferencias (P > 0,05) en la abundancia del ARN-3 del Virus Mirafiori de las venas grandes de la lechuga (MLBVV) o el ARN-2 del Virus asociado de las venas grandes de la lechuga (LBVaV) entre estos grupos. Esto sugiere que la severidad de los síntomas expresados en plantas afectadas por esta enfermedad no se relaciona necesariamente con una mayor acumulación de los virus asociados en el hospedero. Por lo tanto, lechugas que muestran síntomas suaves o moderados de venas grandes no necesariamente presentan menor acumulación de virus que plantas que muestran sintomatología más severa. Palabras clave: enfermedad de la vena ancha de la lechuga, expresión de síntomas, Ofiovirus, Varicosavirus, Lactuca sativa. INTRODUCTION Big-vein disease (BVD), one of the most important viral diseases in lettuce (Lactuca sativa L.), is present wherever this crop is cultivated. The economic importance of the disease is associated with losses caused by its typical symptomatology: chlorotic clearing around leaf veins followed by leaf distortion, maturity delay, reduction of head size, or absence of head formation (Walsh, 1994). For this reason, infected lettuce is often unmarketable, thus affecting yield and crop quality. BVD is widespread in lettuce crops in regions of the world with temperate or Mediterranean climates. Lettuce is the second most important vegetable grown in Chile and, according to the last Agricultural Census (INE, 2007), almost 7000 ha of this crop were harvested. A severe outbreak of BVD occurred in open-field and hydroponic-grown lettuce plants in Chile’s central zone during the 2003 winter season (Rosales et al., 2004). The disease has since then spread to all lettuce-growing regions of the country and is considered as the most important viral disease affecting this crop (Rosales et al., 2006). BVD is associated with two viral agents, Mirafiori lettuce big-vein virus (MLBVV; genus Ophiovirus) and Lettuce big-vein associated virus (LBVaV; genus Varicosavirus). Initially, it was postulated that this disease was caused by LBVaV, formerly known as Lettuce big-vein virus (LBVV) (Kuwata et al., 1983). Later, it was proposed that MLBVV, not LBVaV, is the causal agent of BVD since evidence showed that plants infected with LBVaV did not develop symptoms in the absence of MLBVV, whereas plants infected with MLBVV developed big-vein symptoms regardless of the presence or absence of LBVV (Roggero et al., 2000; Lot et al., 2002; Roggero et al., 2003). On the other hand, lettuce plants with big-vein symptoms that are serologically negative for MLBVV, but positive for LBVaV, were described in Italy (Roggero et al., 2003). Both MLBVV and LBVaV have almost always been detected in field-grown lettuce plants with BVD symptoms (Roggero et al., 2003; Navarro et al., 2004; 2005; Hayes et al., 2006) suggesting that LBVV may also play a role in symptom expression or disease development. In a more recent study, Sasaya et al. (2008) developed methods to isolate MLBVV and LBVaV from doubly infected lettuce plants, and separately and consecutively transfer viruses to lettuce seedlings using fungal isolate zoospores. These separation and inoculation methods allowed this group to demonstrate that only MLBVV-infected lettuce plants developed big-vein symptoms, while those infected only with LBVaV did not develop any, thus indicating MLBVV as the causal agent of BVD. Transmission is what these two viral agents have in common since both MLBVV and LBVaV are naturally acquired by the Olpidium brassicae chytrid fungus and are then vectored from root to root to other plants (Roggero et al., 2003). BVD is difficult to control because these viruses can survive from one crop to another in the fungus resting sporesand these structures can persist in the field for decades even in the absence of a host plant (Campbell, 1996). Moreover, the absence of safe and effective chemicals to control the fungal vector along with the lack of economically viable cultural control make it extremely difficult to avoid economic losses provoked by this viral disease. Considering this background information, effective long-term control of BVD is best accomplished through genetic resistance. However no L. sativa genotype has been identified as immune to big-vein (Bos and Huijberts, 1990; Ryder and Robinson, 1995) and only partial resistance is available in cultivated lettuce, characteristic expressed as the