 |
| About Bioline | All Journals | Testimonials | Membership | News |
|
||||||
|
||||||
Chilean Journal of Agricultural Research, Vol. 71, No. 1, 2011, pp. 73-82 RESEARCH Chemical Composition, Insecticidal, and Antifungal Activities of Fruit Essential Oils of Three Colombian Zanthoxylum Species Composición Química, Actividades Insecticida y Antifúngica de Aceites Esenciales de Frutos de Tres Especies Zanthoxylum de Colombia. Juliet A. Prieto1*, Oscar J. Patiño1, Wilman A. Delgado1, Jenny P. Moreno1, and Luis E. Cuca1
[1]
Universidad
Nacional de Colombia, Facultad de Ciencias, Bogotá, KR 30 45 03, Colombia. AA
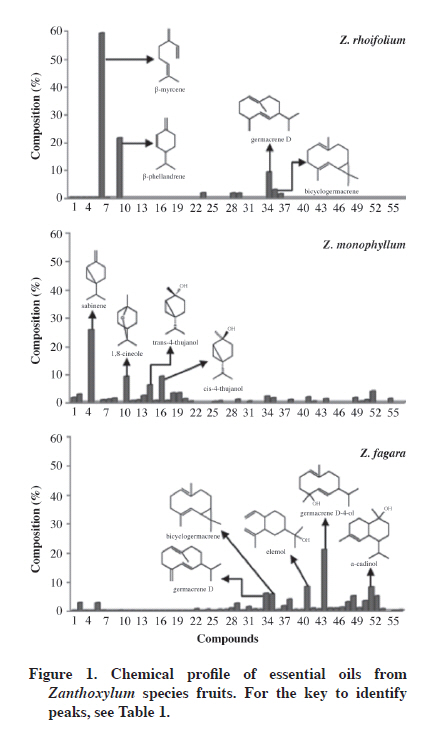
14490. Received: 7 June 2010 Code Number: cj11009 ABSTRACT This study determined the chemical composition of essential oils isolated from Zanthoxylum monophyllum (Lam.) P. Wilson, Z. rhoifolium Lam., and Z. fagara (L.) Sarg. fruits by steam distillation, as well as testing antifungal and insecticidal activities of essential oils as potential pesticides. Gas chromatography-mass spectrometry (GC/MS) analysis identified 57 compounds. The main constituents in Z. rhoifolium oil were β-Myrcene (59.03%), β-phellandrene (21.47%), and germacrene D (9.28%) , the major constituents of Z. monophyllum oil were sabinene (25.71%), 1,8-cineole (9.19%), and cis-4-thujanol (9.19%), whereas fruit oil of Z. fagara mainly contained germacrene D-4-ol (21.1%), elemol (8.35%), and α-cadinol (8.22%). Zanthoxylum fagara showed the highest activity on Colletotrichum acutatum Simmonds (EC50 153.9 μL L-1 air), and Z. monophyllum was the most active against Fusarium oxysporum Schlechtend: Fr. f. sp. lycopersici (EC50 140.1 μL L-1 air). Zanthoxylum monophyllum essential oil showed significant fumigant activity against Sitophilus oryzae (L.). This study demonstrated that Zanthoxylum essential oils exhibit important fungicidal activity on F. oxysporum and C. acutatum, which could become an alternative to synthetic fungicides to control plant fungal diseases, and Z. monophyllum oil is a potential fumigant against S. oryzae. Key words: Zanthoxylum monophyllum; Z. rhoifolium; Z. fagara; Sitophilus oryzae; Fusarium oxysporum f. sp. lycopersici; Colletotrichum acutatum; fumigant, antifungal, insecticidal. RESUMEN En este estudio se determinó la composición química de los aceites esenciales de frutos de Zanthoxylum monophyllum (Lam.) P. Wilson, Z. rhoifolium Lam. y Z. fagara (L.) Sarg. obtenidos mediante destilación por arrastre con vapor y se evaluó la actividad antifúngica e insecticida de los aceites esenciales para estimar su uso como posibles plaguicidas. El análisis por cromatografía de gases-espectrometría de masas (CG/EM) permitió la identificación de 57 compuestos. β-Mirceno (59,03%), β-felandreno (21,47%) y germacreno D (9,28%) fueron los componentes principales del aceite de Z. rhoifolium; los principales componentes del aceite de Z. monophyllum fueron sabineno (25,71%), 1,8 -cineol (9,19%) y cis-4-thujanol (9,19%), mientras que el aceite de frutos de Z. fagara está compuesto principalmente por germacreno D-4-ol (21,1%), elemol (8,35%) y α-cadinol (8,22% ). Zanthoxylum fagara presentó la mayor actividad sobre Colletotrichum acutatum Simmonds (EC50 153,9 μL L-1 de aire) y Z. monophyllum fue el más activo contra Fusarium oxysporum Schlechtend: Fr. f. sp. lycopersici (EC50 140,1 μL L-1 de aire). El aceite esencial de Z. monophyllum mostró actividad insecticida significativa contra Sitophilus oryzae (L.) Este estudio demuestra que los aceites esenciales de Zanthoxylum poseen importante actividad antifúngica sobre F. oxysporum y C. acutatum y podría convertirse en una alternativa frente a los fungicidas sintéticos empleados comúnmente para el control de enfermedades de las plantas, así como el aceite de frutos de Z. monophyllum tiene un potencial para ser utilizado como fumigante contra S. oryzae. Palabras clave: Z. monophyllum; Z. rhoifolium; Z. fagara; Sitophilus oryzae; Fusarium oxysporum f. sp. lycopersici; Colletotrichum acutatum; fumigante, antifúngicos, insecticida. INTRODUCTION Crop losses due to insects and plant diseases caused by fungi, bacteria, and viruses cause significant economic losses (Kotan et al., 2008; Kordali et al., 2008). Insect pests often cause extensive loss of products stored in tropical and semitropical environments (Isman, 2000). For example, Sitophilus species are serious cosmopolitan pests of stored grain (Liu and Ho, 1999). Sitophilus zeamais Motschulsky (maize weevil), S. oryzae L. (rice weevil), and S. granaries L. are the main representatives of this genus, which principally attack rice (Oryza sativa L.), maize (Zea mays L.), wheat (Triticum sativum Lam.), and sorghum (Sorghum bicolor (L.) Moench) among others, through direct feeding on grain kernels causing unfavorable effects on food quality, safety, and preservation (Huang et al., 1997; Tapondjou et al., 2002; Kim et al., 2003; Park et al., 2003; Arannilewa, 2007). Harvest losses due to fungal disease in world crop production may amount to 12% or more in developing countries (Horbach et al., 2010). Many pathogens including Fusarium oxysporum (vascular wilt), F. solani (fruit rot) and Colletotrichum gloeosporoides (fruit rot) cause severe pre- and post-harvest damage to agriculture (Bajpai et al., 2008). Fusarium is a plant pathogen that causes wilt diseases of several economically important plants and is also known to produce toxins thought to contribute to wilting by affecting cell membrane permeability and disrupting cell metabolism (Garcés de Granada et al., 2001; Pawar and Thaker, 2007). Colletotrichum are pathogens that cause anthracnose in a wide range of woody and herbaceous crops. Symptoms are broad-ranging and include stem rot, dieback, and seedling blight. Fruits are affected during the pre- and post-harvest period (Roca et al., 2003; Muñoz et al., 2009). Synthetic insecticides and fumigants are widely used to control grain pests and plant diseases. However, the indiscriminate application of synthetic products has led to various problems including toxic residues in treated products, environmental pollution, and resistance against pesticides by microorganisms and grain insect pests (Huang et al., 1997; Isman 2006; Bakouri et al., 2008; Kotan et al., 2008; Ye et al., 2010). Therefore, because of increasing drawbacks of the continued use of conventional fumigants, an effort is needed to develop new alternative pesticides to replace those being currently used. Essential oils are potential botanical sources of alternative compounds to