delayed appearance of symptoms combined with a reduced number of symptomatic plants at market maturity (Ryder and Robinson, 1995; Hayes and Ryder, 2007). Traditional breeding has developed cultivars with some levels of resistance such as Pacific and Pavane (Ryder and Robinson, 1991; Compositdb, 2004), but resistance levels of cultivars released to date are not as high (Kawazu et al., 2010). In this scenario, the use of resistant transgenic plants is an attractive alternative to control BVD. Kawazu et al. (2009) recently developed a transgenic lettuce line resistant to MLBVV by introducing inverted repeats of the viral coat protein gene (Kawazu et al., 2009). Further characterization of this line showed transgenic lettuce showed high resistance levels to BVD and this characteristic was stably inherited by subsequent generations (Kawazu et al., 2010). It has also been shown that symptomatic and asymptomatic plants from both resistant and susceptible lettuce cultivars can accumulate MLBVV and LBVaV (Hayes et al., 2006), and that some accessions of the wild relative Lactuca virosa L., an important source of resistance to BVD, may also have symptomless infections of MLBVV (Hayes et al., 2008). The mechanism responsible for the resistant or susceptible phenotype to BVD is unknown. Likewise, it is not clear what can drive symptom expression in different lettuce, wild and cultivated. To gain a better understanding of the role of virus accumulation in symptom severity in lettuce infected with big-vein, we studied viral antigen and viral RNA accumulation along with symptom severity induced in different systemically infected lettuce cultivars. MATERIALS AND METHODS Plant material Two sets of plants were used in our study. First, leaves from 14 field-grown lettuce cultivars maintained in a variety field plot located in the northern zone of Santiago, Chile were collected to assay virus levels by Double Antibody Sandwich (DAS) ELISA and semi-quantitative reverse transcription-polymerase chain reaction (sqRT-PCR). Fully expanded leaves from seven iceberg lettuce cultivars (Canery Row, Yardena, Desert Storm MI, Winterhaven, Vanguard 75 MT, Sea Green, and 0524), and seven romaine lettuce cultivars (Blonde Maraichere, Conconina, Ballon, LAC002, LAC003, LAC008, and PI278063) were sampled 4 mo after transplanting seedlings to the field during the 2008 winter season. These plant materials were naturally infected by BVD since the plot had a long history of BVD outbreaks. Lettuce leaves were brought to the lab, classified as mild, moderate, or severe for big-vein symptoms by the disease severity scale previously described by Hayes et al. (2006): Mild = narrow vein clearing and/or symptoms occurring on less than 50% of leaf area, moderate = both wide and narrow vein clearing, symptoms occurring on more than 51% of leaf area, and severe = wide vein clearing occurring on 100% of the leaf (Figure 1). RNA was extracted from this material as described below. Another set of plants was infected under greenhouse conditions for the quantitative analysis of virus RNA levels. Twenty-five plants of each genotype of the iceberg cvs. Sharpshooter and Winterhaven, romaine accessions from Instituto de Investigaciones Agropecuarias (INIA) germplasm bank LAC004 and LAC009, and the BVD tolerant cv. Pavane (Ryder and Robinson, 1995; Hayes et al., 2006) were germinated in a potting mix soil:perlite in a ratio of 2:1 and grown for 4 wk. Plants were then transferred to a 1 L pot containing the same potting mix. Plants were grown with a day/night temperature of 20/18 °C under natural greenhouse daylight conditions and the percentage of symptomatic plants was recorded weekly for 75 d post-transplant (DPT). Tissue was sampled from three plants showing mild symptoms and three plants showing severe symptoms. RNA was extracted from this material as described below. Serological detection Serological assays were carried out by DAS-ELISA with polyclonal antibodies. The commercial reagent source was Bioreba AG (Reinach, Switzerland) for MLBVV detection and Prime Diagnostics (Plant Research International B.V. Prime Diagnostics, Wageningen, The Netherlands) for LBVaV detection. Protocol followed the standard procedures recommended by the manufacturer in each case. Coating antibodies and conjugates were diluted 1/1000 for both reagent sets. A composite sample of about 0.3 g from at least three expanded