fumigants being currently used because of their low toxicity for warm-blooded animals, high volatility and toxicity for stored grain pests and plant microorganisms (Lee et al., 2001; Abad et al., 2007). Zanthoxylum genus (Rutaceae) is made up of about 250 species of trees and shrubs in the world’s tropical and temperate regions (Pirani, 1993). It is economically very important as a source of edible fruits, raw material for the cosmetics and perfume industries, as well as culinary applications. In Asia, Z. bungeanum Maxim. fruits are the most popular huajiao commercial product called “da hong pao’’ (big red robe). “Green huajiao”, fruit of Z. schinifolium Siebold & Zucc. (Yang, 2008), is the other widely used spice in Sichuan. Zanthoxylum species have shown significant insecticidal and antifungal activity. The bark methanol extract of Z. xanthoxyloides caused significant mortality rates in S. oryzae and Callosobruchus maculatus, two stored-product insect pests (Owusu et al., 2007). Zanthoxylum monophyllum bark methanol extract showed significant activity against seven human pathogen fungi (Gómez et al., 2007). Ethanolic extracts of Z. americanum leaves, fruits, stem bark, and root demonstrated a broad spectrum of antifungal and antibacterial activity against Candida albicans, Aspergillus fumigatus, Cryptococcus neoformans, and Fusarium oxysporum (Bafi-Yeboa et al., 2005). In addition to culinary applications, many species of Zanthoxylum are used in traditional medicine especially in America, Africa, and Asia. Zanthoxylum rhoifolium Lam is popular in South America for inflammatory, microbial, cancerous, and malaria processes (Da Silva et al., 2007a; 2007b). Zanthoxylum fagara is used in Cuba for the treatment of diarrhea, chest diseases, intermittent fever, earaches, and tooth diseases (Dieguez-Hurtado et al., 2003). Zanthoxylum monophyllum is used as an analgesic to treat nasal inflammation, jaundice, and eye infections in Venezuela and as colorant (Cuca et al., 1998; Díaz and Ortega, 2006; Da Silva et al., 2007a). Zanthoxylum species accumulate volatile oils in leaves, fruits, and inflorescences (Adesina, 2005). There are numerous reports on the chemical composition and the various biological activities of Zanthoxylum species essential oils (Choochote et al., 2007; Boehme et al., 2008; Yang, 2008). The chemical composition of essential oils of Z. rhoifolium flowers, fruits, and leaves (Gonzaga et al., 2003; Moura et al., 2006; Da Silva et al., 2007a; 2007b; Boehme et al., 2008), and biological properties of fruit and leaf essential oils, such as antibacterial and cytotoxic resultshave been previously reported (Moura et al., 2006; Da Silva et al., 2007b; Boehme et al., 2008). The volatile chemical composition of Z. monophyllum and Z. fagara leaves from Costa Rica have also been reported (Setzer et al., 2005). However, there are no studies about their biological properties. The composition and biological properties of fruit essential oils of Z. monophyllum and Z. fagara have not yet been investigated. We report the chemical composition, insecticidal activity against S. oryzae, and antifungal activity against F. oxysporum f. sp. lycopersici and C. acutatum of fruit essential oils of three Colombian plant species: Z. monophyllum, Z. fagara, and Z. rhoifolium. MATERIALS AND METHODS Plant material The fruits of Z. monophyllum (4º11’24.3’’ N, 74º30’48.9’’ W) and Z. fagara (4º11’22.0’’ N, 74º30’58.4’’ W) were collected in January 2008 in the town of Icononzo, Tolima, Colombia, whereas Z. rhoifolium fruits (4º19’30.4’’ N, 74º26’17.1’’ W) were collected in February 2008 in the town of Fusagasugá, Cundinamarca, Colombia. Plant samples were identified by the Colombian National Herbarium of the Universidad Nacional de Colombia. Voucher specimens of Z. monophyllum (Lam.) P. Wilson (COL-517520), Z. rhoifolium Lam. (COL-522896), and Z. fagara (L.) Sarg. (COL-522891) were deposited in the Colombian National Herbarium of the Universidad Nacional de Colombia, Bogotá, Colombia. Isolation of essential oils Samples of fresh fruits (approximately 2 kg) of each Zanthoxylum species were submitted to steam distillation (ca. 4 h). Oils were dried over anhydrous sodium sulfate and stored at 0-5 ºC for further analysis. Gas chromatography-flame ionization detector analysis (GC/FID) Volatile compound analysis was performed with a gas chromatography system (Hewlett Packard 5890 GC) with a fused capillary column (50 m × 0.25 mm × 0.25 µm, HP-5MS, Crossbond 5% phenyl-95% dimethylpolysiloxane, Sigma-Aldrich, St. Louis, Missouri, USA) directly coupled to a flame ionization detector (FID). Injection conditions were the following: injector temperature at 250 ºC; oven temperature program at 45 ºC (2 min), 150 ºC (5 min) at a rate of 2 ºC min-1, then at 150 ºC (2 min), 280 ºC (5 min) at a rate of 8 ºC min-1; split 30:1 during 1.50 min, carrier gas He: 1 mL min-1, constant flow; sample volume 1 µL. Gas chromatography-mass spectrometry analysis (GC/MS) The GC/MS analyses were performed in electronic impact (EI) mode with a Hewlett Packard-5890 GC system with a fused capillary column (50 m × 0.25 mm × 0.25 µm, HP-5MS, Crossbond 5% phenyl-95% dimethylpolysiloxane) directly coupled to a Hewlett Packard 5973 selective mass detector. Injection conditions were the same as described above. The mass spectrometer was operated at 70 eV. Oil component identification Oils were analyzed by GC-MS and GC with capillary columns (HP-5MS). Chemical constituents were identified based on the comparison of their mass spectral pattern and retention indices with those obtained from the Wiley 138.L, NBS 75K.L, and SDBS libraries, as well as those published by Adams (1995). Retention indices (RI) were calculated according to the literature (Van Den Dool and Kratz, 1963). Insect rearing Rice weevils were obtained from a colony maintained in the Laboratory of Entomology, Universidad Nacional de Colombia, Bogotá. Weevils were reared on maize grains. Cultures were maintained in the dark at 25 ± 1 °C and 70 ± 5% relative humidity. Fungal cultures Fusarium oxysporum f. sp. lycopersici was obtained from the culture collection of the Universidad de Cundinamarca (Laboratory of Phytopathology). Colletotrichum acutatum was obtained from the culture collection of the Universidad Nacional de Colombia, Bogotá (Laboratory of Vegetal Natural Products, Faculty of Science). Cultures were maintained and grown on potato dextrose agar (PDA) medium and incubated at 28 ± 1 °C. Insecticidal activity assay To determine the fumigant