leaves of each lettuce plant was used in the ELISA test and each sample was analyzed in duplicated wells on the plate. Absorbance readings were performed at 405 nm every 30 min for 90 min with the Bio-Kinetics Reader, EL 312e (BioTek Instruments, Winooski, Vermont, USA). The cut-off or threshold value, which discriminates positive samples, was calculated following the formula recommended by ELISA Data Analysis (Bioreba, 2009), which discriminates positive results from background readings. The formula, cut-off = (mean + 3 SD) × 1.1 considers the arithmetic mean of negative and blank control readings plus three standard deviations of these values multiplied by 1.1. Total RNA extractions Total RNA was isolated from approximately 100 mg of plant tissue by carrier adsorption-based isolation technology provided by the Invisorb Spin Plant RNA kit (Invitek, Berlin, Germany), and the manufacturer’s protocol was followed. First, the sample was thoroughly ground under liquid nitrogen to a fine powder with a mortar and pestle. Then, the powder was resuspended in the cell lysis solution provided by the kit and, after removing genomic DNA by prefiltration, total sample RNA was bound to a silica-based spin filter by pouring the lysate mixed with ethanol. After washing the filter, elution of total RNA was performed with specific reagents contained in the kit. The purity and concentration of the nucleic acid was determined by spectrophotometric measurement of optical density (OD) at 260 and 280 nm. Semi-quantitative RT-PCR (sqRT-PCR) This analysis was performed with target genomic sequences of LBVaV and MLBVV located on viral RNA 2 and 3, respectively. In both cases, a region of the coat protein (CP) gene was selected for amplification. The reference housekeeping gene was the mitochondrial NADH dehydrogenase subunit 4 (Nad4) from lettuce. First, cDNA was synthesized from 2 mg of total RNA in a reaction (20 mL) containing 1 mL 5X First-Strand Buffer [250 mM Tris-HCl (pH 8.3), 375 mM KCl, 15 mM MgCl2], 1 µM random primer, 0.5 mM of each dNTP, 0.01 M DTT, 10 U of RNaseOUT, Recombinant Ribonuclease Inhibitor (40 units μL-1) (Invitrogen, Carlsbad, California, USA), and 50 U of M-MLV Reverse Transcriptase (200 U μL-1) (Invitrogen). The reverse transcription step was performed following incubation for 10 min at 25 ºC, 1 h at 37 ºC, and 10 min at 75 ºC. Then, for RT-PCR, 2 μL of the 20 μL RT reaction was used as template. The reaction was performed in a final volume of 25 μL. The mixture contained 2.5 μL of 10X PCR buffer (500 mM KCl, 100 mM Tris-HCl (pH 9.0 at 25 °C) and 1.0% Triton X-100®), 1.6 mM MgCl2, 0.4 mM each of dNTP (dATP, dGTP, dCTP, and dTTP), 0.1 μM of each primer (Table 1), 2.5 U of Taq DNA polymerase (Promega Corporation, Madison, Wisconsin, USA). Thermocycling conditions were initial denaturation for 5 min at 94 ºC and the following cycle was then repeated 36 times for MLBVV and 27 times for LBVaV and Nad4 gene: 30 s at 94 ºC, 30 s at 60 ºC, and 30 s at 72 ºC. In all cases, a final extension of 2 min at 72 ºC was included. Products from sqRT-PCR were separated and visualized by electrophoresis in agarose gels stained with ethidium bromide. Table 1. Primer sequences and reaction conditions used for semi-quantitative reverse transcription-polymerase chain reaction (RT-PCR) and real time PCR.
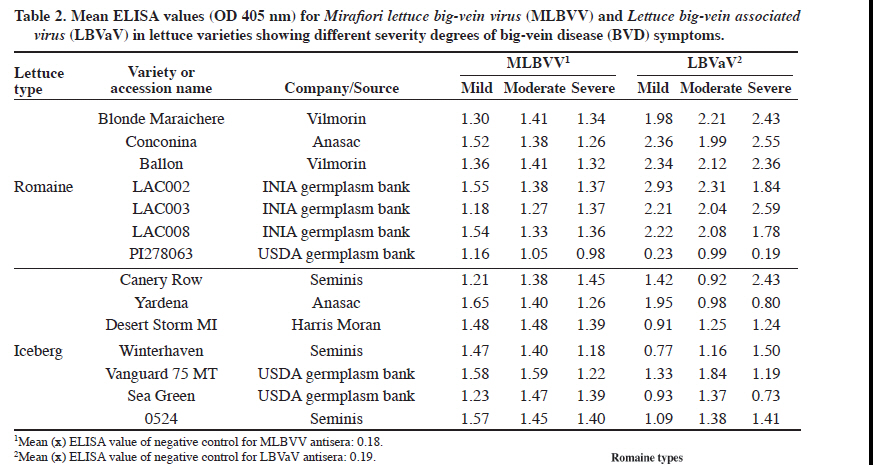
MLBVV: Mirafiori lettuce big-vein virus; LBVaV: Lettuce big-vein associated virus. Acquisition of gel images and semi-quantitative analysis Images of RT-PCR ethidium bromide-stained agarose gels were taken with the UltraCam 8gD digital imaging system (UltraLum, Claremont, California, USA) and band quantification was performed with SigmaGel gel analysis software (Jandel Scientific, San Rafael, California, USA) to determine pixel intensity. Data was then normalized with regard to Nad4 pixel intensity and expressed as mean ± standard error of mean (SEM). Semi-quantitative measurements of viral RNAs were expressed as arbitrary units (AU). Statistical significance was verified by two-way ANOVA. A value of P < 0.05 was considered statistically significant. Data analysis was with STATA 10.1 (StataCorp 2007. Stata Statistical Software: Release 10. College Station, TX: StataCorp LP). Absolute quantification by real time PCR Three experimental and three biological replicates made up each analyzed sample. RNA (1 μg) was transcribed