toxicity of Zanthoxylum oils, paper filter disks (Whatman Nº 1, 2-cm diameter pieces) were adhered to the inside of the Petri dish covers (90 x 15 mm) and then impregnated with oil at doses calculated to give equivalent fumigant concentrations of 242-967 μL L-1 air (20, 40, 60, and 80 μL oil). Twenty adult insects (1 to 10-d-old) were placed on each Petri dish. Phostoxin®-Fugran (phosphine - 300 μg L-1 air) and Nuvan® 50 E.C. (clorvox - 100 μL L-1 air) were used as positive controls. Petri dishes were sealed with Parafilm and incubated at 25 ± 1 °C and 70 ± 5% RH. Each concentration and control was replicated three times. Mortality was determined after 12, 24, and 48 h from the start of exposure (Negahban et al., 2007; Kotan et al., 2008). Insect mortality percentage (%M) was calculated by Abbott’s correction formula (Pitasawat et al., 2007). LC50 and LC95 values, as well as the corresponding 95% confidence intervals, were estimated by probit analysis (Finney, 1971). In vitro antifungal activity PDA plates were prepared with glass Petri dishes (90 x 15 mm) for the in vitro antifungal activity test. Agar plugs of actively growing cultures in PDA were placed on one half of the Petri dish (covered with PDA) and a sterilized paper disk was placed 2 cm from them. A 10 μL aliquot of each essential oil was added to the paper disks in each of the PDA plates (maximum 5 μL per disk). Plates were immediately sealed with Parafilm after adding each essential oil and incubated for 3 d at 28 °C. The diameter of concentric fungal mycelia was measured and compared with the untreated control. Medium effective concentration (EC50) values were determined for essential oils that caused fungal growth inhibition. Aliquots of 2, 5, 7, 10, and 15 μL (23.5 - 176.5 μL L-1 air) of each essential oil were added to paper disks in each of the PDA plates (maximum 5 μL per disk). Plates were replicated three times in each treatment. (Lee et al., 2007; Kotan et al., 2008). In addition, Benlate 50WP (methyl-[1-[(butylamino) carbonyl]-1H-benzimidazol-2-yl]carbamate - 2 mg mL-1) and Derosal®-Bayer (methyl-1H-benzimidazol-2-ylcarbamate - 1 mg mL-1) were employed as chemical controls in the F. oxysporum f. sp. Lycopersici and C. acutatum assays, respectively. Statistical analysis Data are presented as mean ± standard error. Statistical significance was determined by the Duncan and Tukey tests; ANOVA determined whether results obtained for antifungal and insecticidal activity assays were statistically different. Statistical significance was set at P < 0.05. RESULTS AND DISCUSSION Oil chemical composition Fifty-seven compounds were identified by gas chromatography and mass spectrometry data in the fruit essential oils of three Zanthoxylum species (Table 1). Oils mainly contain monoterpenes and sesquiterpenes. Identified volatile components accounted for 89 to 99% of oil composition. Monoterpenes represent more than 70% of Z. rhoifolium oil composition (80.5%), Z. monophyllum (71.6%), and only 6.16 % of Z. fagara oil composition. Sesquiterpenes represent 88.8% of Z. fagara oil composition. Chemical profiles obtained for these oils showed differences in composition among the studied species (Figure 1) although volatile constituents have been previously reported in other species of the Zanthoxylum genus (Moura et al., 2006). Table 1. Chemical composition of Zanthoxylum species oils.