into cDNA by random hexameres (IDT, Integrated DNA Technologies, Coralville, Iowa, USA) and M-MLV reverse transcriptase (200 U μL-1) (Invitrogen) under the same conditions described for reverse transcription of sqRT-PCR, but it was followed by RNase H treatment (New England Biolabs, Ipswich, Massachusetts, USA). Total cDNA was quantified at 260 nm with a Picodrop spectrophotometer (Picodrop Limited, Walden, UK). Quantitative PCR (qPCR) was performed by the Step One Real Time PCR System (Applied Biosystem, Carlsbad, California, USA) in a final 20 mL volume. Reaction components were 10 μL of Fast SYBR® Green PCR Master mix (Applied Biosystem), 300 ng of each primer (Table 1), and either 125 ng of cDNA or 5 μL of plasmid dilution standard. Cycling parameters were as follows: (i) initial incubation at 95 °C for 10 min and (ii) 40 cycles of 3 s at 95 °C followed by 30 s at 60 °C. Dissociation curves were then generated to evaluate the formation of non-specific products. Cycle threshold (Ct) values were automatically determined with Step One version 2.0 software (Applied Biosystems, Foster City, California, USA). To quantify virus in lettuce samples, two plasmids (3015 bp backbone) containing a fragment of 562 (nucleotides 66-627 in GenBank accession AF525935 for MLBVV) or 943 bp (nucleotides 121-1063 in GenBank accession AY366414 for LBVaV) of coat protein gene of each virus generated the standard curve. Concentration of purified plasmid DNA was determined spectrophotometrically (Picodrop Limited, Walden, UK), and plasmid copy number was then determined by calculating the molecular weight of each cloned plasmid according to recommendations given by Applied Biosystems (2003). Ten-fold dilution series of plasmid DNA generated the standard curve. This curve ranged from 5 × 107 to 5 × 102 copies for plasmid containing the MLBVV insert, and 5 × 107 to 5 x 103 copies for plasmid containing the LBVaV coat protein insert. These dilutions were used in qPCR reactions. Analysis of copy number, linear regression, and melting curve analysis were performed with the Step One version 2.0 software. Finally, qPCR data were analyzed for normality by the Shapiro-Wilk test, and statistical significance was verified by the non-parametric Kruskal-Wallis test. In the case where statistical significance was found, post-hoc multiple comparison tests were performed between groups. RESULTS AND DISCUSSION Serological detection of MLBVV and LBVaV in lettuce leaves and symptom recognition A previous study about LBVaV and MLBVV distribution in the plant revealed that the highest concentration occurred in roots and old lettuce leaves, and when hearted (L1) to fully expanded (L7) leaves were compared, higher titers of both viruses were consistently found in the latter (Navarro et al., 2004). Therefore, fully expanded leaves allocated in the L6 layer were selected to perform all tests described in the present study. A total of 42 lettuce leaf samples were analyzed by ELISA using specific antisera for the presence of the virus associated with BVD. All samples except romaine cv. PI278063, were positive to the presence of the two viruses associated with BVD (Table 2). Leaves from this cultivar classified with mild and severe symptoms were positive only to the presence of MLBVV, and absorbance values obtained in these samples for LBVaV did not reach the cut-off or threshold value required to be considered positive. In several other studies, both MLBVV and LBVaV have almost always been detected in field-grown lettuce plants with big-vein symptoms (Lot et al., 2002; Roggero et al., 2003; Navarro et al., 2004; Colariccio et al., 2005; Hayes et al., 2006; Sasaya et al., 2008). In our study, ELISA results revealed that in analyzed naturally infected lettuce leaves, 100% of plants with mild, moderate, and severe symptoms were infected by MLBVV, as well as 96% of the samples were co-infected by LBVaV and MLBVV. Hayes et al. (2006) found similar results for symptomatic plants, although their results also showed that, in the case of non-symptomatic plants, 24 and 69% were positive for MLBVV and LBVaV, respectively. Our results do not include data for non-symptomatic plants because the field selected for this study had a long history of BVD outbreaks and presented 100% symptomatic plants at the end of the crop season. It is now widely accepted that MLBVV is the causal agent of BVD, and LBVaV does