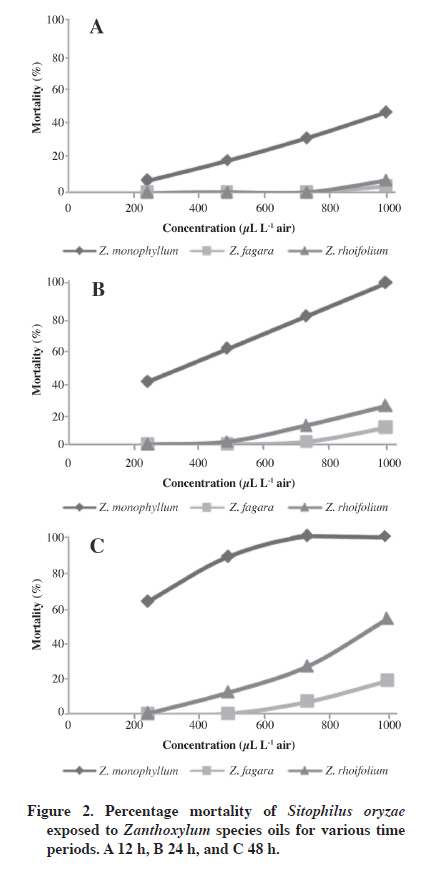
1Calculated retention index. The main constituents found in Z. rhoifolium fruit oil were β-myrcene (59.03%), β-phellandrene (21.47%), germacrene D (9.28%), and bicyclogermacrene (3.13%). Approximately 99.2% of Z. rhoifolium oil composition was characterized. The remaining unidentified components were mainly sesquiterpenes. The abundance of monoterpenoid and sesquiterpenoid compounds in fruit essential oil is in accordance with one previous report (Gonzaga et al., 2003); however, oil composition described in this study was qualitatively and quantitatively different, particularly regarding the major constituents. This suggests a considerable variability in the studied oil samples because of the influence of the ecological and chemical environment of each species, which affects the presence and abundance of secondary metabolites (Spitaler et al., 2006). The major constituents of oil derived from fruits collected in Brazil were reported as menth-2-en-1-ol (46.2%), β-myrcene (30.2%), (-)-linalool (15%), and terpineol (8.45%). The major constituents identified in Z. monophyllum fruit oil were sabinene (25.71%), 1,8-cineole (9.19%), trans-sabinene hydrate (9.19%), and cis-sabinene hydrate (6.25%). Zanthoxylum fagara fruit oil mainly contained germacrene D-4-ol (21.1%), elemol (8.35%), α-cadinol (8.22%), germacrene D (5.96%), bicyclogermacrene (5.75%), epi-α-muurolol (5.15%), and 5-neo-cedranol (5.12%). Approximately 89.94% of Z. monophyllum and 94.2% of Z. fagara oil compositions were characterized. The remaining unidentified components were monoterpenes and sesquiterpenes. The detailed composition of fruit essential oils of Z. fagara and Z. monophyllum are reported for the first time in this study. Insecticidal activity of the oils Results show that the essential oils of Z. fagara, Z. rhoifolium, and Z. monophyllum have different insecticidal activity against S. oryzae adults. Insecticidal activity rises by increasing the dose and exposure times (Figure 2). Zanthoxylum monophyllum essential oil showed the best fumigant activity against rice weevil, Z. rhoifolium oil had weak fumigant toxicity while Z. fagara essential oil was inactive (Figure 2). Zanthoxylum monophyllum essential oil caused significant mortality (about 90 to 99%) at 976 μL L-1 air dose after 24 h exposure. After 48 h of treatment, a 484 μL L-1 air dose is enough to cause 90% insect mortality. Fumigant activity of Z. monophyllum can be attributed to 1,8-cineole, terpinen-4-ol, and α-terpinene present in the essential oil; these compounds have shown 100% mortality on insects of the Sitophilus genus after 12 h exposure (Lee et al., 2004; Kordali et al., 2006). The commercial fumigants Phosphamin (100 μg L-1 air) and Nuvan 50 (50 μL L-1 air) showed 100% mortality before 12 h exposure. Zanthoxylum monophyllum essential oil was less active than commercial products. Higher concentrations of essential oil or longer exposure times are necessary to obtain effects similar to those of commercial fumigants (Table 2). Table 2. Fumigant toxicity of Zanthoxylum monophyllum and Z. rhoifolium essential oils against S. oryzae.
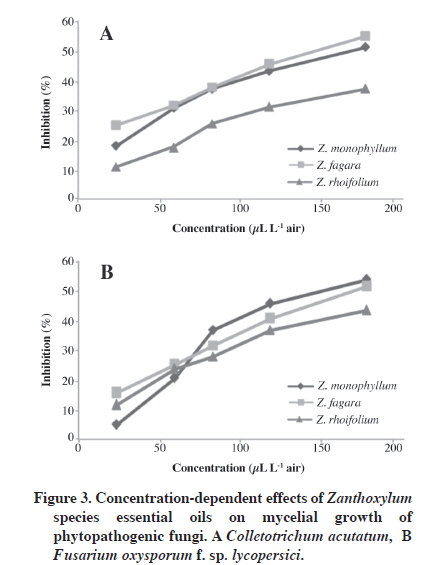
1Fiducial limits. Terpinen-4-ol, 1,8-cineole, and α-terpinene are absent in Z. rhoifolium and Z. fagara essential oils. Nevertheless, Z. rhoifolium shows insecticidal activity, which suggests that this activity can be due to the main oil components (β-myrcene and β-phellandrene) or to a possible synergistic effect of the compounds in the essential oil. Oil antifungal activity Essential oils were also evaluated as fumigants against two phytopathogenic fungi, F. oxysporum f. sp lycopersici and C. acutatum, which resulted in significant antifungal activity against both. For doses greater than 7 µL per Petri dish, antifungal activity of essential oils was found to be similar to or greater than the positive controls: benomyl (37.4 ± 0.10% inhibition growth) for the F. oxysporum f. sp lycopersici assay and carbendazym (29.9 ± 0.13%inhibition growth) for the C. acutatum assay (Figure 3). Zanthoxylum fagara showed the highest antifungal activity on C. acutatum (EC50 153.9 μL L-1 air) and Z. monophyllum was the most active against F. oxysporum f. sp lycopersici (EC50 140.1 μL L-1 air) (Table 3). Minor components of Z. fagara were (E)-caryophyllene, T-muurolol, and α-cadinol (in a concentration of 2.7, 5.2%, and 8.1% respectively) and Z. monophyllum (in a concentration of 1.0, 1.6, and 4.1% respectively) have shown antifungal activity against various phytopathogenic fungi including the Fusarium and Colletotrichum species when they were assayed as pure compounds. These results suggest that Z. fagara and Z. monophyllum activity can be attributed to the presence of these compounds (Chang et al., 2008). Table 3. Fumigant toxicity of Zanthoxylum species oils against Colletotrichum sp. and Fusarium oxysporum.
Zanthoxylum rhoifolium essential oil has β-myrcene as its main component (59.0%), which is a compound that has shown antifungal activity in previous studies (Chang et al., 2008). This result suggests that moderate activity of the oil may be due to the presence of β-myrcene or the mixture of this compound with other metabolites that are toxic to the insect and present in the oil. CONCLUSION We hoped to find new natural products for biocontrol of two important phytopathogenic fungi and stored-grain insect pest. The development of natural pesticides would also help to decrease the negative impact of synthetic agents, such as residues, resistance, and environmental pollution. In conclusion, fruit essential oils of Z. monophyllum, Z. fagara, and Z. rhoifolium could be recommended as fumigants against F. oxysporum and C. acutatum and as alternatives to synthetic fungicides in agriculture. Fruit essential oil of Z. monophyllum also showed an important insecticidal activity against S. oryzae and could be proposed as an important alternative insecticide to control this pest. It is important to note that Z. fagara essential oil was inactive against S. oryzae, but presented significant antifungal activity against two phytopathogenic fungi, a result that shows the selectivity of this essential oil against the phytopathogenic fungi species and the pest insect used in the bioassays. However, further studies are required to determine the cost, applicability, and safety of these oils as potential pesticides; it should be determined which of the secondary metabolites present in essential oils are responsible for antifungal and/or insecticidal activities, and then to establish if these substances are produced by plants regardless of their ecological environment. ACKNOWLEDGEMENTS The authors are grateful to Colciencias and to the Universidad Nacional de Colombia for financial support. We also thank the Chromatography Laboratory at the Universidad Industrial de Santander for recording GC and GC/MS data and to the Colombian National Herbarium of the Universidad Nacional de Colombia for plant identification. LITERATURE CITED
Copyright 2011 - Chilean Journal of Agricultural Research The following images related to this document are available:Photo images[cj11009t3.jpg] [cj11009t2.jpg] [cj11009f2.jpg] [cj11009f3.jpg] [cj11009f1.jpg] [cj11009t1b.jpg] [cj11009t1a.jpg] | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| |||||||||

{kind=link}
{kind=link}
{kind=link}