not appears to play any etiological role in symptom expression or influence the outcome of MLBVV infection (Lot et al., 2002; Sasaya et al., 2008). However, co-infection with both MLBV and LBVaV is common and most lettuce plants become infected with both viruses after 100 DPT, thus indicating the high transmission efficiency of both viruses by the fungal vector (Navarro et al., 2004). Field samples analyzed in this study were collected 4 mo post-transplant, which explains the high rate of co-infection found in these samples. Another interesting aspect is the high variation for symptom expression found for each cultivar used in this study. The symptom scale allowed classifying them as mild, moderate, and severe. All cultivar symptoms were chlorosis or white vein-banding with leaf crinkling. These symptoms were more obvious in iceberg than in romaine types (Figure 1). However, the OD value obtained by the DAS-ELISA test was not necessarily related to symptom severity observed in lettuce samples, hence showing that disease symptoms for BVD do not necessarily reflect a higher accumulation of these viruses in plant host tissue (Table 2). Semi-quantitative measurement of RT-PCR products and BVD symptom severity Relative abundance of viral RNAs in infected plants was measured by sqRT-PCR. This parameter was compared among plants classified in each symptom intensity group and among lettuce types used in this study. Analyzed plant materials correspond to the same cultivars listed in Table 2. Figure 2 shows agarose gel visualization of sqRT-PCR fragments obtained after viral RNA amplification. In order to have semi-quantitative image analysis, band density was analyzed with an image program and normalized with regard to Nad4 pixel intensity. Gel visualization and semi-quantitative image analysis showed higher MLBVV-RNA3 accumulation in iceberg than in romaine lettuce types. Normalized data showed that iceberg accumulated 2.2 times more viral RNA than romaine types (P < 0.05) (Table 3). On the other hand, romaine types showed increased LBVaV-RNA2 accumulation (P < 0.05). Table 3. Relative quantification of (Mirafiori lettuce big-vein virus, MLBVV, and Lettuce big-vein associated virus, LBVaV) viral RNAs among romaine and iceberg varieties used in sqRT-PCR analysis.
*Indicates statistical significance according to two-way ANOVA (P < 0.05). Data is expressed as mean ± standard error of the mean (SEM). AU: arbitrary units. However, when we compared virus accumulation for different BVD symptom severity levels in the disease scale, statistical analysis showed that the mean relative abundance of MLBVV-RNA3 was not significantly different among analyzed disease severity groups (mild, moderate, and severe). Similar results are shown for LBVaV-RNA2 accumulation (Table 4). Table 4. Relative quantification of (Mirafiori lettuce big-vein virus, MLBVV, and Lettuce big-vein associated virus, LBVaV) viral RNAs among different severity groups for romaine and iceberg cultivars used in sqRT-PCR analysis.
ns: non significant according to two-way ANOVA (P > 0.05). Data is expressed as mean ± standard error of the mean (SEM). AU: arbitrary units. Absolute quantification of viral RNAs by real time PCR Considering that semi-quantitative measurements of viral RNAs did not show any relationship between disease symptom severity and viral RNA accumulation in infected lettuce plants, we decided to use a more powerful technique that allows the quantitative measurement of viral RNA accumulation. Tthree biological and three technical replicates of each plant disease severity group were used in the experiment. Quantification of MLBVV and LBVaV in plant samples was performed by comparing the cycle threshold (Ct) value of each sample to the Ct values of the standard regression lines generated with plasmid containing a fragment of the coat protein gene of each virus. Thus, the amount of starting template in the PCR reaction, expressed as the copy number of target MLBVV-RNA3 or LBVaV-RNA2 equivalents per microgram (μg) of total RNAs in the original plant extract, could be accurately determined (Table 5). These values were used as estimates of viral accumulation in the different plant samples belonging to the different symptom severity groups used in this assay. Table 5. Absolute quantification of viral RNAs associated with big-vein disease by real time polymerase chain reaction (PCR).
1Quantification of Mirafiori lettuce big-vein virus (MLBVV)-RNA3 in plant samples was performed by comparing cycle threshold (Ct) value of each sample to Ct values of the standard regression lines (R2 = 0.999; Y = -3.285x + 45.135). 2Quantification of Lettuce big-vein associated virus (LBVaV)-RNA2 in plant samples was performed by comparing Ct value of each sample to Ct values of the standard regression lines (R2 = 0.0996; Y = -3.114x + 43.057). Data is expressed as means ± standard error of the mean (SEM). In all cases, tissue from fully expanded leaves was used for analysis. Statistical analysis showed that the quantitative data was not normally distributed so statistical significance was verified by the non-parametric Kruskal-Wallis test. The analysis revealed there were no significant differences (P > 0.05) in the copy number or viral RNA equivalents among lettuce samples assigned to each symptom severity group (Table 6). This was valid for the quantification of MLBVV-RNA3 and LBVaV-RNA2. However, this test found significant differences (P < 0.05) for viral equivalents of MLBVV-RNA3 among lettuce cultivars used in the experiment. Post-hoc comparison tests between lettuce cultivars resulted in differences (P < 0.005) in the following pairs: ‘LAC004’ and ‘Winterhaven’, ‘LAC009’ and ‘Sharpshooter’, as well as ‘Sharshooter’ and ‘Winterhaven’ (data not shown). However, this result probably does not have any biological meaning because we used the viral RNA copy number for the mild, moderate, and severe lettuce samples all together. Table 6. Absolute quantification of (Mirafiori lettuce big-vein virus, MLBVV, and Lettuce big-vein associated virus, LBVaV) viral RNAs among different disease severity groups used in sqRT-PCR analysis.
ns: non significant according to Kruskal-Wallis test (P > 0.05). Data is expressed as median ± interquartile range. Several analyses of plant-virus interaction have failed to detect a relationship between symptom severity and virus accumulation (Handford and Carr, 2007; Pagán et al., 2007) although there are some exceptions (Bazzini et al., 2007). Recent studies indicate that symptoms are derived from specific interactions between virus and host components having the potential to disrupt host physiology. In this scenario, variations in symptom severity do not reflect the level of virus accumulation, suggesting that competition for host resources is not the major contributor to this disruption (Culver and Padmanabhan, 2007). CONCLUSIONS Most models assume that virulence is an unavoidable consequence of within-host multiplication of parasites. On the other hand, interaction of a plant with a compatible viral pathogen generally leads to a systemic infection in which the host manifests disease symptoms. In the case of big-vein disease (BVD) in lettuce, it is common to find a variation in vein-banding symptoms among different lettuce cultivars, and sometimes within plants from the same cultivar. In accordance with the proposed objective, we have addressed the role of virus accumulation in symptom severity in BVD-infected lettuce. First, the serological test showed that there was no difference in virus coat protein accumulation (for Mirafiori lettuce big-vein virus or Lettuce big-vein associated virus) among lettuce plants showing different disease symptom severity, allowing us to classify them as mild, moderate, and severe. Then, we explored the viral RNA accumulation and symptom severity induced in different systemically infected mature lettuce leaves. Two different techniques (semi-quantitative RT-PCR and real time PCR) indicated that the abundance of MLBVV-RNA3 or LBVaV-RNA2 did not have an impact on symptom severity shown by the host plant. Symptom severity expressed by bigvein diseased lettuce plants did not necessarily reflect virus multiplication of either virus associated with this disease, as defined by viral protein or viral RNA accumulation. ACKNOWLEDGEMENTS This research study was supported by FONDECYT Proyecto Regular de Iniciación en Investigación Nº11060173 and project FIA-PI-C-2005-1-A-051. LITERATURE CITED
Copyright 2011 - Chilean Journal of Agricultural Research The following images related to this document are available:Photo images[cj11008f1.jpg] [cj11008t2.jpg] [cj11008t1.jpg] [cj11008t6.jpg] [cj11008t5.jpg] [cj11008t3.jpg] [cj11008f2.jpg] [cj11008t4.jpg] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}